
经颅直流电刺激对基于偏定向相干因效脑网络特征的影响
2022-11-24 07:19金荣杭罗志增史红斐
中国生物医学工程学报 2022年4期
金荣杭 罗志增#* 史红斐
1(杭州电子科技大学智能控制与机器人实验室,杭州 310018)
2(浙江大学医学院附属第四医院, 浙江金华 322000)
引言
经颅直流电刺激(transcranial direct current stimulation, tDCS)是一种通过微弱的恒定电流使大脑皮层发生极化从而改变皮层兴奋性的非侵入式神经调控技术[1]。按刺激电流极性的不同,tDCS 可分为阳极刺激和阴极刺激。有研究表明,阳极刺激增强皮质的兴奋性,使神经元膜电位去极化;阴极刺激则降低皮质兴奋性,使静息膜电位超极化[2]。tDCS 具有刺激电流小、极性和位置可调、安全性高、操作方便等优点[3],可调控人脑神经活动、促进脑功能连接重建,实现神经调节机制的可塑性变化。近5年来,功能磁共振成像(functional magnetic resonance imaging,fMRI)和单光子发射断层成像(positron emission computed tomography,PET)等现代成像技术和脑电信号分析手段的不断成熟,使单纯电刺激影响脑活动的研究进入到了更深层次的脑组织分析和神经系统的层面,tDCS 技术再度成为了研究热点[4]。本研究基于偏定向相干(parietal directed coherence, PDC)因果分析方法,构建被试在不同tDCS 范式下的运动想象因效性脑网络模型,利用复杂网络模型理论,提取因效性脑网络的有效特征,为运动想象的脑连接状态以及皮层活跃性评估提供有效测度和理论支撑。
运动想象(motor imagery,MI)具有激活大脑自身细胞可塑性的潜力[5],可加速修复外部肢体与大脑间的神经功能连接,即人体做真实肢体运动与肢体运动想象但无明显动作输出的情况下,大脑的感觉运动皮层区均处于活跃状态。Irma 等[6]研究发现,阳极tDCS 刺激右侧主运动区能够提高左手运动想象的分类识别率。孟献龙等[7]通过经颅直流电刺激主运动区和辅助运动区发现,同时刺激辅助运动区和主运动区能更加有效地改善运动想象脑功能网络特征。Jana 等[8]发现tDCS 阳极刺激主运动区不仅能调节和增强运动想象,还能诱导产生在静息状态下不会发生的自主运动想象。由此推断,tDCS 刺激主运动区能有效调节运动想象。
利用脑网络对脑功能机制和脑认知状态进行基础研究具有重要的意义[9]。目前,已有研究人员应用tDCS 对脑卒中患者的脑网络进行研究。Darkow 等[10]利用tDCS 刺激卒中后失语症患者的M1 区,改善了患者脑功能的整体正常化,增加了脑网络内部通信。Lefebvre 等[11]的研究结果表明,tDCS 结合常规康复训练对于改变慢性卒中后患者感觉运动皮层的脑网络结构具有积极作用。Hordacre 等[12]研究卒中后患者实施tDCS 后脑功能网络连接性变化情况,显示脑连通性增强。大量的研究表明tDCS 对脑网络存在一定影响。因此,基于脑网络,分析研究tDCS 对运动想象的影响具有重要意义。然而目前大多数研究都是基于皮尔逊相关系数或者相位同步构建脑功能网络,分析tDCS 对运动想象的影响,但脑功能网络构建是基于不同脑区间信号强度的统计关系,两个节点间的功能连接是无向的。因此,功能网络不能用于描述脑区间的动态信息流向,仅从脑功能网络的角度分析tDCS 对运动想象的影响势必存在缺陷。而在功能网络基础上发展起来的因效网络,不仅体现了节点间的功能连接,而且反映了节点间的因果关系[13],可将不同大脑功能区间的相互作用描述成动态的信息流。因效性脑网络能更加有效地反应脑网络连接变化情况。
本研究以功能脑区通道信息流入、流出率为局部特征,平均聚类系数和全局效率为全局特征,研究tDCS 对运动想象因效性脑网络特征的影响,探索特征差异的神经生理调节机制。
1 材料与方法
1.1 实验对象
实验被试共16 位,男8 位,女8 位,年龄(24±2)岁,均为右利手,身体状态良好,受教育水平相当。每位被试在刺激前后均要求完成左、右手抓握茶杯两种运动想象范式各20 次的实验任务,并记录相应数据。所有被试已被告知实验具体流程以及潜在风险,并签署了知情同意书。
1.2 设备及参数设置
脑电信号采集设备为博睿康公司的64 导联脑电采集仪,同时采用西班牙Neuroelectrics 公司的4×1高精度经颅电刺激仪进行刺激,脑电信号的采样频率设置为1 000 Hz。
本研究将刺激的目标区域定为右侧主运动区(C4)和左侧主运动区(C3)。刺激方式分为阳极刺激、阴极刺激和伪刺激。范式1:阳极置于C3 点,阴极置于右眼眶上侧(Fp2);范式2:阴极置于C3 点,阳极置于右眼眶上侧(Fp2);范式3:阳极置于C4点,阴极置于左眼眶上侧(Fp1);范式4:阴极置于C4 点,阳极极置于左眼眶上侧(Fp1)。刺激电流的发放时序如图1 所示,阴/阳极tDCS 刺激电流为1 mA,刺激开始和结束均有30 s 淡入淡出时间,电流慢慢由0 变化到1 mA,使被试不会感到过度异样的感觉,1 mA 电流刺激时长20 min。伪刺激组的电流刺激大小设置为1 mA,时长2 min,具有相同时长的淡入淡出时间,使得被试在与真实刺激的感觉上没有差异。刺激电极在10-20 国际标准导联分布中的物理位置如图2 所示。
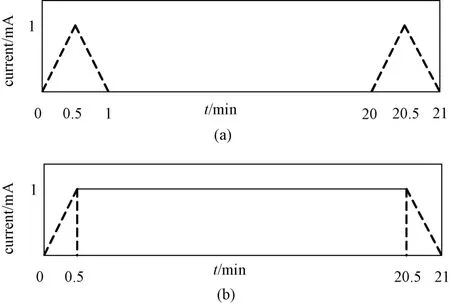
图1 刺激电流发放时序。(a) 伪tDCS 电流(b) 阳极/阴极tDCS 刺激电流Fig.1 Timing of stimulus current discharge. (a)sham tDCS current (b) cathode/ anode tDCS current
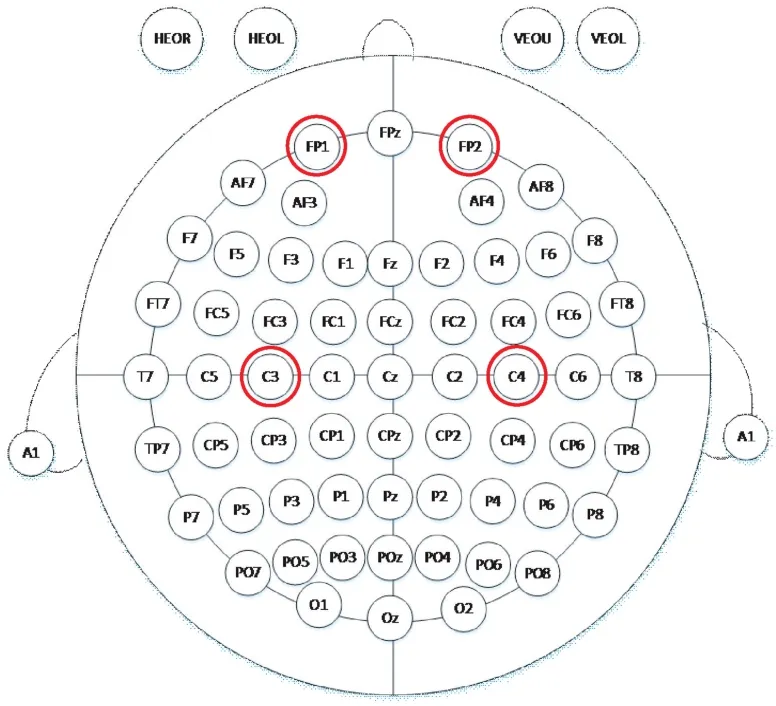
图2 刺激电极的位置分布Fig.2 Position distribution of stimulation electrodes
在运动想象脑电信号采集过程中,眨眼难以避免, 眼球或眼皮运动等引起的眼电信号(electrooculogram, EOG)沿颅骨传播与脑电信号发生融合,使EEG 产生伪迹。为滤除眼电伪迹,每轮实验中,脑电采集设备需要连续同步采集受试者的EEG 及额外独立的EOG 信号,以便EEG 预处理消除伪迹。
1.3 运动想象实验范式
文献[14]的研究表明,视觉和听觉引导引导运动想象能提高运动想象皮层活跃度,增强运动想象效果。为了尽可能排除视觉刺激对运动想象的影响,本研究选用左、右手抓握茶杯的视频异步引导被试进行左、右手运动想象任务。以被试执行右手运动想象任务为例,具体实验模式的实施时序如图3 所示。
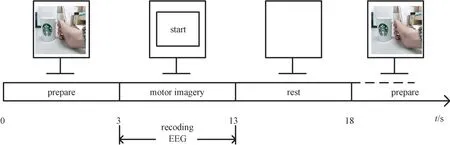
图3 运动想象实验流程Fig.3 Flow chart of motor imagery experiment
实验时被试坐在扶手椅中,显示屏放距离被试1.5 m,与其眼部水平的位置。整个实验过程中被试正视显示屏。当屏幕抓握茶杯的视频时,提示被试开始准备运动想象,准备时间3 s;当屏幕上出现“start”字样时,被试开始运动想像任务,运动想象时间10 s。当屏幕变为白屏时,提示被试结束运动想象进入5 s 的放松状态。每位被试重复运动想象实验10 次。
1.4 因效性脑网络构建
根据多导联头皮EEG 构建因效性脑网络,首先需要选取导联数目并将各导联作为一个节点,之后采用因果分析方法量化各导联信号间的相互作用关系,然后把量化结果作为各导联间的有向连接关系,从而建造大脑的因效网络。格兰杰因果性分析[15](Granger causality analysis, GCA)是其中一种因果分析方法。为分析不同脑区间的动态信息流向,本研究选用在GCA 基础上发展起来的偏定向相干因果分析方法构建因效网络。
为了实现对运动想象的因效性网络构建,以EEG 信号为基础,对给定长度为N的EEG 时间序列x(n) ∈RM,利用p阶多变量自回归模型(multivariate autoregressive,MVAR)求系数矩阵Ar,其具体数学表达式为
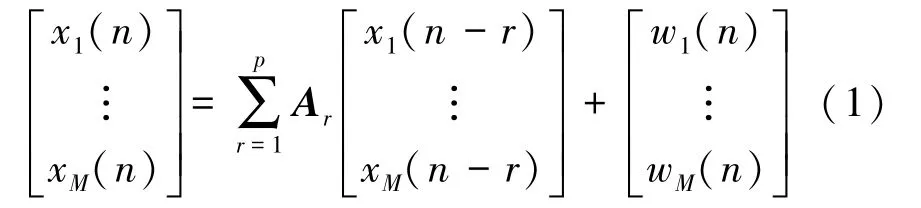
式中,w表示白化噪声矢量,RM表示导联数目为M的EEG 数据段。式(1)计算所得Ar为M×M维系数矩阵,其具体数学表示形式为
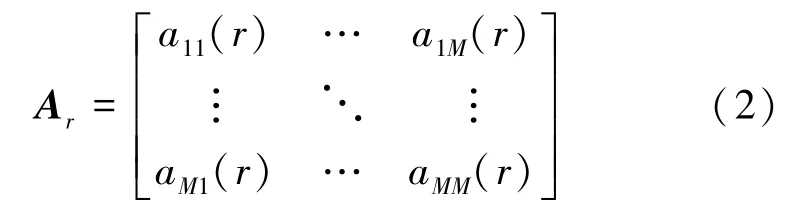
式中,r=1,2,…,p。aij(r) 表示区域i到区域j的线性影响。
将Ar转化到频域:
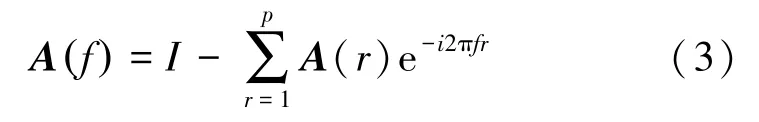
模型阶数p越大,其描述对象样本的能力越强。但是阶数越高,参数越多,容易造成过拟合的现象。一般由信息量准则(Akaike's information criteria, AIC)与贝叶斯信息准则(Bayes information criterion, BIC)综合确定阶数。为了简化计算,本利用AIC 准则获取MVAR 模型的滞后阶数p,AIC的定义为
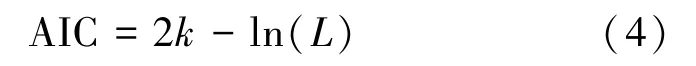
式中,L为模型参数的似然函数,k表示参数个数。AIC 准则通过计算最小AIC 值找到最优阶数,本研究计算获得阶数p=9。从区域j到区域i的PDC 网络连接参数定义为

式中,Aj(f) (j=1,2,…,M)为矩阵A(f)的列向量,Aij(f)为矩阵A(f)第i行,第j列的元素。为根据MAVR 参数进行变换获得的连接测度,其具体表示如下:

一般而言,计算的是某一段频率下,各个通道间因果性的平均PDC 值,即:
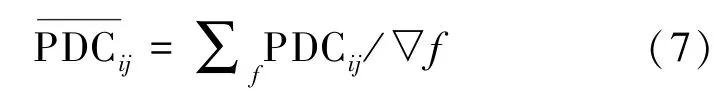
徐佳琳[16]的研究表明,大脑在进行肢体运动想象时,运动想象EEG 节律(mu 节律)会在相应的脑区发生变化,表现出显著特征。本研究选择mu 节律的频带特征运算。
1.5 阈值选择
不同的阈值将对因效性脑网络的拓扑结构产生直接的影响,因此阈值的选择关系到脑网络构建的好坏。阈值选择过大有可能遗漏一些重要的连接边;选择太小则会增加不必要的连接边。由于阈值对构建的脑网络有直接影响,因此不管是功能性脑网络还是因效性脑网络,阈值的选择都是重点问题。基于脑网络的特性,选择阈值需要遵循以下3个原则[17-18]:
1)阈值的选择应该尽量使得构建的脑网络中不存在孤立的节点,以此来保证脑网络的完整性;
2)阈值的选择应该使得脑网络具有较高的全局效率和局部效率,保证脑网络的小世界特性不被破坏;
3)在对比实验中,阈值的选择应该在保证脑网络小世界特性的前提下尽量相同,减少阈值差异对脑网络特征的影响。
本研究设置阈值步长为0.002,基于上述原则最终选择的阈值如表1 所示。

表1 不同范式刺激下左、右手运动想象因效性脑网络阈值δ 选取Tab.1 Selection of threshold δ of effective brain networks due to effects of left and right hand motor imagination under different paradigms
确定δ阈值后,对因效性脑网络进行阈值处理,获得0-1 关联矩阵,其表示如下:
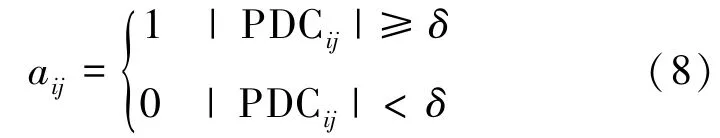
aij =1 表示通道i和通道j之间具有较强的关联性。
1.6 测度选择
因效性脑网络构建完成后,利用复杂网络的测度方法可以得到脑功能网络的特征参数。经颅直流电刺激前后,在网络特征参数上的异同,可以反映出刺激对大脑内部连通特性的影响[19]。
1.6.1 通道信息流入、流出率
脑功能网络的连通增率(connectivity increment rate)是以功能皮质区域为中心的区域特征网络,它反映了网络位置和连通性的变化,并减少了网络信息的丢失[20]。本研究基于因效性脑网络和连通增率提出了通道信息流入、流出增率,反映通道信息流向的变化情况,其表示如下:
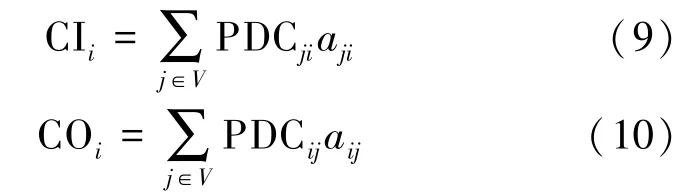
式中, CI 表示通道i的信息流入率,CO 表示通道i的信息流出率,PDCij表示通道i和通道j间因果性的PDC 值,aij如式8 所示,V表示所选功能脑区通道的集合。
1.6.2 聚类系数
聚类系数是衡量脑功能网络内部集群特性和连接紧密程度的重要参数,表示网络中某节点的邻接点又互为邻居的可能性,聚类系数越大,脑功能网络内部节点连接越紧密。如网络中节点i的聚类系数是指与节点i相邻的节点中实际存在的边数Ei与邻居节点中可能存在的最多边数之比,定义为:
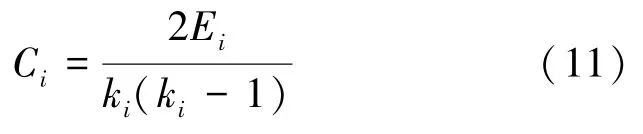
式中,Ei为节点i邻居间实际连接的边数,ki表示节点i具有的邻节点数,ki(ki-1)/2 表示在这个ki邻节点之间最多可能有的边数。由于复杂网络中节点数比较多,因此不去研究网络中每个节点的聚类系数,而是分析整个网络的平均聚类系数,对无权网络中所有节点的聚类系数取平均就可以得到整个网络的平均聚类系数,定义为

式中,N为整个网络中的节点数。
1.6.3 全局效率
全局效率体现网络整体的信息传递能力,是用来描述网络整体属性的,相比于局部效率,全局效率更能够反映网络的实际情况。全局效率越大,网络的连通性也好,各节点间信息传输效率更快,定义为
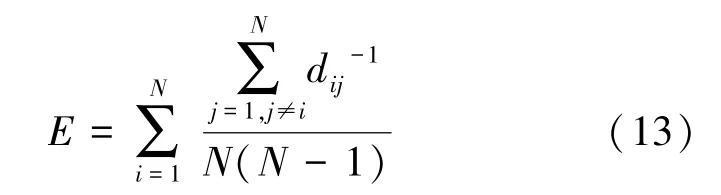
式中,N为整个网络中的节点数,dij表示节点i与j之间的距离,即连接两个节点的最短路径的边数。
1.7 统计学分析
基于t检验选择通道信息流入、流出率,聚类系数和全局效率等因效性脑网络的特征参数,对被试伪刺激和不同tDCS 实验范式刺激后进行显著性分析。本研究选用α=0.05 的统计显著水平,通过计算检验统计量,查相应界值表确定P值。若P>0.05 则称差异无统计学意义;若P<0.05,则认为差异具有统计学意义。
2 结果
经颅直流电刺激对运动想象脑功能网络因果效应的影响分析时,采用运动相关Fz、FCz、Cz、C1、C2、C3、C4 通道EEG 信号。
2.1 运动脑区通道信息流入、流出率对比分析
根据左、右手抓握茶杯的运动想象,分别选择与左臂运动密切相关的C4 电极和与右臂运动密切相关的C3 电极为区域中心,运动相关电极Fz、FCz、Cz、C1、C2、C3、C4 为运动脑区通道集合,进行通道信息流出、流入率对比分析,运动脑区通道信息流入率、流出率对比如图4 所示。图4(a)结果显示,伪刺激和阳极刺激C4 后的运动脑区C3 通道信息的流出/流入率分别为0.161±0.009/0.209±0.008,0.189±0.007/0.240±0.016,差异较为显著(P<0.05)。阴极刺激C4 后的C3 通道信息流入率为0.170±0.009,与伪刺激相比差异较为显著(P=0.018),C3 通道信息的流出率为0.153±0.012,与伪刺激相比差异不大(P>0.05)。由图4(b)可知,被试在进行左手运动想象任务时,伪刺激和阳极刺激C4 后的运动脑区C4 通道信息的流出/流入率分别为0.142±0.014/0.193±0.013,0.223±0.025/0.258±0.023,差异显著(P<0.01)。阴极刺激C4后的C4 通道信息流出率为0.109±0.009,与伪刺激相比差异显著(P=0.009),C4 通道信息流入率为0.184±0.008,与伪刺激相比差异不大(P>0.05)。由此可以推断阳极刺激能提高通道信息交流的频率,阴极刺激则会降低对应通道的信息流出,进而导致同一功能脑区中的其他通道的信息流入下降,一定程度上降低了通道间的信息交流频率。图4(c)和图4(d)的结果与图4(a)和图4(b)的结果趋于一致。
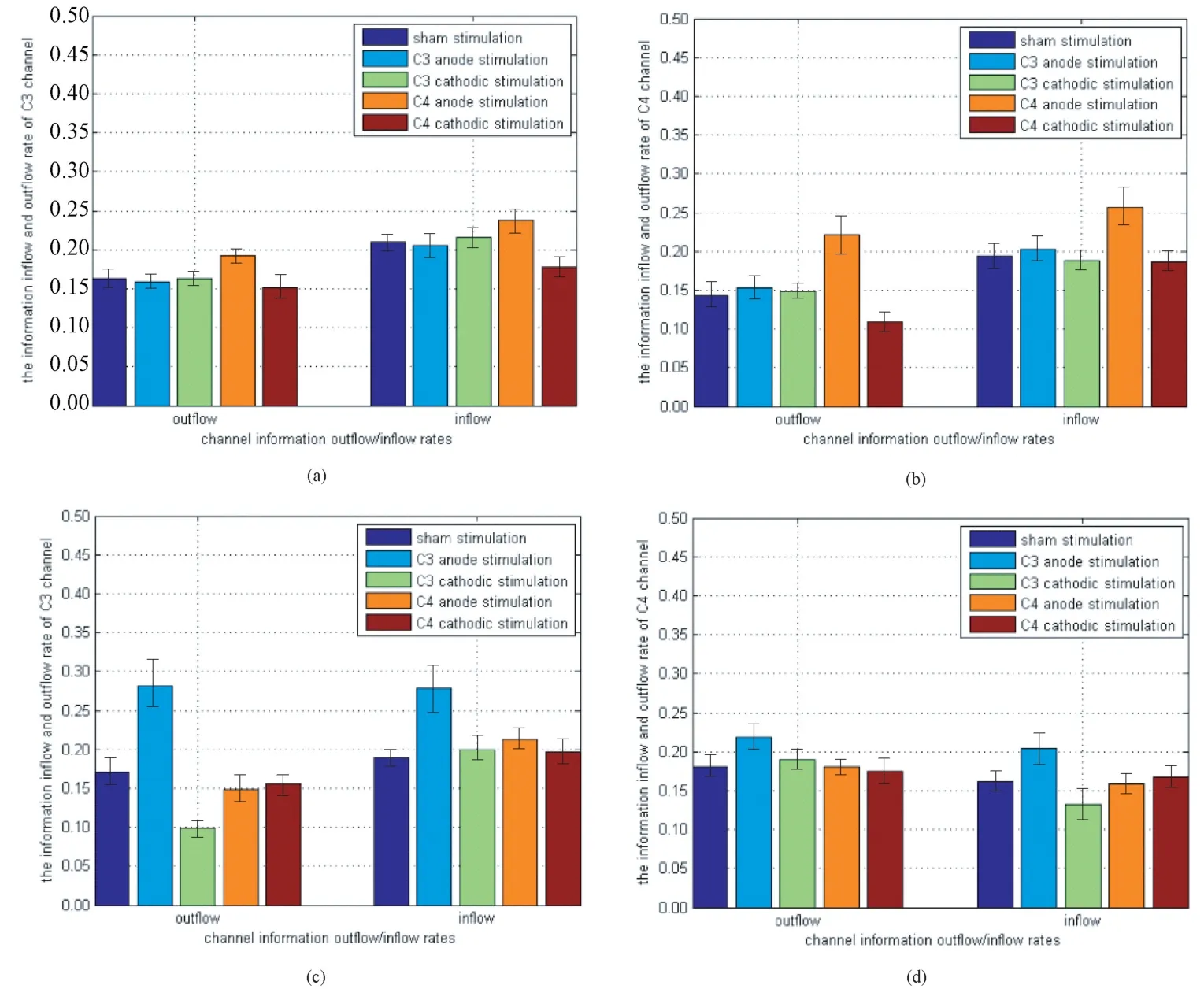
图4 运动脑区通道信息流入、流出率对比。(a)左手运动想象C3 通道;(b)左手运动想象C4 通道;(c)右手运动想象C3 通道;(d)右手运动想象C4 通道Fig.4 Comparison of channel information inflow and outflow rates in motor brain regions. (a) Left hand motor imagery C3 channel; (b) Left hand motor imagery C4 channel; (c) Right hand motor imagery C3 channel; (d)Right hand motor imagery C4 channel
2.2 平均聚类系数对比分析
图5 是左、右手运动想象在不同范式tDCS 刺激后的平均聚类系数C分析结果。被试执行左手运动想象任务时,阳、阴极刺激C4 后的平均聚类系数分别为0.817±0.021、0.356±0.037,与伪刺激的平均聚类系数0.585±0.046 相比,差异均十分显著(P<0.01)。而阴、阳极刺激C3 则与伪刺激相比差异不大(P>0.05),阴、阳极刺激C3 后,被试执行右手运动想象任务的结果与左手运动想象任务趋于一致。由此可以看出阳极刺激与想象任务相关的功能脑区能增加不同脑区之间的紧密程度,阴极则相反。
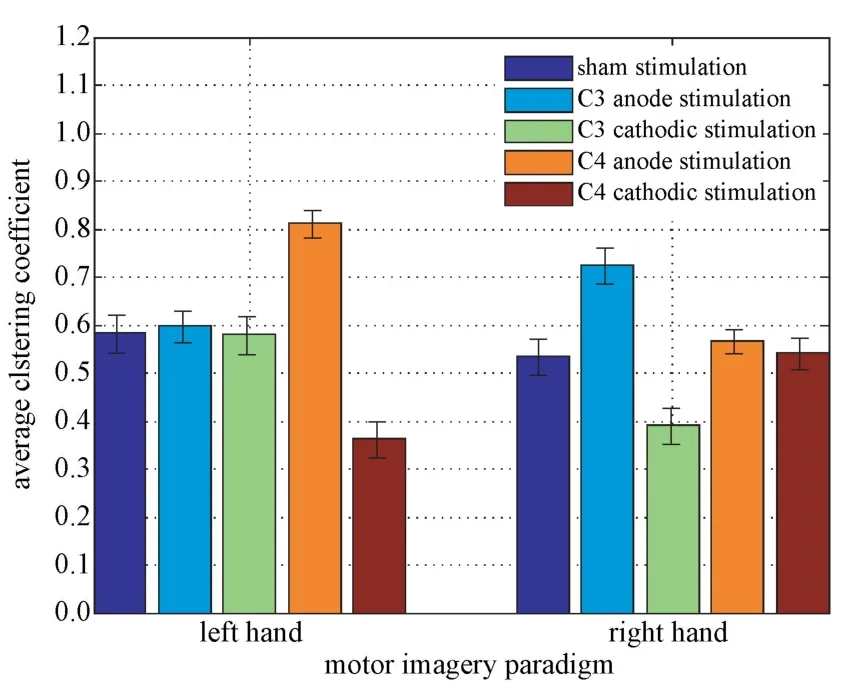
图5 左、右手运动想象任务的平均聚类系数Fig.5 The average clustering coefficient of left and right hand motor imagery tasks
2.3 全局效率对比分析
左、右手运动想象在不同范式tDCS 刺激后的全局效率如图6 所示。从中可以看出被试在执行左手运动想象任务时,阳、阴极刺激C4 后的全局效率分别为0.491±0.091、0.252±0.024,与伪刺激后的全局效率0.347±0.031 相比,差异显著(P<0.01)。阴、阳极刺激C3 则与伪刺激相比差异不大(P>0.05)。被试在进行右手运动想象任务时,阳、阴极刺激C3 后的全局效率分别为0.492±0.025、0.249±0.011,与伪刺激后的全局效率0.359±0.028 相比,差异显著(P<0.01),而阴、阳极刺激C4 则与伪刺激相比差异不大(P>0.05)。由此可以推测,在刺激与想象任务相关功能脑区时,tDCS 确实具有阳极激活而阴极抑制皮层活性的功能。
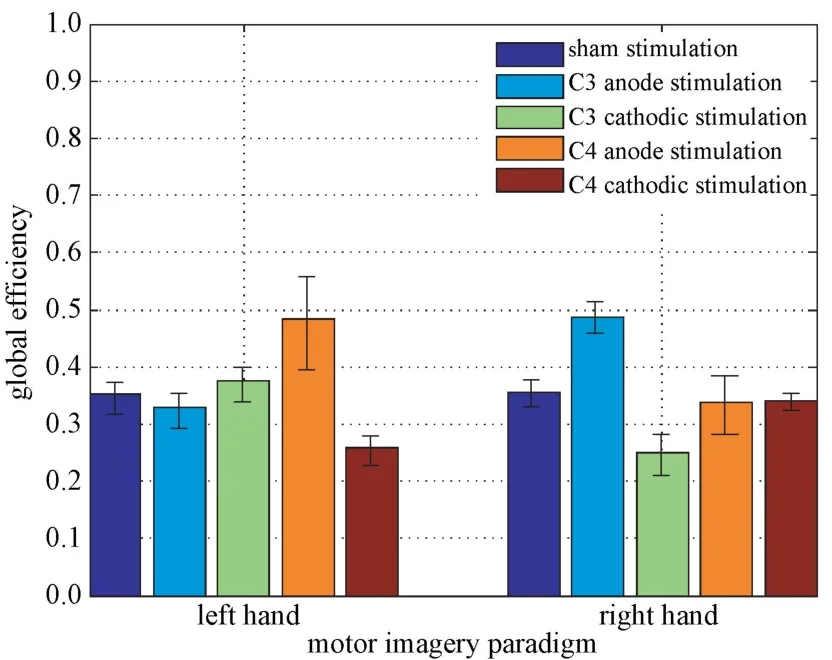
图6 左、右手运动想象任务的全局效率Fig.6 Global efficiency of left and right hand motor imagery tasks
3 讨论
Allman 等[21]研究发现tDCS 阳极刺激脑卒中患者健侧的初级运动区,3 个月后经上肢运动研究量表和沃尔夫运动功能测试得分显示真刺激组的运动功能恢复优于伪刺激组,MRI 显示初级运动区在刺激后显著激活。Notturno 等[22]研究脑缺血大鼠实验发现,经过tDCS 阴极刺激后的大鼠脑梗塞面积小于对照组,而且记录到的去极化电位明显少于对照组,表明阴极减弱降低神经元去极化。Stagg等[23]通过功能性磁共振造影研究发现,阳极tDCS短期的刺激效应显示,在运动辅助区和受刺激的M1区有明显的活动性提高。Grami 等[24]对比tDCS 和伪tDCS 刺激下脑功能网络连接情况发现,tDCS 能够控制运动执行和心理想象,调节远端皮层区域的活动。以上证据提示tDCS 电流影响神经元的静息膜电位,阳极电流促进神经元电位去极化而阴极引起膜电位超极化改变,增强大脑皮质的兴奋性或抑制其活动。
针对功能性脑网络不能用于描述脑区间的动态信息流向的问题,本研究基于偏定向相干构建了因效性脑网络,并在因效性脑网络的基础上结合连通增率提出了新的特征,功能脑区通道信息流入、流出率,以此作为局部特征,结合平均聚类系数和全局效率等两个全局特征,从多个层面分析tDCS 对运动想象因效脑网络特征的影响,通过4 种tDCS 范式实验,阳极刺激提升活跃性,阴极刺激抑制活跃性,阳/阴极tDCS 刺激均能提高特征区分度,且与运动想象任务相关。
实验结果表明,tDCS 刺激被试运动想象任务功能对应的运动脑区时,阳极刺激会提高刺激通道的信息流入、流出率,提高平均聚类系数和全局效率;阴极刺激运动脑区,刺激通道的信息流出率下降,平均聚类系数和全局效率降低。值得注意的是,实验被试执行左手运动想象,tDCS 阳/阴极刺激C4 通道对C4 通道信息流入/流出率的影响效果远大于对C3 通道信息流入/流出率的影响。由此推断,阳极tDCS 能够提高刺激脑区皮层神经元的兴奋性,运动想象能激活相关功能脑区的神经元使其冲动并向其他脑区传导,若被试tDCS 阳极刺激通道与运动想象任务相关脑区通道一致,则能充分激活相关脑区皮层的神经元,提高皮层活跃性,加强该脑区与其他脑区间的信息交流,进而使其相关功能脑区信息交流更加频繁。
实验选择的被试均为健康的青年,但是对于年龄不同、身体状况存在差异的人群,其大脑结构和活跃性不完全一致,因此实验的结论存在一定的局限性,无法适用于所有情况。在未来的研究中可以扩大被试人群的类别,使研究结果更具普适性。
通过对比分析tDCS 对运动想象因效脑网络特征的影响研究,探索特征差异的神经生理调节机制,为以后tDCS 对运动想象脑电信号特征影响的深入研究提供了一定的理论支撑和参考价值。脑电信号特征的差异无疑将为提高运动想象-脑机接口识别率创造条件。
4 结论
本研究针对4 种范式刺激和伪刺激模式下,左、右手运动想象任务的因效性脑网络特征进行对比分析,判断tDCS 刺激对运动想象的影响,以通道信息流入、流出率作为因效脑功能网络的局部特征,以平均聚类系数和全局效率作为因效脑功能网络的全局特征,探索不同tDCS 刺激方式对运动想象脑功能网络影响的具体机制。研究结果表明,阳极tDCS 刺激能有效激活该脑区皮层的活跃性,使脑区间信息交流更加频繁,增加脑网络的聚集程度,增强脑网络的连通性,提高脑网络的全局效率。阴极tDCS 刺激则会抑制脑区皮层的活跃性,降低脑区信息的流出,减少脑网络的聚集程度,降低脑网络的全局效率。未来的研究希望可以发现tDCS 刺激是否能对复杂的运动想象仍具有显著的影响效果,同时将其运用在运动想象的识别中是否能对识别率产生显著的提升效果。
猜你喜欢
化工管理(2022年14期)2022-12-02
安徽医科大学学报(2022年11期)2022-11-01
有色设备(2022年2期)2022-08-06
化工管理(2021年7期)2021-05-13
陶瓷学报(2021年1期)2021-04-13
军民两用技术与产品(2021年10期)2021-03-16
浙江大学学报(理学版)(2021年1期)2021-01-26
电子制作(2018年12期)2018-08-01
健康管理(2017年3期)2017-04-20
