
柳杉潜在适生区预测及气候变化对其分布的影响
2022-11-29 13:13:42戴旻峻李鑫玉王敏求文亚峰
西北林学院学报 2022年6期
戴旻峻,李鑫玉,王敏求,文亚峰
(中南林业科技大学 风景园林学院,湖南 长沙 410004)
气候是决定生物栖息地适宜性的主要决定因素之一,在过去百万年中,全球的平均气温和降水量在冰期和间冰期之间波动剧烈[1]。研究表明,在第四纪末次间冰期(LIG,距今约0.12~0.14 Ma)期间,全球平均温度较当前高2 ℃左右,而末次盛冰期期间(LGM,距今约22 000年),全球陆地约有24%被冰川覆盖,气候寒冷而干燥,导致许多物种分布范围大幅度缩减,甚至灭绝[2-3]。目前,随着现代社会碳质量浓度排放的增加,全球气温升高,政府间气候变化专门委员会(IPCC,Intergovernmental Panel on Climate Change)在第五次评估报告中发布了未来气候变化新情景RCPs(representative concentration pathways),模拟了4种不同CO2质量浓度下的气候数据用于研究分析。植物的生长与分布会随着气候变化而相应地扩大或缩小[4-5],潜在适生分布区的质心位置也会随之发生迁移。研究植物对气候环境变化的适应性,即不同气候变化情景下潜在适生区范围的动态变化,有利于探究影响植物生长的主要环境变量,为植物资源保护与引种栽培提供科学依据,对生物多样性保护具有重要意义[6]。
柳杉(Cryptomeriajaponicavar.sinensis)为东亚特有的第三纪孑遗植物,是现存杉科植物中最原始的类群。化石资料表明,柳杉在侏罗纪晚期或更早起源于东亚的东北地区,随后在适宜的气候环境下扩散至欧洲及北美等地区,而第三纪晚期和第四纪冰期的全球剧烈气候变化使得该物种在欧美地区相继灭绝[7],仅在我国东南部及日本群岛有天然种群得以留存[8]。Tsumura等利用14个日本柳杉天然种群和1个柳杉种群进行了物种分布区模拟(SDM)分析,结果表明,气候变化,尤其是降水,对柳杉的种群数量及其分布具有深刻影响,但遗憾的是仅使用了1个柳杉种群,无法充分说明柳杉在我国的适生区分布范围[9]。Cai等[10]认为全新世以来,由于生境破坏、栖息地减少与人类活动等的干扰,进一步导致了柳杉的种群数量进一步衰减。柳杉具有较高的经济和观赏价值,是良好的用材林和园林绿化树种。目前,仅在我国浙江、福建和江西等地有少量天然种群留存,因此探究其当前及未来气候变化下潜在分布区的变化,对该物种的保护与引种利用具有重要意义[11]。
近年来,MaxEnt模型作为评价物种生存环境范围最常用的模型,被广泛应用于农作物适宜区预测、动植物潜在适生区评价、外来入侵物种风险评估和药用植物潜在生境分布等领域[12-14]。MaxEnt模型较之于其他生态位模型,具有所需样本量少、受样本影响小、预测结果较好等优点[15],已在花榈木(Ormosiahenryi)[16]、沙冬青(Ammopiptanthusmongolicus)[17]和闽楠 (Phoebebournei)[18]等植物中应用且均取得了较好的效果。目前,柳杉在我国的潜在适生区分布范围以及不同气候情景下影响其分布范围的变化情况尚待研究。基于此,本研究结合MaxEnt模型与ArcGIS空间分析技术,依据我国柳杉分布点数据和20个环境变量,预测不同气候情景下柳杉的潜在适生区范围及其变化,解析气候变化对柳杉资源分布的影响,以期为我国柳杉遗传资源保护、引种以及扩大柳杉适生区分布范围提供科学依据。
1 材料与方法
1.1 物种分布数据的获取与处理
从中国数字植物标本馆(CVH,http://www.cvh.org.cn/)、全球生物多样性信息网(GBIF,http://www.gbif.org)、《中国植物志》、野外实地调查和公开发表的文献获取柳杉在我国的分布点数据。为确保样点信息的准确性,通过Google Earth进行坐标拾取并校准,删除重复和错误分布点。此外,为避免群集效应导致MaxEnt模型过度拟合(Overfitting)[19],在地图上1 km×1 km的网格中仅保留1个分布点,最终获得80个柳杉在我国的有效地理分布点(图1,审图号GS(2019)1831号),并将分布点的经纬度坐标保存为.csv格式用于后续分析。
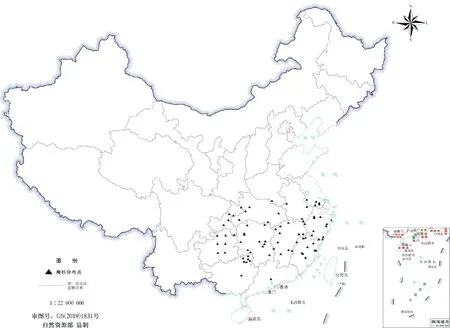
图1 柳杉在我国分布点信息
1.2 环境数据的获取及筛选
从全球气候数据库(Worldclim,http://www.worldclim.org/)下载当前、过去及未来气候情景下的19个环境变量(bio1~bio19)和1个地形变量(海拔,Alt),空间分辨率为30″(地理空间分辨率为1 km)(表1)。其中,当前时期的气候数据下载于1970-2000年;末次盛冰期气候数据(LGM)来源于日本东京大学气候系统研究所、日本环境研究所和日本全球环境研究中心联合开发的适用于东亚地区的MIROC模式[20];未来时期的(2070s)气候情景采用我国国家气候中心开发的BBC-CSM模式,选取3种典型浓度路径情景(RCPs)RCP 2.6、RCP 4.5和RCP 8.5,分别对应辐射强度2.6、4.5、8.5 W/m2的温室气体的排放,故本研究共涉及5种情景。
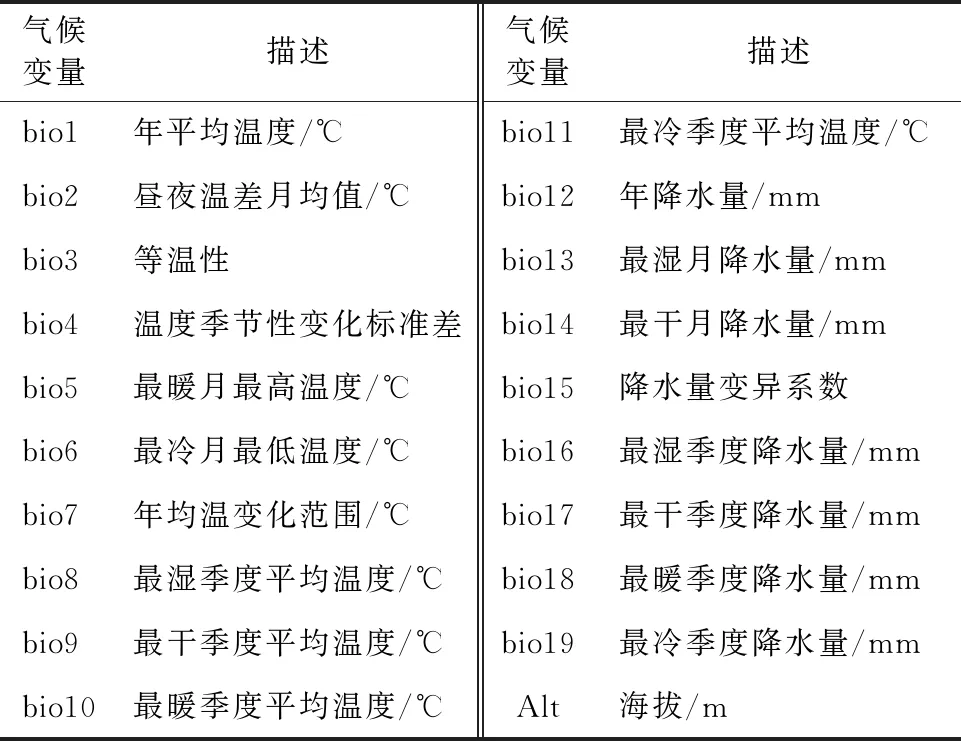
表1 柳杉地理分布评价的19个环境变量和1个地形变量
考虑到环境变量间存在较强的多重共线性,可能会引入冗余信息导致模型过度拟合[21-22],因此,需要多个环境变量进行相关性分析和筛选。基于ArcGIS10.4软件提取得到的柳杉分布点生物气候变量及地形变量的数据,利用R中的“cor”函数计算各环境变量之间的相关性,并运用“corrplot”程序包输出可视化结果[23],保留相关性较低的生物气候变量(|r|<0.8),最终筛选得到5个变量用于后续模型预测。
1.3 MaxEnt模型预测及其适生区划分
基于筛选得到的环境变量,利用MaxEnt 3.4.1软件[24]与ArcGIS10.4软件预测不同气候情景下柳杉在我国的潜在适生区范围。MaxEnt参数设置如下:75%的数据作为训练集用于模型构建,25%的数据作为测试集用于模型检验,重复模拟10次;刀切法(Jackknife test)计算各生物气候变量的贡献率;受试者工作特征曲线(receiver operating characteristic,ROC)检测模型精度,并计算AUC值(训练数据集和测试数据集的受试者工作特征曲线下的面积,是检测模型精确度的指标);运用R中的“ggplot”程序包绘制环境变量核密度图。最后利用ArcGIS 10.4软件对结果进行重分类与可视化,采用自然间断点分级法(Jerk’s natural breaks)将适生区依据分布频率P划分为4个等级:非适生区(0≤P<0.1,灰色)、低度适生区(0.1≤P<0.3,黄色)、中度适生区(0.3≤P<0.5,橙色)、高度适生区(0.5≤P<1,红色)[22]。
1.4 分布区域范围变化与质心迁移
利用ArcGIS 10.4软件中的SDMtoolbox工具[22]模拟不同气候情景下的适生区范围变化趋势。将当前时期与末次盛冰期(LGM)、3个未来气候情景(RCP 2.6、RCP 4.5、RCP 8.5)的适生区栅格图层文件分别重叠,明确其扩增、收缩以及保留区域,计算过去、当前与未来适生区的面积变化及其质心位置,描述不同气候情景下质心迁移的方向和距离,阐明柳杉在中国适生区分布的动态迁移路径[23-24]。
2 结果与分析
2.1 MaxEnt模型预测精度
受试者工作特征曲线(ROC)检测结果显示(图2),训练数据集(training data)与测试数据集(test data)的AUC值分别为0.947和0.942,均接近于1,说明模型具有较高的稳定性和精度,模拟结果准确,可用于柳杉适生区划分研究。
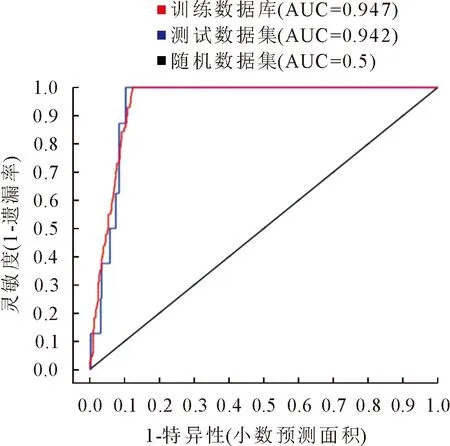
图2 柳杉适生区模拟结果ROC曲线评价
2.2 环境变量贡献率及其变化范围
根据相关性检验结果,从19个环境变量和1个地形变量中最终筛选得到5个变量(|r|<0.8)用于模型构建,分别为昼夜温差月均值(bio2)、等温性(bio3)、最湿季度平均温度(bio8)、降水量变异系数(bio15)和最暖季度降水量(bio18)。MaxEnt模型计算得到各个环境变量贡献率与刀切法检验结果显示(表2、图3),降水量变异系数(bio15)的贡献率最高(45.0%);其次为最暖季度降水量(bio18)(34.8%),昼夜温差月均值(bio2)、等温性(bio3)和最湿季度平均温度(bio8)的贡献率分别为16.4%、3.7%和0.2%。前4个生物气候变量的总贡献率达到了99.8%,说明这些变量是构建模型的主要环境变量。昼夜温差月均值(bio2)与降水量变异系数(bio15)对标准训练增益率分别达到1.56和1.42,说明这2个变量单独使用时能比其他环境变量包含更多有效的气候环境信息。
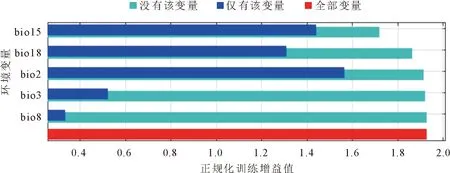
图3 环境变量刀切法检验
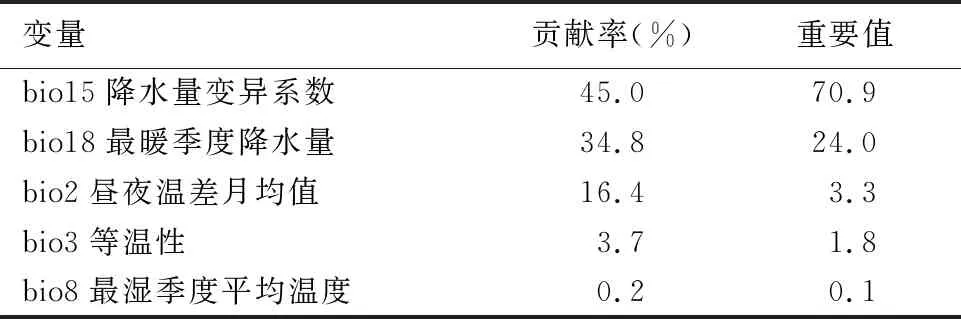
表2 各环境变量对柳杉分布的贡献率
影响柳杉适生区分布的5个主导环境变量的数值范围分析结果显示(图4):当前气候情景下,柳杉适生区环境变量的数值范围为:最暖季度降水量(bio18)最适宜的变化范围在500~1 100 mm,且600 mm时适生概率达到最高;降水量变异系数(bio15)最适宜的范围为58~60;昼夜温差月均值(bio2)在8 ℃时达到最适宜状态;等温性(bio3)最适宜范围在24.4~26.8。较未来气候情景(RCP 4.5)相比,柳杉适生区分布的降水量和温度略有增加,但是每个变量的分布概率都明显降低。
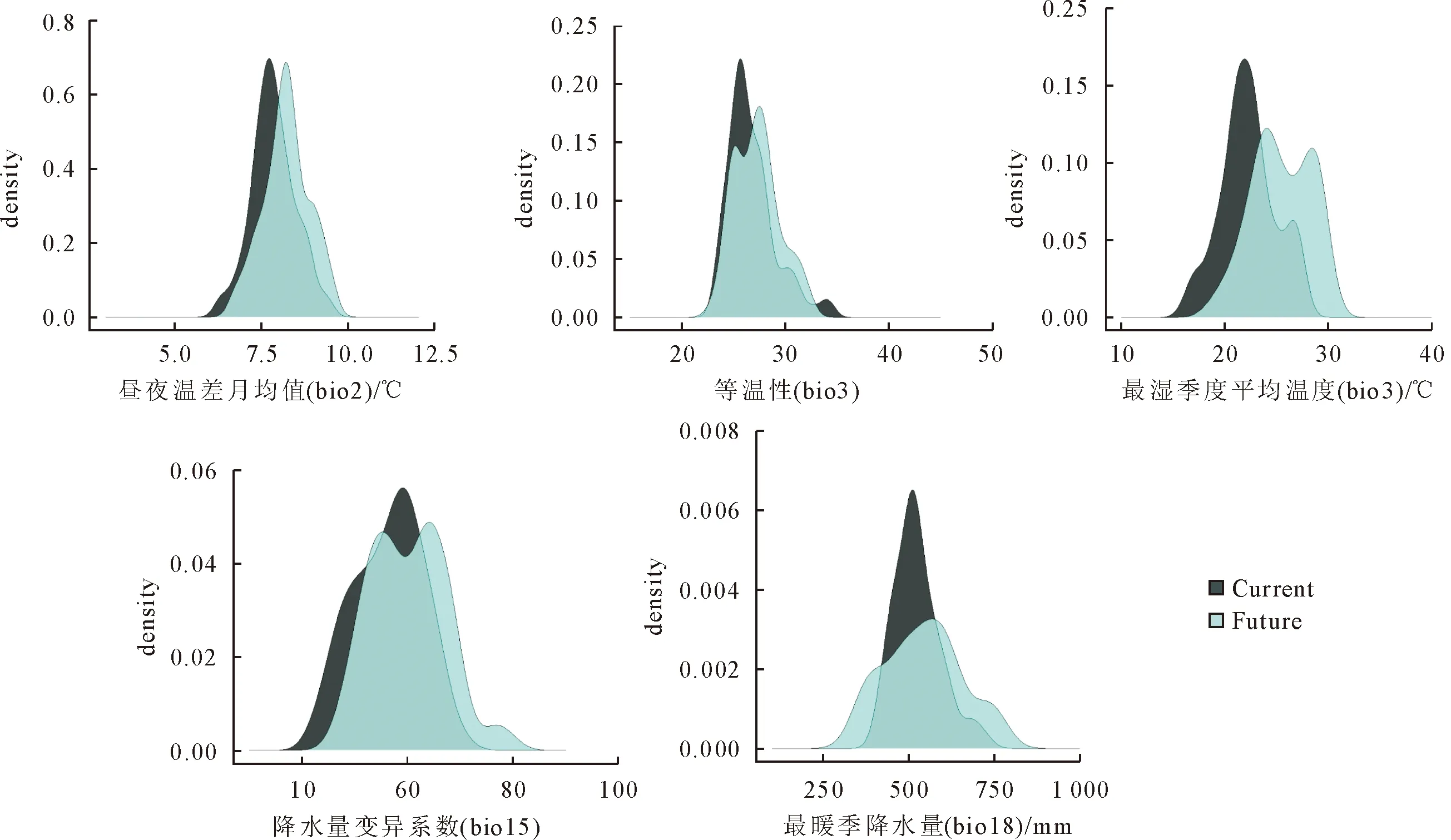
图4 影响柳杉分布的5个环境变量的数值范围变化
2.3 不同气候情景下柳杉的适生区分布及其动态变化
MaxEnt模型预测结果显示,当前时期柳杉在我国的最适分布区范围为107°42′-121°48′E,24°36′-32°00′N(图5),总适生区面积153.558×104km2,其中高适生区面积58.402×104km2,占总适生区面积的38.03%,核心分布区集中于长江以南地区,包括湖南、湖北、江西、福建、安徽和浙江等省区(表3、图5)。
末次盛冰期(LGM)时,柳杉在长江以南的地区广泛分布,总适生区的面积较当前时期增加了0.590×104km2,其中,高适生区和低适生区的面积分别增加了0.480×104、0.356×104km2,但中适生区的面积则减少了0.256×104km2。
较当前时期,未来3种CO2排放情景(RCP 2.6、RCP 4.5、RCP 8.5)下,柳杉总适生区的面积略有减少,但高适生区面积随CO2质量浓度升高呈先增加后减少的趋势,其中在RCP 4.5的排放情景下,面积达到最大(60.324×104km2)(表3),主要增加区域为适生区中部及湖南中部岳阳洞庭湖区和永州阳明山、大云山区域(图5、图6)。
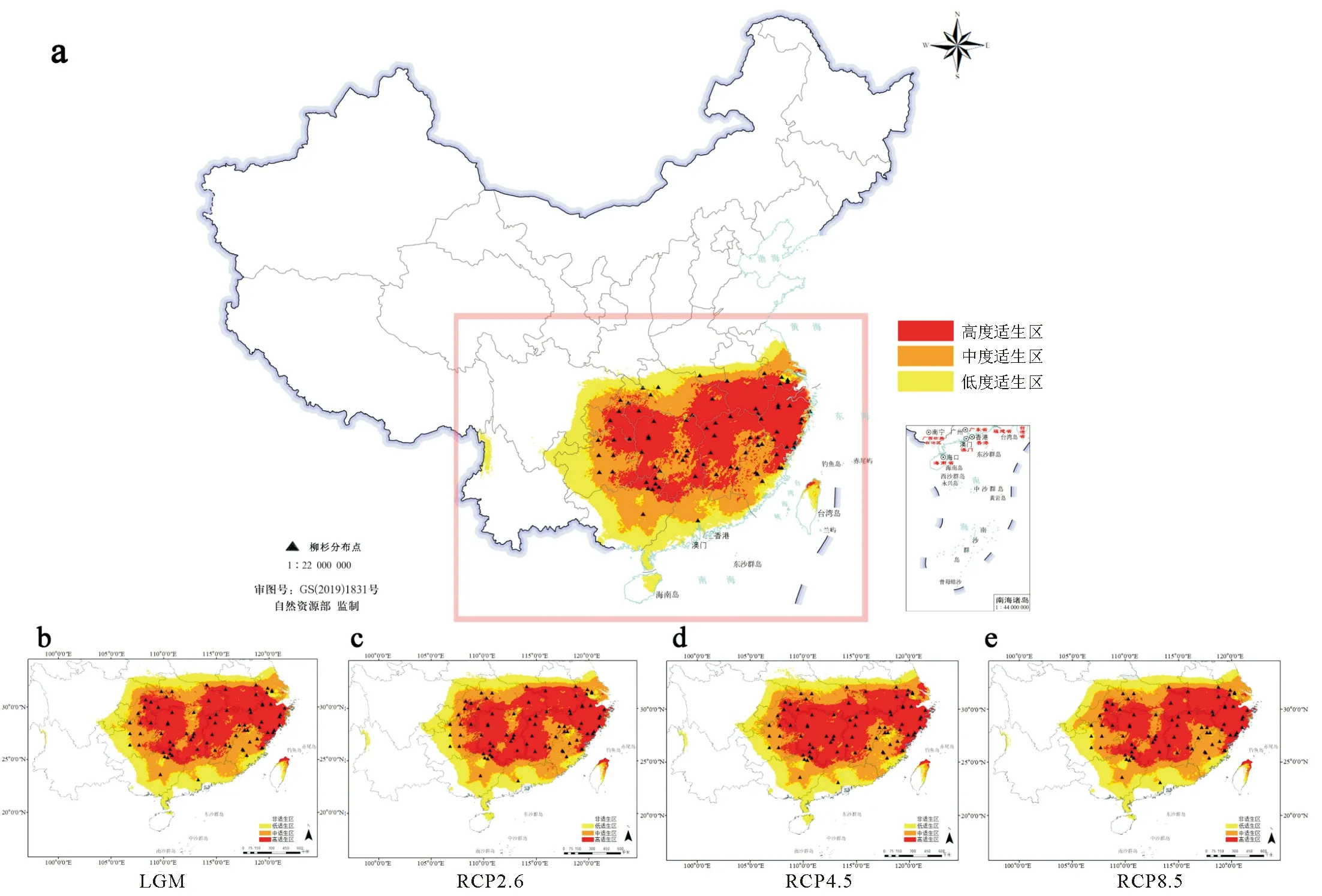
注:a为当前;b为末次盛冰期;c、d、e为未来3种不同气候情景下(RCP 2.6、RCP 4.5、RCP 8.5)。
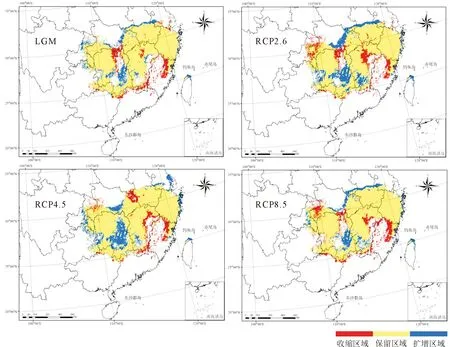
注:LGM时期与未来2070s均以当前时期作为基准。
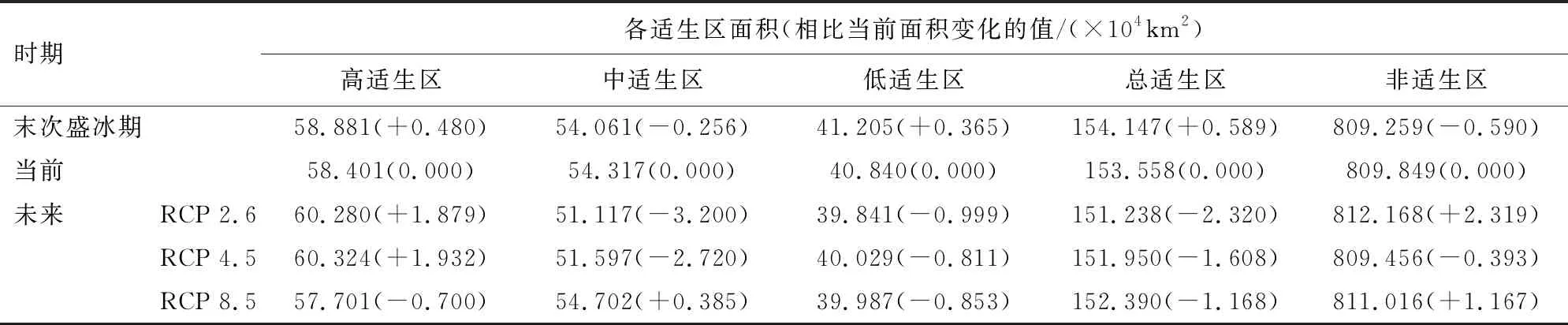
表3 柳杉不同气候情景下适生区面积
质心迁移分析结果显示,所有质心位置纬度均在28°-29°N。当前时期,适生区质心坐标为114°45′27″E和28°36′06″N。在LGM时期,质心较当前柳杉适生区质心位置呈现向东偏南方向迁移了29.178 km。未来(2070s)不同气候情景下,质心迁移方向和距离不同。其中,RCP4.5气候情景下柳杉潜在适生区质心位置较当前时期迁移距离最远,为33.122 km。而在RCP8.5和RCP2.6气候情景下,质心位置分别向西(12.303 km)和向北迁移(12.920 km)(图7、表4)。
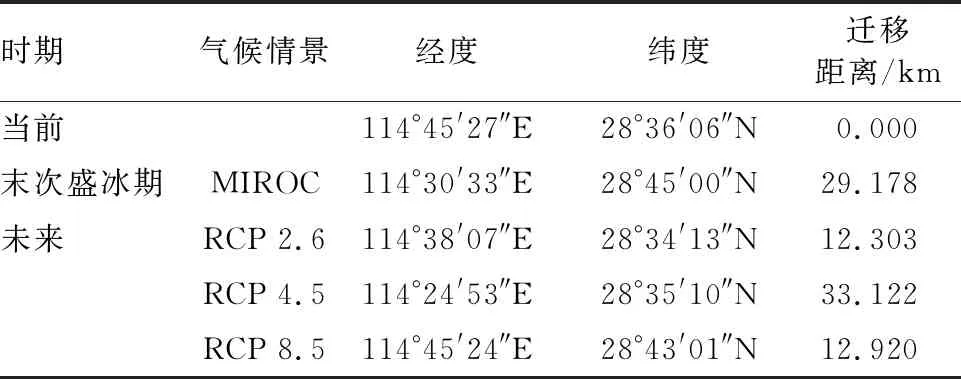
表4 质心坐标与迁移距离
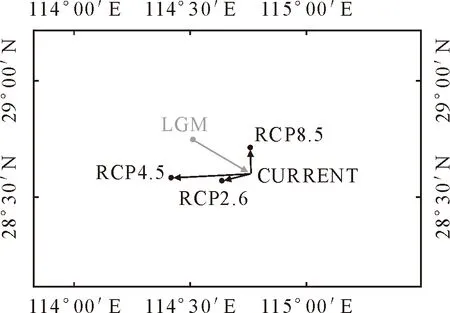
图7 不同气候情景下直线的迁移路线
3 结论与讨论
3.1 影响柳杉适生区分布主导环境变量
本研究结果表明,影响柳杉适生区分布的主要环境变量为降水量变异系数(bio15)、最暖季度降水量(bio18)、昼夜温差月均值(bio2)、等温性(bio3)和最湿季度平均温度(bio8),前4个变量贡献度高达99.8%,其中降水量变异系数(bio15)和最暖季度降水量(bio18)的贡献率较高,说明柳杉适生区分布主要受降水和温度影响,这与其生物学特性有关。植物对生长环境的需求,在一定程度上决定了该物种的分布范围。有研究表明,柳杉为中等喜光植物,喜温暖湿润、夏季凉爽的山区气候,常生长于海拔400~2 500 m的山谷边、潮湿的树林或山坡中[25]。本研究结果与Tsumura等(2020)利用广义加法模型(GAM)物种分布模型(SDM)评估气候变化对东亚地区柳杉遗传分化与种群分布的结果一致[10]。环境变量(年平均气温与降水量)与分布概率的响应曲线结果显示,柳杉最适宜的年均温范围为15~20 ℃,年均降水量范围为1 200~1 960 mm。由此可见,柳杉最适宜生长于降雨量充沛且昼夜温差较大的山林地区,不适宜在寒冷干燥、土壤贫瘠的地区[20,26]。因此,建议选取柳杉引种栽种位置时,选择降水丰富、气温凉爽和排水通风良好的地点。
3.2 柳杉适生区分布对气候变化的响应
本研究基于MaxEnt模型,分别模拟了柳杉在末次盛冰期(LGM)、当前时期以及3种未来CO2排放场景(2070s)的潜在适生区范围。结果显示,各时期均取得了较好的模拟效果(0.9 气候环境变化对植物现代地理分布的影响较为深刻,尤其是在第四纪冰期-间冰期的循环时期[27]。第三纪晚期至第四纪冰期期间,受全球气候变化影响,导致许多动植物在欧美等地区相继灭绝,分布范围大幅度缩减,曾经广泛分布的第三纪孑遗植物逐渐退缩至东亚、北美或欧洲西南部的冰期避难所[28]。气温变暖,降水减少会严重影响植物的生长和生理生态特征,导致大多数植物的地理分布格局发生显著变化,有的甚至会直接导致许多植物物种的原始分布区域破碎化,种群数量骤减,最终致使物种灭绝,造成生物多样性的降低[29-30]。本研究结果显示,末次盛冰期(LGM),柳杉在我国的适生区范围最广。与其他广泛分布日本和中国东部许多温带物种不同,柳杉分布范围在冰期收缩后并未发生扩张,并在未来气候情景下呈持续缩减趋势。造成此现象的原因可能与全球气候变化导致的降水量减少有关。此外,由于对木材的需求,大量采伐也导致柳杉在我国的种群数量及种群分布范围的减少[10]。因此,研究认为气候环境变化,尤其是降水的减少,是导致柳杉适生区分布区范围持续缩减的主要原因。 在气候变暖影响下,物种的适生区分布范围将缩减并向高纬度迁移,如天山花楸(Sorbustianschanica)、薄片青冈(Quercuslamellosa)[31-32]。但本研究结果显示,柳杉的适生区分布范围仅在RCP 8.5(2070s)气候场景下存在此现象,在其他2种未来气候情景下(RCP 2.6和RCP 4.5)均存在“西迁”的变化趋势。不同气候情景下其分布区范围质心的迁移方向存在差异,可能是由于人类活动和气候变暖程度不一,存在不确定性有关。此外,在未来气候情景下柳杉分布区范围质心的迁移距离也存在差异,其中RCP4.5迁移距离最远。但总的来讲,柳杉适生区分布范围的质心位置与距离相差不大,基本稳定在江西和湖南省交界处,28°-29°N,114°-115°E。 研究结果表明,自末次盛冰期以来,受气候变化与人为干扰因素的影响,柳杉在我国的适生区范围持续缩减,说明当前柳杉的保护情况并不乐观。虽然柳杉在我国资源丰富,人工种群的数量远大于天然种群的数量,但天然林种群能够保留更多的私有等位基因,具有较高的遗传多样性,是研究与保护该物种最佳材料。根据前期调查与研究,建议江西、福建、浙江省分布的柳杉古树或天然种群应加强就地保护,建立自然保护区,最大程度地减少人为破坏与干扰。而对于广泛分布的柳杉人工资源,可基于MaxEnt模型预测结果,有针对性地对其资源进行保护与利用,主要环境变量可为柳杉繁育种植区域的环境因素提供参考依据,营造最适宜的柳杉生长环境,实现资源合理开发利用。此外,可在中适生区适当增加柳杉的迁入,使其有利于柳杉种群繁衍和扩散。对于高适生区,应就地保护与人工栽培并行。鉴于柳杉重要的生态和经济价值,需加强公众对柳杉的了解,禁止乱砍滥伐等行为。此外,在迁地保护及引种栽培过程中,除了要遵循适地适树的原则外,还应综合考虑病虫害、种间竞争和土壤质地等因素,以提高柳杉栽培成活率。3.3 柳杉遗传资源的保护与利用
猜你喜欢
中国野生植物资源(2023年9期)2023-10-20 09:11:34
中国中医药信息杂志(2023年6期)2023-06-13 02:05:36
安徽农学通报(2022年6期)2022-04-07 21:30:29
浙江林业(2020年7期)2020-08-06 14:55:32
电脑爱好者(2020年10期)2020-07-28 17:10:30
植物资源与环境学报(2020年2期)2020-04-20 07:18:02
电脑爱好者(2019年16期)2019-10-30 03:35:12
竹子学报(2018年2期)2018-10-25 02:02:12
中成药(2018年3期)2018-05-07 13:34:25
喜剧世界(2016年7期)2016-08-31 06:21:06
