
基于稳定同位素技术的三峡库区短颌鲚营养生态位特征研究
2022-11-26 08:18吕红健杨丽亚唐晓琪邓华堂张志新姚维志
生态学报 2022年21期
吕红健,杨丽亚,付 梅,*,唐晓琪,邓华堂,张志新,姚维志
1 西南大学水产学院,农业农村部长江上游水生生物多样性保护研究中心,西南大学渔业资源环境研究中心,重庆 400715
2 中国水产科学研究院长江水产研究所,农业农村部长江中上游渔业资源环境科学观测实验站,武汉 430223
3 东京海洋大学海洋科学学部,东京 108- 8477
生态位是生态学中的一个重要概念,至今生态位理论研究发展史已越百年,生态位理论日臻完善[1]。生态位主要指某一物种在生物群落或生态系统中的地位和角色[2]。其中,营养生态位为生态位研究的重要范畴,指动物对其食物资源能够实际和潜在的占据、利用或适应的部分,营养生态位在一定条件下可以反映动物在所研究区域的食性情况,同时可反映动物对资源的利用能力及其在群落或生态系统中的功能位置[3—4]。目前,营养生态位理论已成为探究物种入侵机制,预测外来(或入侵)物种如何对本土物种产生影响及二者间共存模式,判别水域系统中功能相似物种的食物竞争状况等研究的重要支撑理论[5—7]。鱼类营养生态位研究中,以往常采用胃含物分析法探讨鱼类的食物来源,并根据食物组成确定营养生态位宽幅和生态位重叠[4]。近年来,随着稳定同位素技术的发展及其应用领域的拓宽,碳(δ13C)、氮(δ15N)稳定同位素技术被引入营养生态位研究领域[8—10]。与传统胃含物分析方法相比,碳、氮稳定同位素分析法可反映鱼体较长生活周期的营养状况[11—12],已在鱼类营养生态位研究领域得到广泛应用[13—15]。
已有研究表明,水库在蓄水的初期阶段,由于水体营养动力学的改变通常有利于广适性鱼类和外来种的生存,而威胁土著鱼类和特有种[16—17]。三峡库区蓄水以来,随着库区水体加深、水流减缓等变化,形成了一个生态位严重空缺的人造湖泊生态系统[18—20],外来种定殖,对土著种和特有种可能造成严重威胁。迄今为止,三峡库区已发现外来鱼类20余种,且部分种类处于种群数量的暴发阶段[19—20]。如太湖新银鱼(Neosalanxtaihuensis)作为外来物种,自2006年以来,已经成为三峡库区重要的渔业捕捞对象[21]。
短颌鲚(Coilianasus,曾被命名为Coiliabrachygnathus)隶属鲱形目(Clupeiformes)、鳀科(Engraulidae)、鲚属(Coilia),为刀鲚的淡水定居型[22]。据历史调查资料记载,短颌鲚主要分布在长江及淮河流域的中、下游,以及与长江流域相通的淡水湖泊,如鄱阳湖和洞庭湖[23—24]。相关研究表明短颌鲚具有适应力强、性成熟早、繁殖率高的生物学特性[25]。2016年有媒体首次报道了三峡库区部分区县渔民捕捞到“刀鱼”,而后通过线粒体DNA全序列检测方法鉴定三峡库区“刀鱼”实为短颌鲚[26]。另据西南大学渔业资源环境研究中心2018—2020年调查数据显示短颌鲚在三峡库区已经形成了可持续种群。因此,根据生物入侵定义[27],并结合现有的研究资料表明短颌鲚为三峡库区的外来鱼种,但尚不能定义为三峡库区“入侵种”。然而,截至目前针对短颌鲚在三峡库区的分布范围和资源现状研究仍是空白,且有关短颌鲚在三峡库区水生生物群落中的营养生态位特征也未有报道。本研究拟利用碳、氮稳定同位素技术研究短颌鲚及其营养级相近和食性相近物种在三峡库区的营养位置、营养生态位宽幅及营养生态位重叠状况,进而评析库区短颌鲚的营养生态位分化状况,预测短颌鲚的种群发展趋势,为探讨短颌鲚对三峡库区水域生态系统的影响提供科学依据。
1 材料与方法
1.1 研究区域
本研究所涉及的鱼类、螺类样品均于2019年7至10月(即三峡库区枯水期),在位于三峡库区腹地的长江一级支流草堂河及大宁河采集,采样点地理位置见图1。

图1 采样点示意图
1.2 样品采集与处理
1.2.1 样品预分析与研究对象确定
本研究使用双层刺网(网目内径为60mm)和定置串联倒须笼壶(网目内径为30mm)先采集研究水域杂食性与肉食性典型鱼类各3—4尾样本,测量并记录体长(mm)和体重(g)(样本采集信息见表1),通过碳、氮稳定性同位素分析,确定其营养级,初步判断短颌鲚在三峡库区相应水域所处营养位置。初步分析结果详见图2。

表1 实验样本的采集信息
在预分析结果的基础上,根据营养级,并结合胃含物分析结果,选取4种营养层级相近(包括太湖新银鱼、翘嘴鲌Culteralburnus、鳡Elopichthysbambusa、大眼鳜Sinipercakneri)及1种食谱存在较大重叠的鱼类(即鳙Aristichthysnobilis)作为研究对象。分别取短颌鲚及其上述5种鱼类各15个样本用于营养生态位特征分析。此外,依据三峡库区最新调查数据,从周年调查尺度来看,上述6种鱼类中,鳙属优势种,短颌鲚属亚优势种,翘嘴鲌、鳡、大眼鳜、太湖新银鱼均属常见种。
1.2.2 样品处理方法
鱼类样本采集后,测量并记录体长(mm)和体重(g),取其背部肌肉;螺类样本先放置于充分曝气的蒸馏水中过夜,使其肠含物排空,测量壳高(cm)、壳宽(cm)后去壳取其肌肉。所有肌肉样品用去离子水冲洗后,在60℃下连续烘干至恒重,最后用研钵研磨至均匀粉末,并干燥保存,以待碳、氮稳定性同位素比值测定。
1.3 稳定同位素分析
1.3.1 稳定同位素测定
所有样品的稳定性同位素比值测定由深圳精信同位素检测科技有限公司稳定同位素比率质谱实验室进行。使用的分析仪器为美国Thermo公司制造的Finnigan DELTA V Advantage同位素比率质谱仪和Flash 2000 EA-HT元素分析仪。碳、氮稳定性同位素分析分别采用国际标准物质PDB(Pee Dee Belnite)和标准大气氮对比。分析结果表示为δ13C 和δ15N:

式中,X为13C或15N;Rsample为样品所测得的同位素比值,即13C/12C或15N/14N;Rstandard为标准物质(PDB与大气中N2)的同位素比值。测定精度δ13C < 0.1‰,δ15N < 0.2‰。
1.3.2 营养级计算
营养级计算公式如下:
TL=(δ15Nconsumer-δ15Nbaseline)/Δδ15N+λ
式中,TL为消费者的营养级,δ15Nconsumer为消费者氮稳定同位素比值,δ15Nbaseline为水域生态系统中基线生物的氮稳定同位素比值,Δδ15N为相邻营养级之间氮稳定同位素富集度,λ为基线生物的营养级[28]。基线生物通常为寿命长、常年存在且食性简单的初级消费者[29],本研究选取三峡库区水域常年存在且广泛分布的铜锈环棱螺(Bellamyaaeruginosa)作为基线生物[30],铜锈环棱螺为初级消费者因此λ的取值为2,Δδ15N取值参考淡水生态系统中普遍运用的数值3.4‰[28]。
1.3.3 营养生态位的确定
分别以δ13C 值和δ15N值为横、纵坐标,绘制短颌鲚与4种营养级相近鱼类和1种食性相近鱼类的碳、氮稳定性同位素比值二维点集,以营养生态位总面积(即二维点集所围成的凸多变形总面积,Total Area,TA),小样本校正的标准椭圆面积(Sample size corrected version of the standard ellipse area,SEAC)及贝叶斯模型估算的标准椭圆面积(Bayesian estimates Standard Ellipse Area,SEAB)作为营养生态位宽度的度量指标。使用δ13C值变幅(δ13C value variational range,CR) 表示不同鱼类的摄食范围,δ15N值变幅(δ15N value variational range,NR) 代表不同鱼类的营养级跨度。以营养生态位总面积的重叠面积(Overlap Area,OA)作为营养生态位重叠的度量指标[9, 31—32]。TA、SEAC、SEAB均在 R 语言集成开发环境 R Studio中使用siar等数据包计算,OA应用数据分析工具Python计算。
1.4 数据处理
本研究中,数据的统计分析采用Microsoft Excel 2016, SPSS Statistics 20,R Studio 1.3.1073.0和Python 3.5.2;图表绘制采用Origin(2018)。并使用单因素方差分析法(One-way ANOVA)分析短颌鲚与营养级相近、食性相近鱼类的δ13C、δ15N值差异,设定差异显著性水平P为0.05,当P≤ 0.05时为差异显著,当P≤ 0.01时为差异极显著。
2 结果
2.1 三峡库区短颌鲚的营养级及相对位置
通过分析研究水域杂食性和肉食性鱼类典型代表的营养级发现,三峡库区短颌鲚营养级为3.762±0.196 (Mean±SD),较库区大部分鱼类高,仅次于铜鱼及太湖新银鱼,且其营养级略高于蒙古鲌、大眼鳜、鳡、翘嘴鲌(图2)。
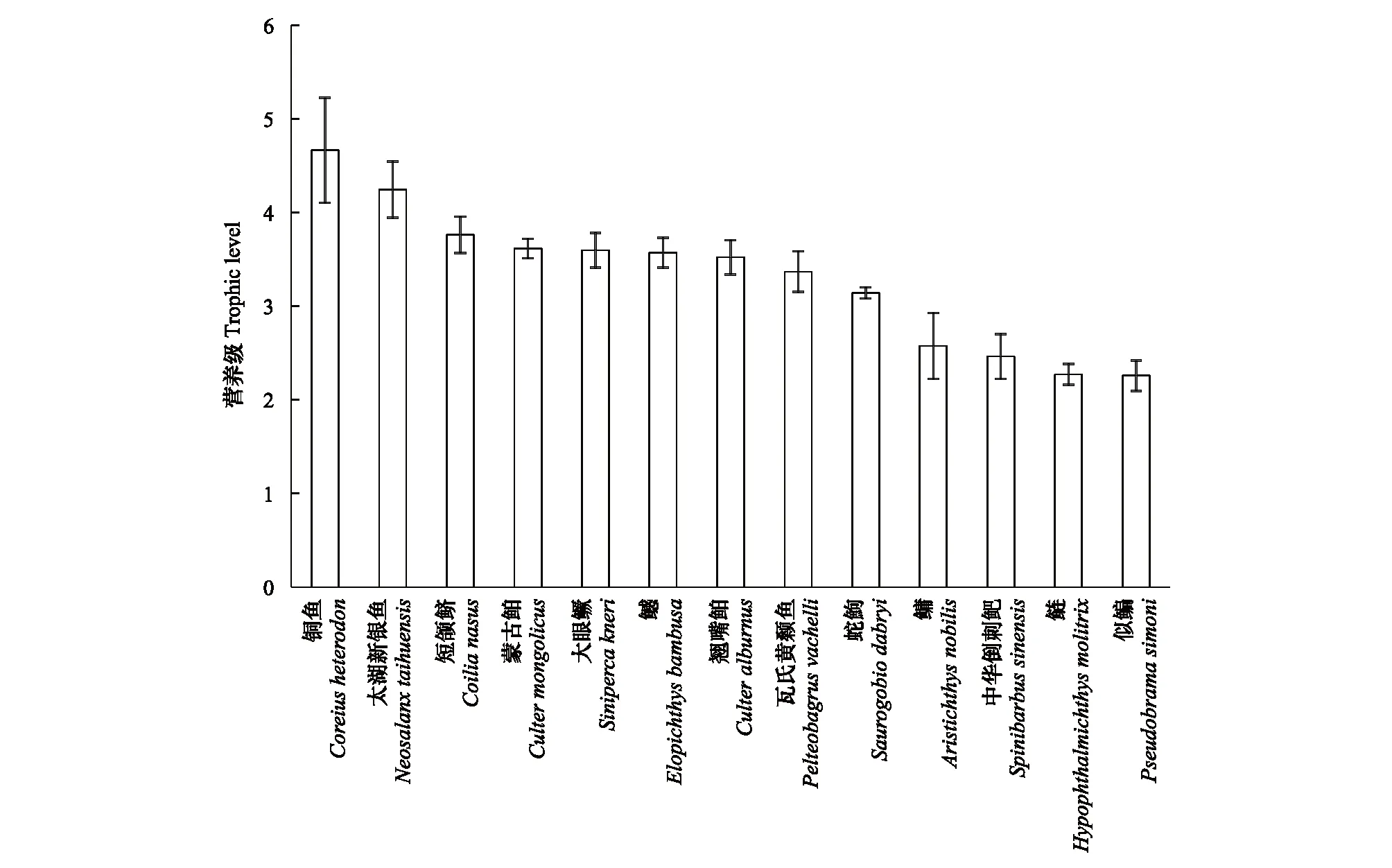
图2 三峡库区短颌鲚及其他12种鱼类的营养级比较
2.2 三峡库区短颌鲚与其他5种鱼类的稳定同位素比值组成特征

图3 三峡库区短颌鲚、太湖新银鱼、翘嘴鲌、鳡、大眼鳜、鳙的δ13C、δ15N均值及标准差
三峡库区短颌鲚δ13C值分布范围为-27.236‰—-22.952‰,平均值为(-25.379±1.213)‰(Mean±SD),δ15N值分布范围为12.112‰—14.497‰,平均值为(13.213±0.668)‰(Mean±SD)。单因素方差分析(ANOVA)结果显示,三峡库区短颌鲚δ13C值平均值较太湖新银鱼和鳙更高,与太湖新银鱼间存在显著差异(n=15,F=5.351,P=0.028<0.05),与鳙间无显著性差异(n=15,F=0.141,P=0.710 > 0.05),但该值低于鳡(n=15,F=42.341,P<0.001)、翘嘴鲌(n=15,F=26.221,P<0.001)、大眼鳜(n=15,F=62.257,P<0.001),且均呈极显著差异。三峡库区短颌鲚δ15N值平均值高于翘嘴鲌、大眼鳜、鳡、鳙,与鳡(n=15,F=7.175,P=0.012<0.05)间存在显著差异,与鳙(n=15,F=120.041,P<0.001)、翘嘴鲌(n=15,F=8.268,P=0.008<0.01)间存在极显著差异,与大眼鳜之间不存在显著差异(n=15,F=2.219,P=0.148>0.05),但该值低于太湖新银鱼(n=15,F=36.010,P<0.001),且呈极显著差异(图3)。
2.3三峡库区短颌鲚与其他5种鱼类δ13C值分布频率
三峡库区短颌鲚δ13C值分布范围为-27.236‰—-22.952‰,变幅仅小于鳙,大于太湖新银鱼、大眼鳜、翘嘴鲌及鳡,表明短颌鲚饵料来源范围较太湖新银鱼、大眼鳜、翘嘴鲌、鳡更广。δ13C值分布图显示,除鳙外,短颌鲚、太湖新银鱼、大眼鳜、鳡的δ13C值分布均呈单峰型,说明这4种鱼类对某些饵料生物有明显的摄食偏好性(图4)。此外,太湖新银鱼的δ13C值处于-28‰—-24‰之间,而短颌鲚δ13C值在此区间分布频率为80%,可见二者δ13C值分布高度接近,这表明短颌鲚与太湖新银鱼的摄食偏好趋近(图4)。
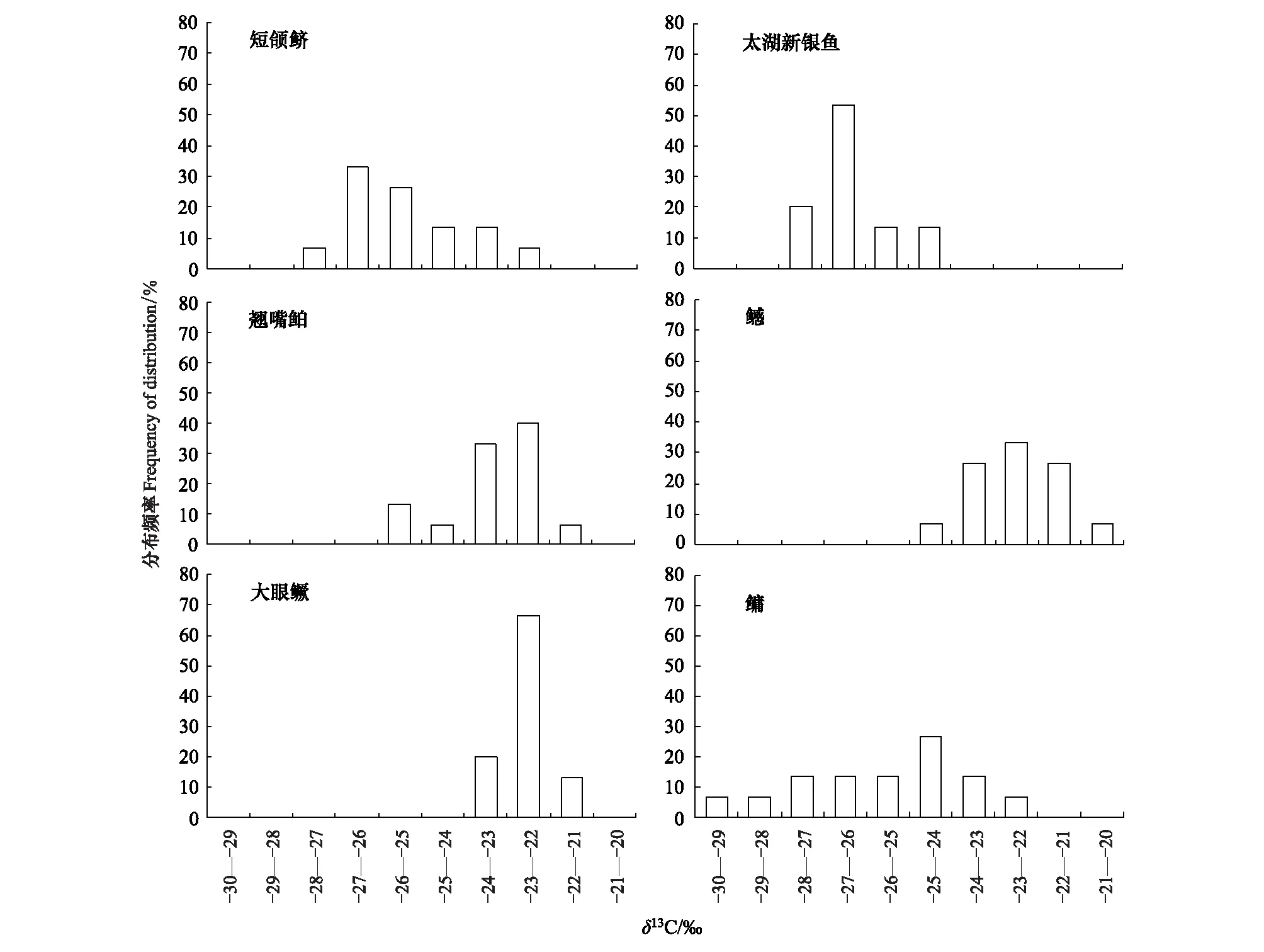
图4 三峡库区短颌鲚、太湖新银鱼、翘嘴鲌、鳡、大眼鳜、鳙的δ13C值分布频率
2.4 三峡库区短颌鲚与其他5种鱼类的营养生态位
三峡库区短颌鲚营养生态位被营养级相近物种所环绕,且目前处于相对空缺的位置(图5)。短颌鲚营养生态位宽幅指标,即营养生态位总面积(TA)、校正标准椭圆面积(SEAc)、标准椭圆面积的贝叶斯估算(SEAB)均低于鳡、鳙,高于太湖新银鱼、翘嘴鲌、大眼鳜。短颌鲚营养级跨度(NR)仅高于翘嘴鲌,低于鳙、鳡、太湖新银鱼、大眼鳜,表明短颌鲚饵料生物的营养级跨度较小(表2)。

图5 三峡库区短颌鲚的营养生态位
目前,三峡库区短颌鲚与其营养级相近物种的营养生态位重叠程度较低,其中短颌鲚与太湖新银鱼的营养生态位重叠面积最大,仅为0.489,分别占短颌鲚、太湖新银鱼营养生态位总面积的8.81%、12.64%。短颌鲚与其余4种营养级相近物种的营养生态位重叠度均低于该物种的营养生态位总面积的10%(表2,图6)。此外,短颌鲚与鳙的营养生态位重叠面积为0.000(表2,图6)。以上结果表明,就饵料资源利用而言,目前短颌鲚与其营养级相近物种间的竞争程度尚不激烈,存在较明显的营养生态位分化。

表2 三峡库区短颌鲚、太湖新银鱼、翘嘴鲌、鳡、大眼鳜、鳙的营养生态位宽幅及重叠面积

图6 三峡库区短颌鲚、太湖新银鱼、翘嘴鲌、鳡、大眼鳜、鳙的营养生态位
3 讨论
3.1 三峡库区短颌鲚稳定同位素组成特征及营养级相对位置
一般来说,消费者的δ13C值反映其饵料生物δ13C值的平均水平,所以δ13C常用来确定消费者的食物来源[33—35],δ13C值变幅(CR)越大,则消费者的饵料生物来源就越广泛[9]。研究结果显示三峡库区短颌鲚δ13C值变幅(CR)大于太湖新银鱼、大眼鳜、翘嘴鲌及鳡,表明短颌鲚饵料来源范围较广泛,这与邓华堂[30]、Zhang等[36]的研究结果一致。食性广泛通常会增加鱼类对生境的适应性,短颌鲚饵料来源广泛有利于其在三峡库区的定居,以及种群的增长和扩散。
已有研究表明δ13C值在食性相同或者相近的物种间差异一般较小,且变化幅度在0—1‰之间[34—35,37]。本研究中,单因素方差分析(ANOVA)结果显示三峡库区短颌鲚δ13C值与鳙间无显著性差异(P>0.05),与太湖新银鱼间存在显著差异(P<0.05),与鳡、翘嘴鲌、大眼鳜间存在极显著差异(P<0.01),且短颌鲚与鳙、太湖新银鱼的δ13C平均值差值均小于1‰,而其与大眼鳜、鳡、翘嘴鲌的δ13C平均值差值大于1‰。该结果表明三峡库区短颌鲚与鳙、太湖新银鱼摄食饵料生物种类可能存在较大程度的重叠,与传统食性分析结果一致[38]。从δ13C值的分布范围和频率研究结果综合来看,三峡库区短颌鲚、太湖新银鱼、大眼鳜和鳡等4种鱼类对某些饵料生物有明显的摄食偏好性,且短颌鲚与太湖新银鱼和鳙的摄食偏好趋近。本研究结果与刘恩生等[38]研究结果一致,其在比较太湖中1龄以内短颌鲚与太湖新银鱼的摄食特性时,发现二者均偏好摄食浮游动物。然而,短颌鲚的食性可能受环境及年龄等因素影响,如Zhang等[36]研究发现鄱阳湖短颌鲚δ13C值会随体长和季节的变化而发生更替或迁移。三峡库区短颌鲚食性随环境等因素的变化有待进一步研究。
物种在食物链中所处的营养位置称为营养级[30, 39]。20世纪80年代,就有研究表明消费者会随着其在食物链中所处的营养位置(即营养层级或营养级)的升高而逐步增加15N的富集,且幅度比13C大得多,消费者组织中的δ15N值往往比其饵料生物大(3.4±1.1)‰[35,40]。基于15N沿食物链富集的特点,消费者组织中的δ15N逐渐被用于确定该消费者的相对营养位置[41—42]。至今,基于δ15N值估计消费者营养位置的计算模型已日趋完善并得到广泛应用[28, 30, 43—44]。本研究结果表明三峡库区短颌鲚营养级为3.76±0.196 (Mean±SD),仅次于铜鱼及太湖新银鱼,略高于蒙古鲌、大眼鳜、鳡、翘嘴鲌,推测其原因为短颌鲚等杂食性鱼类的饵料来源广泛,因此铜鱼、太湖新银鱼和短颌鲚的营养级高于大眼鳜、翘嘴鲌、鳡等肉食性鱼类。邓华堂等[30]研究亦发现大宁河杂食性的铜鱼、光泽黄颡鱼(Pelteobagrusnitidus)和太湖新银鱼营养级较食鱼肉食性鱼类更高。李斌等[45]在研究三峡库区核心区域巫山至万州干流鱼类食物网结构时,也发现铜鱼拥有较高营养级,位于食物网的顶端,其主要原因是铜鱼的食物来源虾类在三峡库区营养级不低[31,45—46],进而导致了铜鱼在三峡库区营养级较高。已有研究认为短颌鲚食性介于浮游动物食性与食鱼肉食性之间[36,47],结合本研究中短颌鲚δ13C值变幅大于太湖新银鱼、大眼鳜、翘嘴鲌、鳡等鱼类,其饵料来源更为广泛,据此推测,相对于蒙古鲌、大眼鳜、鳡、翘嘴鲌而言,短颌鲚富集15N途径可能更丰富,15N富集在不同途径中占比不同可能最终导致了短颌鲚营养级高于大眼鳜等食鱼肉食性鱼类。同时,基于传统胃含物分析法和高通量测序分析法两种方法得到的短颌鲚在三峡库区的饵料生物名录也印证了上述分析结果,即短颌鲚在三峡库区的饵料生物共有7类(包括藻类、轮虫、枝角类、桡足类、甲壳类、水生昆虫、鱼类),共计33种(未发表数据)。此外,还发现三峡库区短颌鲚在蓄水期主要摄食日本沼虾(Macrobrachiummipponensis)和秀丽白虾(Leandermodestus),还存在严重的种内自残行为,即体型较大短颌鲚捕食体型较小短颌鲚的现象,且这一现象在2龄及以上年龄组均十分明显(未发表数据)。因此,蓄水期短颌鲚的主要食物虾类在三峡库区营养级不低[31,45],蓄水期其食物中15N的滞留效应也可能是短颌鲚营养级较高的原因之一。
短颌鲚在三峡库区营养级高于蒙古鲌、大眼鳜、鳡、翘嘴鲌等鱼食性鱼类,也得到了最新研究成果的验证,即何春等[48]在对三峡水库38种主要鱼类营养级研究中发现短颌鲚营养级处于第二位(仅次于铜鱼),并高于研究中所有鱼食性鱼类,包括蒙古鲌、翘嘴鲌、鳡、鲇(Silurusasotus)、沙塘鳢(Odontobutisobscurus)、大眼鳜、达氏鲌(Culterdabryi)等(顺序按营养级由高到低排列)。短颌鲚在营养级相对位置表现出的这一特征,在三峡库区以下,特别是长江中游的其他分布水域,例如长江中游干流(宜昌至湖口段)[49]、湖北荆州长湖[50]、江西鄱阳湖[51]等也存在类似现象。基于上述文献,以及有关碳、氮稳定同位素技术的研究报道,可大体将引起短颌鲚营养级相对位置高于鱼食性鱼类的原因总结为以下两点:(1)短颌鲚的摄食特性所引起的,特别是其对食饵的选择可能是引起生态位相对位置偏高的一个重要原因[49];(2)人类活动对水生生态系统的影响,包括人类活动产生的污水排放引起食物网中生物体内δ15N值的升高[50—53],以及人为捕捞压力所造成的鱼食性鱼类群体小型化和低龄化(因个体大、寿命长、自然死亡率低等原因,鱼食性鱼类更易成为主要捕捞对象),从而引起这些鱼类营养级相对位置的下降[54—55]。换言之,在鱼食性鱼类(包括蒙古鲌、翘嘴鲌、鳡、鲇、大眼鳜等)为主要捕捞对象的三峡库区,作为非主要捕捞对象,甚至是兼捕对象(即副渔获物)的短颌鲚,可能是在人类活动所造成污水排放(造成三峡库区水生生物体内δ15N值升高)和人为捕捞(造成鱼食性鱼类营养级相对位置下降)的双向作用下,导致了其营养级相对位置偏高的结果。此外,在长江十年禁渔的大背景下,为了弄清引起这一现象的原因,后续还有必要结合三峡库区污水排放调查,并进一步开展三峡库区短颌鲚摄食特性,以及营养级相对位置是否会随禁渔时间的延长而发生迁移的研究。
3.2 三峡库区短颌鲚及其营养级相近物种的营养生态位宽幅和生态位重叠
生态位宽幅和生态位重叠是描述种间生态位关系最重要的2个指标[56]。生态位宽幅常用以表征物种对资源及环境需求的特化程度,预测在不断变化的环境条件下,物种对环境的适应性[57]。生态位宽幅受到多个因子影响,包括食物密度及多样性、种间及种内竞争强度等[58—59]。通常,在资源供应不足时,物种的生态位宽幅越大,其对栖息地、食物资源的利用及竞争能力也就越强[58—60]。研究结果显示三峡库区短颌鲚营养生态位目前处于水域生态位相对空缺的位置,且其营养生态位宽幅指标TA、SEAc、SEAB在营养级相近物种中居于中间水平,低于鳡、鳙,高于太湖新银鱼、翘嘴鲌、大眼鳜。结合营养生态位宽幅指标和饵料生物分析结果,认为目前三峡库区的短颌鲚对饵料需求的特化程度不高,从营养角度对环境的适应性较强。
短颌鲚的生态位宽幅处于中间水平可能是种内竞争和种间竞争程度不激烈引起的。根据首次报道短颌鲚在三峡库区存在的时间,以及针对库区短颌鲚年龄与生长特性的研究数据(即短颌鲚年龄在1—6龄之间),推测短颌鲚进入到三峡库区的时间约在5—8年之间。因此,从种内竞争的角度,短颌鲚进入三峡库区时间较短,种群数量还未达到环境容纳量,即种群容量剩余量大,饵料生物相对充足。此外,营养生态位总面积的重叠面积(OA)结果表明三峡库区短颌鲚与其食性或营养级相近物种的营养生态位重叠程度较低,种间竞争程度不激烈。生态位重叠是指两个物种在同一资源状态上的相遇频率[57],是产生利用性竞争的一个必要条件[57,61]。本研究中,短颌鲚与太湖新银鱼的营养生态位总面积(TA)重叠程度最大,但重叠面积也仅占短颌鲚、太湖新银鱼营养生态位总面积的8.81%、12.64%。短颌鲚与鳡、翘嘴鲌、大眼鳜、太湖新银鱼4种营养级相近物种(食鱼肉食性)的营养生态位重叠度均低于该物种的营养生态位总面积的10%。此外,短颌鲚与鳙的营养生态位重叠面积为0.000。据此可以推断,就饵料资源利用而言,目前短颌鲚与其营养级相近和食性相近物种间的竞争程度尚不激烈,存在较明显的营养生态位分化。
关于生态位重叠的研究结果显示三峡库区短颌鲚与太湖新银鱼营养生态位重叠度较低,然而前文中δ13C值的分布范围和频率研究结果显示短颌鲚与太湖新银鱼的饵料偏好相同,这可能是短颌鲚与太湖新银鱼各自偏好摄食不同类型的浮游动物种类所引起的。刘恩生等[38]研究发现太湖新银鱼偏向摄食桡足类,而鲚属鱼类更偏向摄食枝角类。这一结果,在对三峡库区太湖新银鱼和短颌鲚胃含物样本镜检中也得到了验证。一般而言,在饵料供给较为充裕的条件下,物种摄食偏好性较强,易呈现出“专食者”特性[62],短颌鲚与太湖新银鱼营养生态位重叠度较低可能是对三峡库区水域充裕的浮游动物饵料条件的一种响应[30,63]。此外,短颌鲚与食鱼肉食性营养级相近物种营养生态位重叠度较低也可能与摄食策略分化(包括食性的分化、摄食空间的差异、食饵的选择与更替等)有关。
3.3 三峡库区短颌鲚未来的发展趋势
三峡库区短颌鲚饵料来源相对于太湖新银鱼、鳡、翘嘴鲌、大眼鳜更为广泛,其营养生态位目前处于库区水域相对空缺的位置,具有较充足的饵料资源基础。结合已有研究结果,即短颌鲚具有的适应力强、性成熟早、繁殖率高等生物学特性[25—26],推断短颌鲚种群规模将在三峡库区水域进一步扩大。加之,全球气候变化还将增加中国内陆淡水鱼类(包括外来鱼类)的栖息地适应性,从而增大部分水域的外来鱼类入侵风险[64],以及不断变化的社会经济活动,也可能进一步影响或增加外来鱼类的入侵途径[65]。基于上述几点考虑,推测作为外来鱼类的短颌鲚可能会进一步扩散至三峡库区所处长江上游地区的诸多水域,包括支流、水库、湖泊等。
最后,根据本研究结果推断三峡库区现有短颌鲚还未达到环境容纳量,未来不能排除短颌鲚种群数量继续增加的可能。三峡库区短颌鲚种群数量如若增至环境容纳量附近必将导致短颌鲚营养生态位的扩张。根据现有短颌鲚食性研究的历史资料[36, 47],可以推断短颌鲚营养生态位扩张将会影响,甚至可能破坏三峡库区水域现有水生生物多样性。此外,本实验结果仅代表三峡库区现有短颌鲚的营养生态位特征,由于短颌鲚在不同年龄段食性差别较大[47],因此三峡库区短颌鲚营养生态位未来还可能随着其种群平均年龄(或年龄结构)的改变而发生一定的迁移。在长江十年禁渔的大背景下,为了深入了解未来短颌鲚对三峡库区水生生物多样性的影响,后续有必要结合三峡库区短颌鲚种群年龄结构分析,并进一步开展三峡库区不同年龄段或不同体型短颌鲚食性分析研究。此外,十年禁渔背景下,三峡库区水生生物群落中其他鱼类的繁衍和种群恢复会对短颌鲚种群发展有什么影响,且短颌鲚种群扩张又会对其他鱼类的恢复产生怎样的影响,也是亟需解答的问题。
为了扭转长江生态环境恶化趋势,恢复长江天然渔业资源量及水生生物多样性,国家近年来采取了关键之举,例如“长江十年禁渔计划”[66]和近期出台的《中华人民共和国长江保护法》[67]。长江十年禁渔计划有利于长江水生生物资源量及多样性的恢复,但在禁渔期间,长江三峡库区天然渔业资源在“无捕捞”条件下的恢复趋势,即本土鱼类与外来鱼类各自的资源恢复趋势及相互影响,尚不明确。建国以来大水域尚未有长期禁渔后渔业资源恢复动态的报道,可供参考的研究有限,未来可以此为契机,开展三峡库区水域生态恢复与短颌鲚种群关联发展的相关研究,以期为提高三峡库区生态恢复效果提供科学的指导。目前,短颌鲚营养生态位特征研究结果表明,短颌鲚种群规模将在三峡库区水域进一步扩大,且就其食性而言[36, 47],其种群规模的进一步扩大可能会对三峡库区本土鱼类资源量及多样性恢复效果造成影响。因此,基于提高三峡库区水生生物资源量及多样性恢复效果的考虑,建议相关部门未来在长江十年禁渔期间应及时对三峡库区外来鱼类的种群数量和分布进行监测,必要时亦可对外来鱼类采取适当的人为控制手段。
猜你喜欢
当代水产(2021年10期)2022-01-12
读友·少年文学(清雅版)(2020年5期)2020-09-09
读友·少年文学(清雅版)(2020年5期)2020-09-09
读友·少年文学(清雅版)(2020年5期)2020-09-09
红领巾·萌芽(2019年8期)2019-08-27
教育教学论坛(2016年49期)2017-02-27
河北渔业(2014年11期)2014-11-17
河北渔业(2014年8期)2014-08-27
中学生物学(2008年8期)2008-10-27
雕塑(1995年4期)1995-07-12
