
基于生物信息学方法筛选精原细胞瘤进展中的脂代谢关键基因
2022-11-25 05:43:06闫墨刘帅兵王楷斌陈铭哲杨阔
天津医科大学学报 2022年6期
闫墨,刘帅兵,王楷斌,陈铭哲,杨阔
(天津医科大学第二医院泌尿外科,天津市泌尿外科研究所,天津 300211)
睾丸生殖细胞瘤虽然只占男性肿瘤总数的1%~2%,但是却在15~35岁男性中有着较高的发病率,是该年龄段男性最常见的恶性实体肿瘤[1]。睾丸生殖细胞瘤可分为精原细胞瘤和非精原细胞瘤。精原细胞瘤和非精原细胞瘤分别占总病例数的60%和40%。Ⅰ期精原细胞瘤患者的5年特异性生存率可高达99.6%,且放化疗对其有极佳的治疗效果,但是Ⅱ期、Ⅲ期患者的总生存率则低至50%,同时伴随着相对较大比例的复发[2-3]。
肿瘤细胞为了自身增殖和适应环境的需要,细胞内部的生物合成和代谢途径均会发生适应性的变化[4]。脂代谢作为诸多代谢途径的一种,在肿瘤细胞膜生物合成、细胞信号转导、脂化反应以及细胞代谢供能等方面发挥重要作用,通常在转变为恶性表型的肿瘤中,会发生较大的变化[5]。脂肪酸结合蛋白1(fatty acid binding protein,FABP1)分子量为14 kD,是一种可溶性蛋白,由127个氨基酸构成,主要在脂肪酸的摄取、运输、代谢和细胞内转运等方面发挥功能,同时在调节脂代谢以及细胞信号通路中也起到非常重要的作用。Wu等[6]发现FABP1在肾透明细胞癌中与转化生长因子-β1(TGF-β1)、过氧化物酶体增殖物激活受体α(PPAR α)和脂蛋白脂肪酶(LPL)共表达,且可以作为重要的肾透明细胞癌肿瘤标志物;Liu等[7]发现,FABP1表达随胃癌的进展逐渐降低,同时伴随着不良的预后,而这种不良的预后可能与其调节胃癌发展过程中的免疫微环境的作用相关。
笔者通过分析精原细胞瘤相关表达谱数据,发现FABP1在Ⅱ期、Ⅲ期精原细胞瘤中有明显的表达升高,这能显著影响患者的无进展生存。另外,FABP1与脂代谢相关基因显著相关,不同表达的样本中的免疫浸润也有明显差异,这将有助于理解精原细胞瘤进展过程中FABP1的作用机制。
1 材料与方法
1.1 数据资料下载与组织标本获取 从TCGA(The C ancer Genome Atlas,https://portal.gdc.cancer.gov/)和GTEx(Genotype-Tissue Expression Program,https://commonfund.nih.gov/gtex)下载精原细胞瘤(n=156)及睾丸正常组织(n=165)RNA-seq、临床生存及甲基化数据,以上为剔除缺失RNA与临床数据后最终纳入分析的数量。从GSEA(Gene Set Enrichment Analysis,http://www.gsea-msigdb.org/gsea/index.jsp)获取的代谢相关基因集分别为甘油酯代谢(GLYCEROLIPID_METABOLIC_PROCESS)、脂蛋白生物合成(LIPOPROTEIN_BIOSYNTHETIC_PROCESS)、脂蛋白代谢(LIPOPROTEIN_METABOLIC_PROCESS)、膜脂质代谢(MEMBRANE_LIPID_METABOLIC_PRO CESS)、磷脂生物合成(PHOSPHOLIPID_BIOSYN THETIC_PROCESS)以及磷脂代谢(PHOSPHOLIPID_METABOLIC_PROCESS),ISBN:0198506732。收 集病理科石蜡包埋的不同级别精原细胞瘤组织,每组3例。医院伦理委员会(NO.KY2014K112)批准这项研究并且所有患者均签署了知情同意书。
1.2 脂代谢相关差异基因筛选DESeq2包(v1.26.0)用来对Ⅰ期(n=80)以及Ⅱ、Ⅲ期(n=58)精原细胞瘤样本RNA-seq数据进行差异分析,筛选标准为|log2(FC)|≥1且P.adj<0.05,同时对差异分析得到的基因用Ensembl101(http://ftp.ensembl.org/pub/release-101/gtf/homo_sapiens/)进行分子类型的注释。将获取的差异表达基因(DEGs)与GSEA中6组代谢相关基因集中的基因取交集,得到在精原细胞瘤进展过程中与脂代谢相关的差异表达基因(MRGs)。
1.3 免疫组织化学法检测 按照通用型二步法免疫组化试剂盒(PV-9000)说明书中操作对FABP1(1∶500,Anti-Rabbit,Proteintech Group)进行染色。由3名经验丰富的病理科医师分别在相同条件下各自阅片,每张片子随机选取5个高倍镜视野观察,目标细胞胞浆出现淡黄色至棕黄色为阳性染色。根据染色强度对切片进行评价:阴性或未染色(无着色);弱阳性(浅黄色);中等阳性(黄色或者黄褐色);强阳性(棕色)。
1.4 MRGs的相关性、共表达、甲基化以及免疫浸润分析 将1.2分析得到的MRGs进行相关性分析,共表达分析及甲基化分析。GSVA包[8](v1.34.0)和estimate包[9](v1.0.13)用来分析精原细胞瘤RNA-Seq数据,评估精原细胞瘤患者TME中基质细胞比例和免疫细胞的浸润情况。相关系数的绝对值范围0~1,绝对值小于0.3为弱相关或不相关;绝对值介于0.3~0.5为一般相关程度;绝对值介于0.5~0.8为中等程度相关;绝对值大于0.8为强相关。
1.5 生存分析Survival包(v3.2-10)用来进行生存资料的统计分析,survminer包(v0.4.9)用于生存数据的可视化。
1.6 统计学处理R(v3.6.3)用于数据整理、统计分析与可视化。由Pearson秩和检验进行相关性分析。生存数据的统计方法为Cox回归。P<0.05表示差异有统计学意义。
2 结果
2.1 DEGs与MRGs分析结果 对精原细胞瘤RNA-seq数据分析后,共注释出56 493个基因,其中编码mRNA 19 577个,lnRNA 14 077个,miRNA 1 448个。满足差异基因筛选条件的mRNA编码基因共有1 865个,其中上调基因1 462个,下调基因403个(图1A)。去除GSEA下载的6组脂代谢相关基因集中的重复基因后获得691个脂代谢相关基因,与DEGs取交集后共有65个MRGs,见图1B。MRGs FABP1的log2(FC)=5.96,P.adj=1.63E-27,差异倍数与P值极为显著;FABP1在Ⅱ/Ⅲ期精原细胞瘤中有着明显的表达差异(P<0.001,图1C);免疫组化结果显示FABP1在高级别精原细胞瘤中有着更高的表达水平(图1D)。
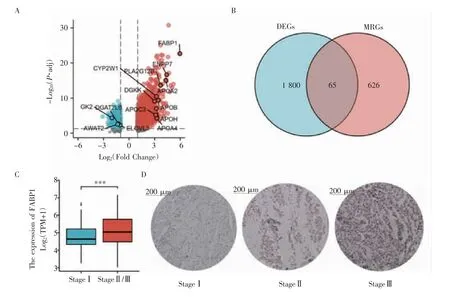
图1 DRGs和MRGs的差异表达基因筛选Fig 1 Differential expression genes screening in DRGs and MRGs
2.2 FABP1与MRGs的相关性、共表达及甲基化分析 从MRGs中筛选出差异显著上调基因碱性鞘磷脂酶(ENPP7)、载脂蛋白A2(APOA2)、磷脂酶A2 XII B(PLA2G12B)、载脂蛋白B(APOB)、载脂蛋白A4(APOA4)、细胞色素P450 2W1(CYP2W1)、载脂蛋白H(APOH)、载脂蛋白C3(APOC3)、二酰基甘油激酶κ(DGKK)和下调基因超长链脂肪酸延伸酶3(ELOVL3)、二酰基甘油O-酰基转移酶2样蛋白6(DGAT2L6)、二酰基甘油O-酰基转移酶2样蛋白4(DGAT2L4、AWAT2)、甘油激酶2(GK2),相关性分析结果显示FABP1与各基因均有显著的相关关系(P<0.01,图2A)。精原细胞瘤样本中FABP1与上调MRGs共表达分析结果按照相关性强弱进行排序,FABP1与各基因均有显著的共表达关系(P<0.001,图2B);除APOB和APOA4和FABP1有强相关关系外,其余各指标均与FABP1有中等相关强度;热图显示在低表达FABP1的精原细胞瘤样本中,各指标的表达低,高表达FABP1样本中各指标的表达高,显示出表达的一致性。下调MRGs的DNA甲 基 化 分 析 表 明 除 了GK2,ELOVL3、DGAT2L6、AWAT2在不同的甲基化探针中均存在较高程度的DNA甲基化(P<0.001,图3)。

图3 表达下调的MRGs在不同甲基化探针中的甲基化水平Fig 3 Methylation levels of down-regulated MRGs in different methylation probes
2.3 FABP1的免疫浸润分析24种免疫细胞的免疫浸润分析结果表明,FABP1的表达与自然杀伤(NK)细胞、NK CD56 bright细胞、肥大细胞、γδ(gamma-delta,Tgd)T细胞、未成熟树突状(iDC)细胞、巨噬细胞和树突状(DC)细胞存在显著正相关(P<0.01,图2C),其中与NK细胞,NK CD56 bright细胞存在中等强度以上相关;FABP1和NK CD56dim bright细胞、活化树突状(aDC)细胞、辅助性T细胞17(Th17)、调节性T细胞(Treg)、细胞毒性细胞、B细胞、T细胞、中央记忆型T细胞和辅助性T细胞存在显著负相关(P<0.01,图2C),其中与中央记忆型T细胞和辅助性T细胞的相关性强度较强。精原细胞瘤中与肿瘤微环境相关的基质细胞与免疫细胞浸润的结果显示高表达FABP1较低表达的肿瘤样本,基质得分高,而免疫浸润得分与估计得分低(图2D)。

图2 FABP1与MRGs的相关性、共表达以及免疫浸润分析Fig 2 Correlation,co-expression and immune infiltration analysis between FABP1 and MRGs
2.4 生存分析结果 高FABP1表达患者的无进展生存期较短。FABP1共表达脂代谢基因PLA2G12B、APOB、APOA2、APOC3、APOA4和CYP2W1的 生 存 结果与FABP1一致;而在精原细胞瘤中低表达的MRGs如DGAT2L6与AWAT2则显示相反的生存结果(P<0.01,图4)。

图4 FABP1与各指标高低表达组无进展生存期比较的生存曲线Fig 4 Survival curve of comparison of progression-free survival time in FABP1 and other genes high-low expression group
3 讨论
睾丸生殖细胞瘤作为15~35岁年轻男性中发病率较高的实体恶性肿瘤,因其发病的特殊性,往往对该年龄段男性的正常生活造成较大影响,除导致不育外,甚至会使患者产生不良预后而死亡,精原细胞瘤作为其中重要的类型更应给予足够的关注[10]。因为Ⅰ期精原细胞瘤患者具有极佳的生存率,所以为了在精原细胞瘤进展过程中鉴别其有无向高级别肿瘤进展的倾向,将TCGA数据库中Ⅱ期、Ⅲ期与Ⅰ期精原细胞瘤作差异表达分析筛选出FABP1及相关脂代谢相关差异基因。同时分析了FABP1与MRGs的相关性与共表达关系,MRGs在不同甲基化探针中甲基化的情况,不同表达的FABP1与肿瘤中免疫细胞的免疫浸润关系以及表达FABP1与MRGs患者的生存差异。笔者认为,FABP1可能通过调节肿瘤细胞与肿瘤微环境中免疫细胞的脂代谢途径,增强肿瘤细胞的增殖与转移能力,减弱免疫细胞对肿瘤细胞的杀伤作用,从而介导精原细胞瘤的进展。FABP1具有成为鉴别精原细胞瘤进展以及潜在治疗靶点的标志物的潜力。
细胞增殖是肿瘤的共性,而肿瘤的转移往往意味着预后不良,在此过程中,肿瘤细胞通常具有特征性的脂代谢变化,需要脂肪酸来合成细胞膜和细胞活动所需的信号分子[11-12]。Nieman等[13]发现,FABP4在卵巢癌网膜转移灶中较原发性卵巢癌中表达增多,作为肿瘤细胞中脂质运输的介质,在肿瘤生长和转移过程中起到关键作用。FABP1在Ⅱ期、Ⅲ期精原细胞瘤中差异表达升高极为明显,通过分析FABP1与其MRGs之间的共表达与相关关系,发现FABP1与APOB、APOA4、PLA2G12B、APOC3、APOA2、DGKK、CYP2W1、ENPP7共表达,这些基因编码蛋白参与脂质运输和细胞内细胞信号的传递;FABP1与ELOVL3、DGAT2L6、DGAT2L4、AWAT2和GK2呈负相关,对下调的MRGs进行甲基化分析,其中ELOVL3、DGAT2L6、AWAT2有较高的甲基化水平。DAGT可通过增加脂质的存储来减少可用脂质[11]。Bagnato等[14]发现DGAT1在人成纤维细胞中过表达可降低其增殖与侵袭性;而ELOVL3则能够参与形成脂滴、调控脂类合成以及增强脂肪细胞脂肪酸氧化能力[12]。综上所述,FABP1可能通过与脂代谢相关基因共同作用,增强精原细胞瘤细胞对脂肪酸摄入的同时,增加可用脂质以及减少脂肪酸氧化,进而提高肿瘤细胞的增殖与迁移能力。
肿瘤免疫微环境是肿瘤细胞周围存在的免疫细胞以及具有免疫调节功能的物质的总称,是肿瘤微环境的重要组成部分[15]。肿瘤免疫微环境在肿瘤发生、发展中发挥重要的调节作用,如促进肿瘤细胞免疫耐受以及肿瘤免疫逃逸等[16]。在调控过程中,肿瘤细胞不是简单地增长,而是在积极主动的调整自身,与免疫细胞相互影响,采取各种策略如限制免疫细胞获取营养物质来使肿瘤免疫延后、改变乃至完全停止,在晚期肿瘤中,这种机制会变得尤其精妙且复杂[4,17]。同样,免疫细胞也会发生改变来适应肿瘤发展,例如恶性肿瘤细胞和调节性T细胞可以驱动T细胞发生衰老。衰老的T细胞表现出不平衡的脂代谢。这种不平衡的脂代谢会改变脂质种类和T细胞中脂滴的积累[18]。通过生物信息学分析发现,脂代谢基因FABP1在精原细胞瘤中具有较高的差异表达,并可能通过调节脂代谢来发挥特定的作用。这一过程同样可能发生在其肿瘤免疫微环境中。笔者首先通过ssGSEA算法评估了FABP1与24种免疫细胞的浸润,发现在高表达FABP1样本中存在更高比例的NK细胞及NK CD56 bright细胞浸润;在低表达FABP1样本中存在更高比例的辅助性T细胞与中央记忆型T细胞浸润。接着利用estimate算法评估精原细胞瘤中各成分情况,发现高表达FABP1的肿瘤中基质成分升高,这与基质细胞能够支持肿瘤生长是一致的;免疫浸润及肿瘤细胞比例均降低,说明免疫微环境在精原细胞瘤不同阶段中产生了较大的变化,在Ⅰ期主要以适应性免疫细胞为主,而在Ⅱ、Ⅲ期以固有免疫细胞为主,且免疫浸润程度显著降低,FABP1可能在这一过程中通过调节脂肪酸代谢发挥关键作用,高表达FABP1的肿瘤细胞与免疫细胞竞争性摄取脂肪酸,从而改变精原细胞瘤的免疫微环境。
FABP1与相关MRGs的Kaplan-Meier生存曲线差异显著,这意味着可以通过针对相关靶点来改善患者预后,延长生存时间。综上所述,本研究认为精原细胞瘤在进展过程中可能发生脂肪酸代谢重编程,肿瘤细胞以FABP1为代表与MRGs相互作用,通过调控肿瘤细胞的脂质运输与细胞内转导信号的合成来增强肿瘤的增殖与迁移能力,在此过程中,肿瘤细胞竞争性摄取脂肪酸,改变肿瘤免疫微环境,进而降低患者预后,这为理解FABP1在肿瘤进展中发挥的作用具有一定的启发。脂代谢在精原细胞瘤细胞中的具体机制,还需要进一步实验研究,探索FABP1作为潜在的诊断标志物与治疗靶点在其中发挥的具体作用以及其与免疫调节的关系,为精原细胞瘤的发生进展提供理论依据。
猜你喜欢
畜牧兽医学报(2023年7期)2023-07-31 08:32:22
昆明医科大学学报(2020年12期)2021-01-26 00:44:02
健康必读·下旬刊(2020年9期)2020-09-13 22:55:19
实用口腔医学杂志(2017年6期)2017-09-19 02:51:42
山东医药(2017年20期)2017-07-01 19:52:07
中南医学科学杂志(2017年1期)2017-01-12 20:25:49
中国学术期刊文摘(2016年1期)2016-02-13 14:05:23
山东医药(2015年44期)2015-02-28 14:29:02
现代检验医学杂志(2015年2期)2015-02-06 02:00:48
沈阳医学院学报(2014年4期)2014-12-27 13:44:30
