
七彩神仙鱼免疫球蛋白超家族成员jam-a和cd22基因克隆及表达分析
2022-11-23 07:39:12马腾飞高建忠陈再忠
淡水渔业 2022年6期
刘 鑫,温 彬,3,马腾飞,缪 琳,高建忠,3,陈再忠,3
(1.上海海洋大学,农业农村部淡水水产种质资源重点实验室,上海 201306;2.上海海洋大学,上海水产养殖工程技术研究中心,上海 201306;3.上海海洋大学,水产科学国家级实验教学示范中心,上海 201306)
七彩神仙鱼(Symphysodonaequifasciatus)隶属于鲈形目(Perciformes)慈鲷科(Cichlidae)盘丽鱼属(Symphysodon),原产于亚马逊河流域,因其体色艳丽,条纹多样等特征,被誉为“观赏鱼之王”[1]。该物种具有独特的亲代抚育行为,即仔鱼孵化后的四周内以亲本体表分泌的粘液为营养和免疫来源[2-3]。BUCKLEY等[4]发现七彩神仙鱼粘液中含有大量营养和非营养因子,包括免疫球蛋白、激素和必需离子。免疫球蛋白M(IgM)在亲鱼产卵后逐渐增加,在幼鱼达到自由游泳阶段时达到峰值,然后下降到最初的水平,这种变化类似于哺乳动物在母乳喂养期间为子代提供的被动免疫[5-6]。
免疫球蛋白超家族(IgSF)由至少编码一个免疫球蛋白(Ig)结构域的基因组成,其成员具多种细胞功能,如细胞表面抗原受体、细胞粘附分子和细胞因子受体等[7]。连接黏附分子(JAMs)是IgSF成员,其中,JAM-A分子参与细胞旁通透性、白细胞跨内皮迁移、上皮间质转化和血小板活化等生物过程[8-9]。DU等[10]克隆了草鱼(Ctenopharyngodonidella)Gcjam-a,发现Gcjam-a可能是呼肠孤病毒的受体,在病原体免疫防御中发挥作用。cd22属唾液酸结合免疫球蛋白样凝集素(Siglecs),也是IgSF成员,主要表达于B细胞,调节其存活、增殖、信号转导以及抗体的产生[11]。本实验组前期对七彩神仙鱼育幼期皮肤转录组分析发现,jam-a和cd22基因表达分别在育幼期间和育幼结束后显著上调,暗示着二者可能参与了亲子的免疫防御,且扮演不同的角色。
嗜水气单胞菌(Aeromonashydrophila)可导致多种疾病的发生,是淡水养殖鱼类常见的病原菌[12-13],气单胞菌属会导致七彩神仙鱼患肠炎病[14]。本实验选取嗜水气单胞菌[15]为感染模型,从七彩神仙鱼皮肤中克隆获得了jam-a和cd22,并通过荧光定量PCR法分析了其在组织中的表达量,以及细菌刺激后各组织中的表达模式,以期初步了解jam-a和cd22在七彩神仙鱼免疫系统中可能的作用,为丰富IgSF分子在硬骨鱼类中的免疫功能提供基础。
1 材料与方法
1.1 实验材料
健康的七彩神仙鱼(体长10~13 cm,体质量80~100 g)取自上海海洋大学滨海养殖基地,暂养1周[(28±0.5) ℃,pH 7.8±0.5,充氧]。嗜水气单胞菌菌株来自上海海洋大学病原库。
1.2 实验方法
1.2.1 七彩神仙鱼总RNA提取与cDNA的合成
七彩神仙鱼暂养后,选6尾健康的七彩神仙鱼,解剖收集肝脏、心脏、脾脏、头肾、皮肤、肠道、肌肉、鳃、脑和性腺,共10个组织,使用TRIzol(Invitrogen)提取各组织中的总RNA,1%琼脂凝胶电泳测定RNA完整性,其浓度和纯度用分光光度计测定。根据PrimeScriptTMRT Master Mix试剂盒(TaKaRa)说明书合成cDNA。
1.2.2sajam-a和sacd22基因克隆
根据前期七彩神仙鱼皮肤转录组获得jam-a和cd22 cDNA部分序列,利用Primer 5设计开放阅读框(open reading frame,ORF)特异性引物(表1),分别用于克隆验证上述基因的ORF序列。以七彩神仙鱼皮肤中的cDNA为模板,利用PCR方法及上述引物对sajam-a和sacd22进行扩增。扩增体系25 μL:cDNA 1 μL,正反向引物各1 μL,金牌 Mix green(TsingKe)22 μL。反应条件:98 ℃,2 min;98 ℃,10 s,56 ℃,10 s,72 ℃,10 s,34个循环;72 ℃,1 min。PCR产物以1%的琼脂凝胶电泳测定并切胶回收,送至上海生工生物公司进行测序。
1.2.3sajam-a和sacd22基因生物信息学分析
使用NCBI上的ORF Finder查找sajam-a和sacd22的ORF区,并获得相应的氨基酸序列;应用ExPASy工具(http://web.expasy.org/compute_pi/)预测等电点(pI)及分子质量(Mw); SignalP 4.1和TMHMM 2.0分别预测信号肽和跨膜结构域;SMART分析蛋白结构域;Blast程序搜索与其他物种的jam-a和cd22基因同源性分析;使用Clustal-X和MEGA 5.0软件构建Neighbor-Joining系统发育树。
1.2.4sajam-a和sacd22基因的组织表达分析
根据sajam-a和sacd22的ORF区设计荧光定量PCR特异性引物(表1)。以七彩神仙鱼各组织的cDNA为模板,β-actin为内参基因,利用荧光定量RT-PCR的方法测定各组织中sajam-a和sacd22的表达模式。反应体系20 μL:2×T5 Fast qPCR Mix(TsingKe) 10 μL,cDNA 模板1.6 μL,正反向引物各0.8 μL,ddH2O 6.8 μL。反应条件:95 ℃,1 min;95 ℃,10 s,56 ℃,5 s,72 ℃,10 s,40个循环;95 ℃,5 s;65 ℃,5 s;95 ℃,15 s。采用2-ΔΔCt法计算sajam-a和sacd22在组织中的相对表达量。
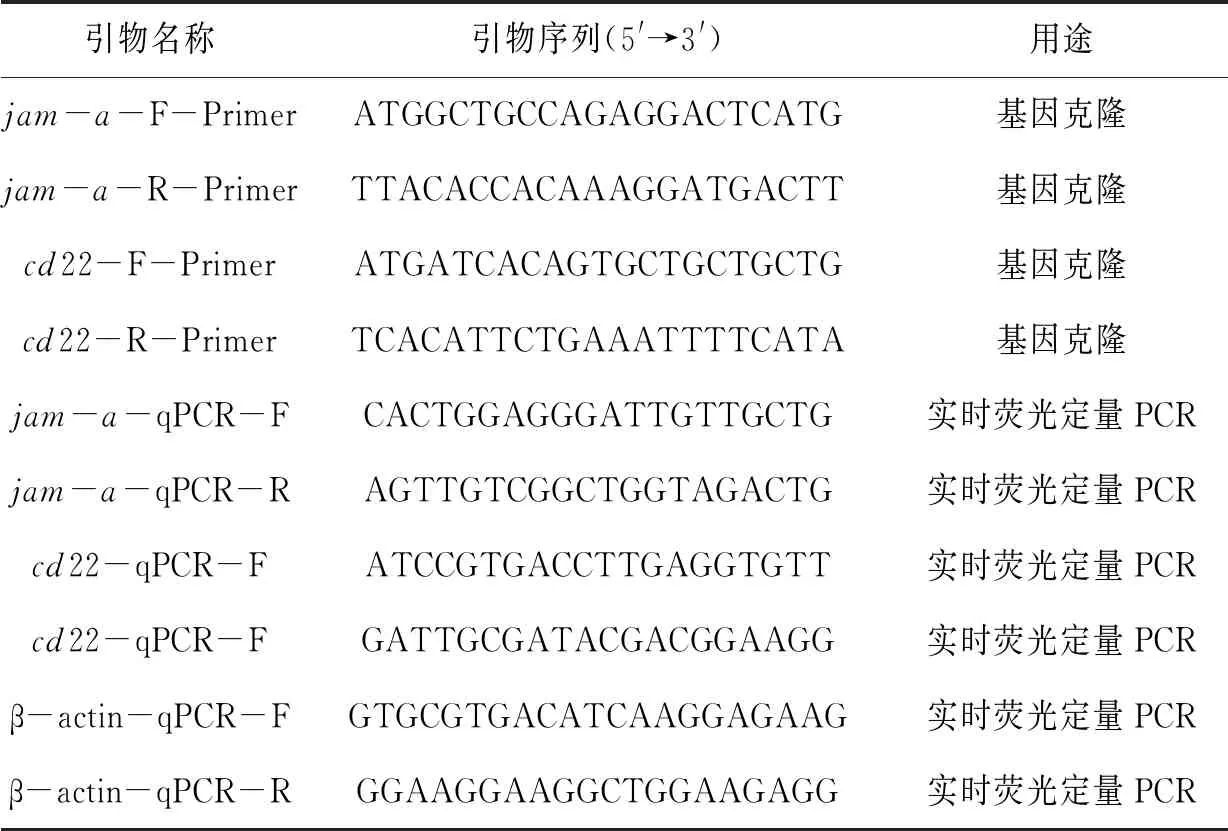
表1 引物信息
1.2.5sajam-a和sacd22基因免疫刺激后的表达变化
将健康的七彩神仙鱼分为2组,每组36尾鱼,分别饲养于已曝气的水中。实验组腹腔注射1x107cfu/mL的菌液(2 mL/尾),对照组注射等量的PBS缓冲液。在注射后的0、6、12、24、48、72 h后,分别从两组随机各取6尾鱼,对其进行解剖取样,主要取样组织为皮肤、肠道、头肾和脾脏。提取各组织中的总RNA,反转录得到cDNA。使用qRT-PCR测定免疫刺激后各组织不同时间点sajam-a和sacd22的表达模式。反应体系、程序及表达量计算方法同上述组织表达。采用SPSS软件进行单因素方差分析。
2 结果与分析
2.1 sajam-a和sacd22的克隆及序列分析
七彩神仙鱼sajam-a和sacd22的ORF序列长度分别为903 bp和1 032 bp(图1),分别编码300和343个氨基酸(aa)。sajam-a预测蛋白理论Mw为32.51 kDa,PI为8.84,其N基端存在1个信号肽(aa 1~20),随后是1个Ig(aa 26~126)和1个Ig C2结构域(aa 128~213),羟基末端是1个跨膜结构域(aa 236~257)(图1A)。sacd22蛋白理论Mw为38.12 kDa,PI为8.82,其氨基酸序列存在1个信号肽(aa 1~18),2个Ig结构域(aa 26~128 134~202),1个Ig C2结构域(aa 219~278)和1个跨膜结构域(aa 297~319)(图1B)。

图1 七彩神仙鱼sajam-a(A)和sacd22(B)的cDNA序列及推导的氨基酸序列
2.2 sajam-a和sacd22氨基酸序列比对及系统进化树分析
BLAST比对发现,鱼类中jam-a氨基酸序列有较高的相似度(51.83% ~ 93.00%),sajam-a的氨基酸序列与尼加拉瓜湖始丽鱼(Archocentruscentrarchus)的相似度最高,为93.00%;尼罗罗非鱼(Oreochromisniloticus)次之,为71.76%(图2A)。cd22氨基酸序列在鱼类中也有较高的相似度(71.72%~92.71%),sacd22的氨基酸序列与尼加拉瓜湖始丽鱼的相似度最高,为92.71%(图2B)。各物种jam-a和cd22氨基酸序列构成的系统进化树表明,哺乳类和鱼类分别单独成簇,七彩神仙鱼jam-a和cd22均与尼加拉瓜湖始丽鱼的进化距离最近(图3)。
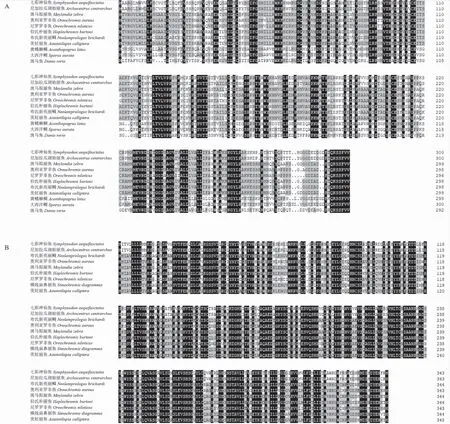
图2 jam-a(A)和cd22(B)氨基酸的多重序列比对
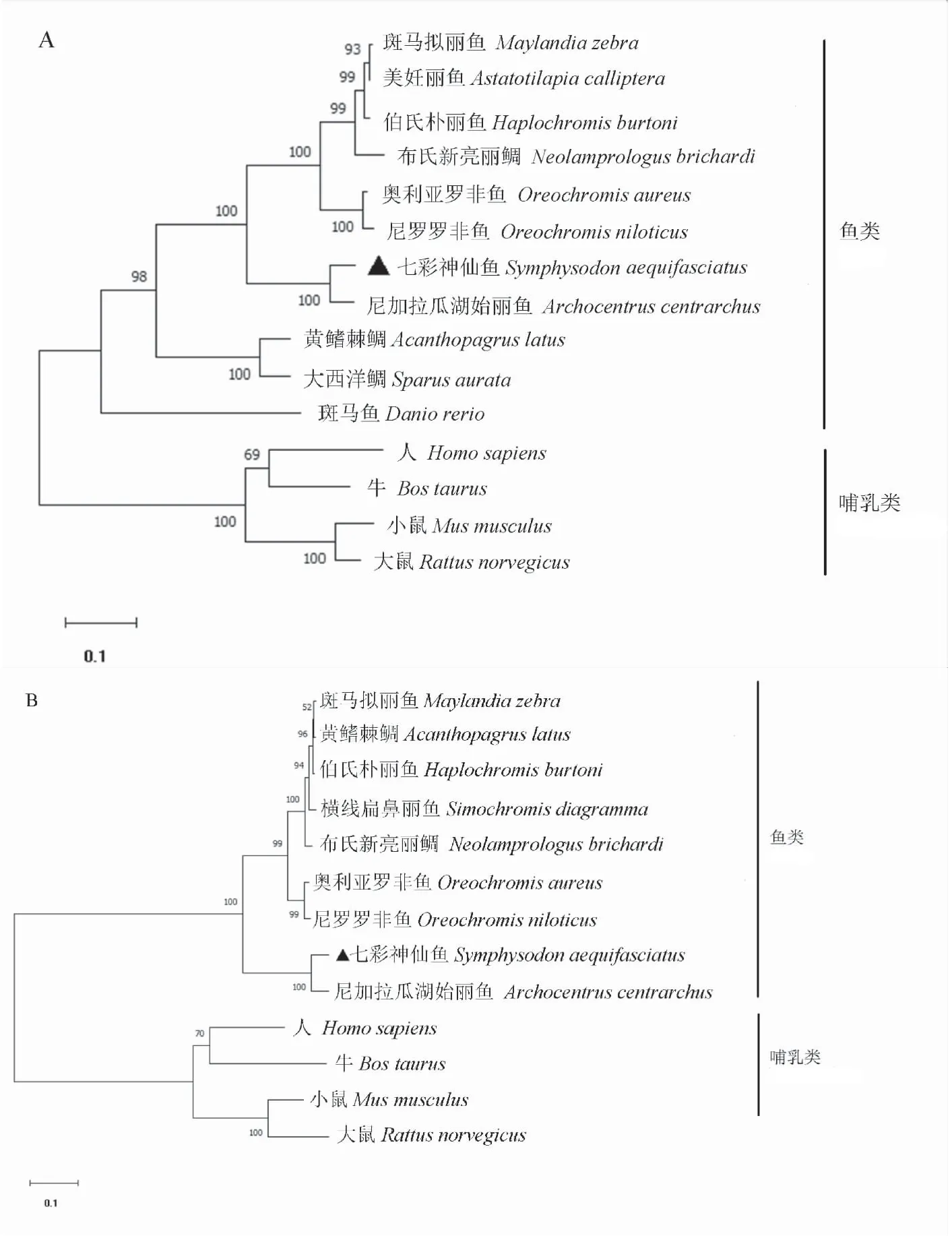
图3 基于NJ法构建的jam-a(A)和cd22(B)系统进化树
2.3 sajam-a和sacd22的组织表达模式
如图4所示,sajam-a和sacd22在健康七彩神仙鱼的10个组织中都有表达,并存在显著的组织差异性。sajam-a在鳃中表达量最高,其次是肝脏、头肾、肠道和皮肤,在肌肉中表达量最低(图4A)。sacd22在脾脏和头肾表达量较高,同样在肌肉中几乎不表达(图4B)。
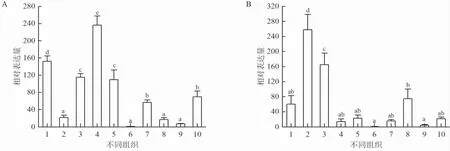
图4 sajam-a(A)和sacd22(B)在七彩神仙鱼不同组织中的表达特征
2.4 sajam-a和sacd22基因免疫刺激的表达变化
嗜水气单胞菌感染七彩神仙鱼后,定量分析测定七彩神仙鱼肠道、头肾、皮肤和脾脏组织中jam-a和cd22的时序表达模式。结果发现,在肠道、头肾、皮肤和脾脏中,sajam-a的表达量均在6 h时显著上升,且在肠道和头肾中6 h达到峰值,分别为对照组的5.30倍和5.60倍,12 h开始降低(图5A和B);在皮肤和脾脏中12 h达到峰值,分别为对照组的7.05倍和8.57倍,在24 h降低,48 h达到最低值,72 h恢复正常水平(图5C和D)。肠道和头肾中,sacd22的表达量在6 h显著上升,在头肾中72 h达到峰值(图6A和B);脾脏组织sacd22的表达水平在48 h显著上调,72 h达到峰值(图6D);皮肤组织sacd22的表达水平在12 h显著上调,48 h达到峰值(图6C)。
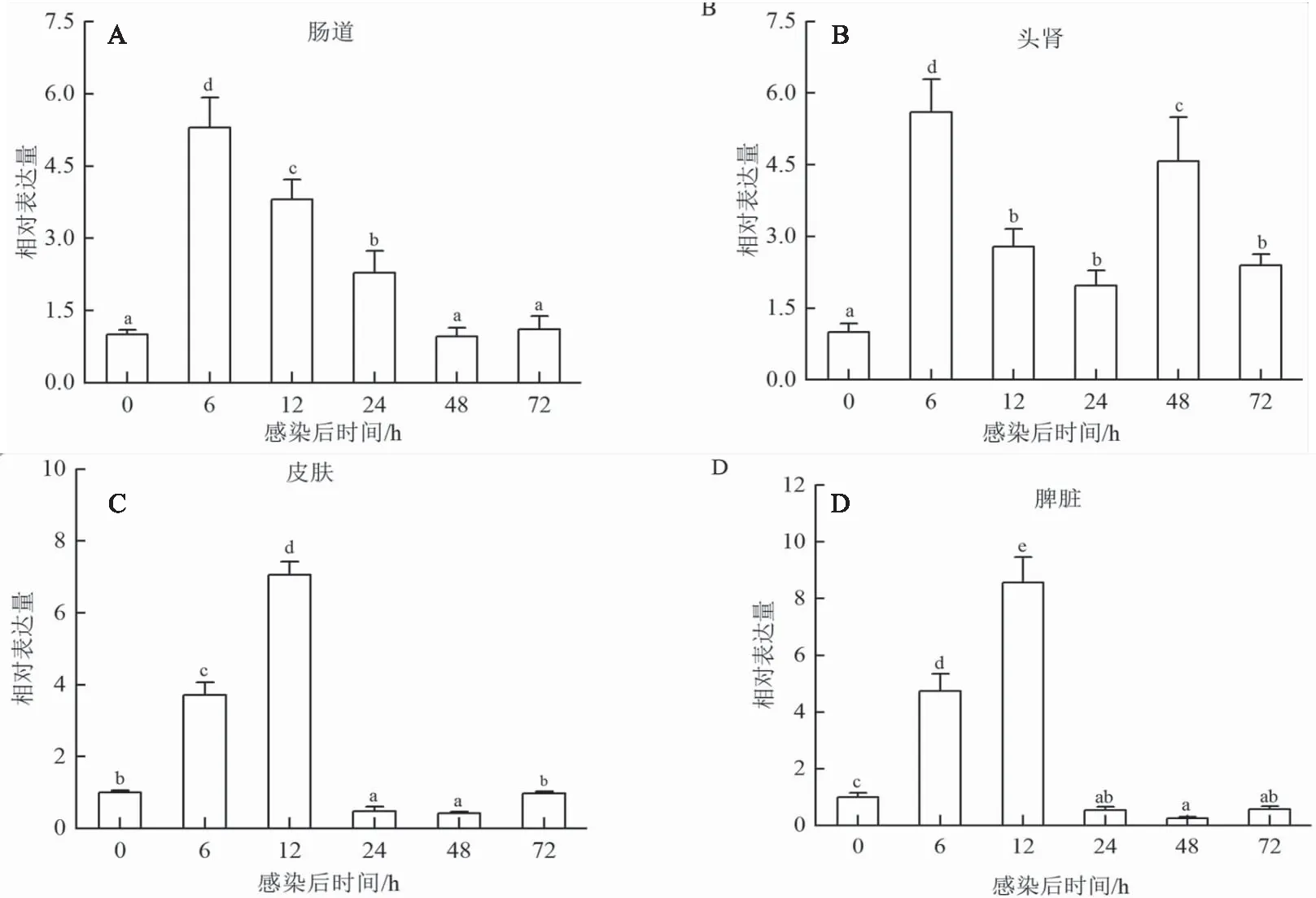
图5 嗜水气单胞菌感染后sajam-a在七彩神仙鱼不同组织中(A-D)的表达分析
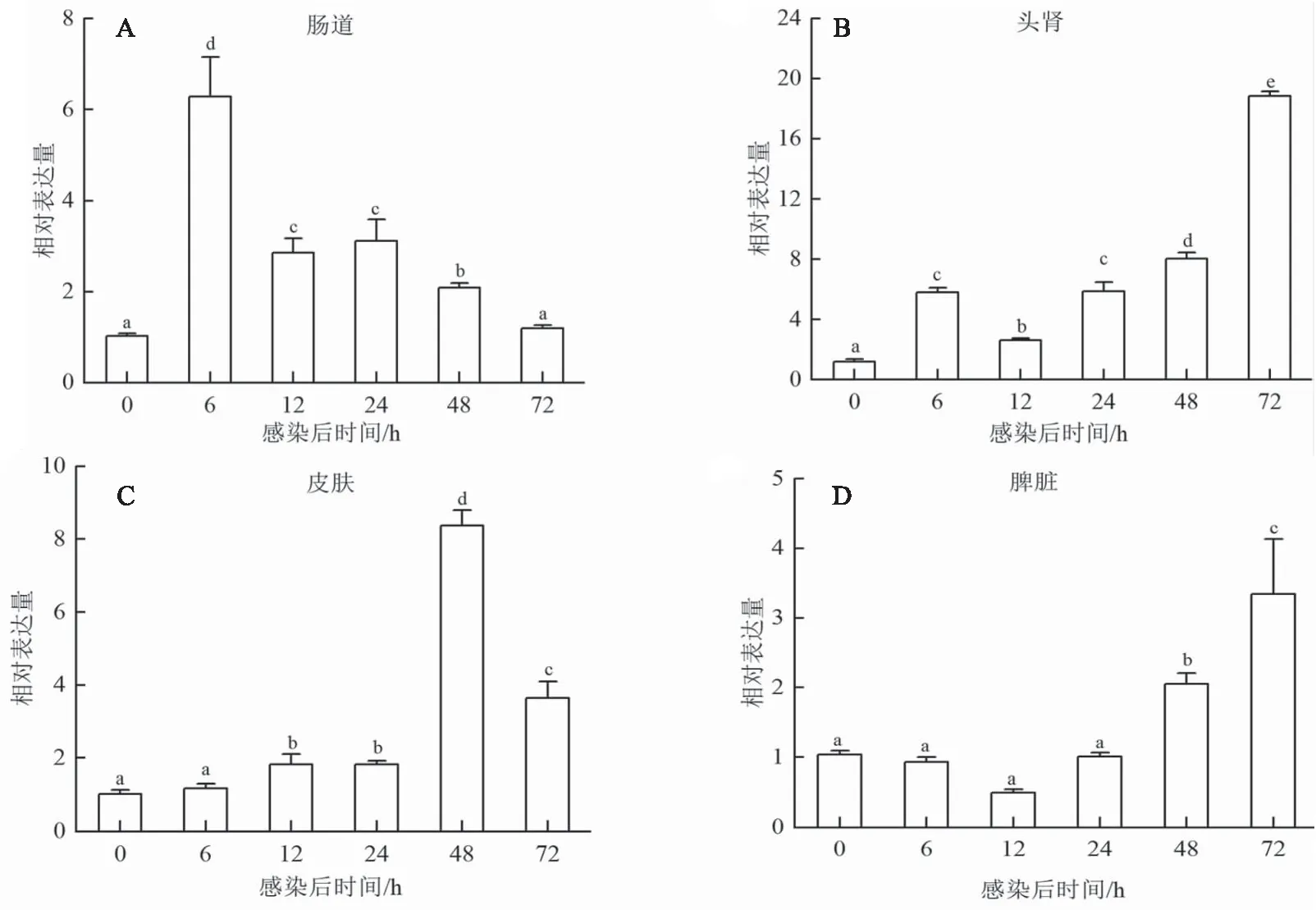
图6 嗜水气单胞菌感染后sacd22在七彩神仙鱼不同组织中(A-D)的表达分析
3 讨论
IgSF成员通过突变和选择进化出大量的蛋白质,这些蛋白质参与了细胞识别、粘附、发育和形态发生的过程,在特异性和非特异性免疫反应中发挥了关键作用[16-17]。本研究根据七彩神仙鱼皮肤转录组测序结果,克隆获得sajam-a和sacd22。其推导的氨基酸序列均包含1个信号肽,表明其蛋白质可能分泌到细胞外基质中,作为细胞间信使行使功能[16]。另外,两个基因均至少包含一个Ig结构域,这也是证明其为IgSF成员的重要依据。序列比对显示,sajam-a和sacd22的氨基酸序列均与同为慈鲷科的尼加拉瓜湖始丽鱼同源性最高,这一结果证明了克隆得到的sajam-a和sacd22基因序列的准确性。进化树分为鱼类和哺乳动物两大支,jam-a和cd22氨基酸序列进化关系与动物本身的进化分类吻合,揭示jam-a和cd22在进化过程中的保守性。
组织测定表明sajam-a和sacd22在各组织中都有不同的表达模式,并存在显著的组织差异性。其中,sajam-a在鳃中表达量最高,其次是肝脏、头肾、肠道和皮肤。鳃是重要的免疫组织,可以与水生环境中的各种病原体相互作用,其免疫功能和结构完整性是鱼类正常生长和抗病能力所必需的[18]。肠道和皮肤存在着淋巴组织,包含着淋巴细胞、巨噬细胞等,是鱼类重要的黏膜免疫组织,能抵御病原体的入侵[19]。头肾和脾脏是硬骨鱼中最大的淋巴组织和免疫活性组织,巨噬细胞较多,具有吞噬及捕获抗原的功能[20]。草鱼的Gcjam-a主要在鳃和肠道中表达,其次为肾和肝等免疫组织,结果与本研究相似[10]。在七彩神仙鱼中,sacd22也在10个组织中广泛表达,在脾脏和头肾等免疫组织中表达量较高。sajam-a和sacd22在七彩神仙鱼这些免疫相关组织中的高表达,表明其在机体免疫机制中发挥重要的作用。
嗜水气单胞菌感染鱼体后,sajam-a和sacd22的表达量在肠道、头肾、脾脏和皮肤中均呈现显著上调,但二者具有不同的表达模式。皮肤是机体免疫防御的第一道屏障,在皮肤中sajam-a迅速响应,表达量6 h开始显著上调,12 h达到峰值。同样,sajam-a的表达量在另外3个组织中6 h均显著上调,然后表达量逐渐下降,恢复到正常水平。而sacd22的表达量在皮肤中12 h开始上调,脾脏中48 h开始上调,并且发现sacd22在72 h内维持较高的表达量。JAMs在细胞间接触部位触发细胞内信号级联的能力调节多种生物过程,包括白细胞渗出、紧密连接形成和维持、细胞旁通透性、细胞极化、屏障功能调节、细胞迁移和血小板活化的调节[9]。在哺乳动物中,已经证实jam-a为轮状病毒(Rotaviruses)[21]、猫杯状病毒(Felinecalicivirus)[22]、呼肠孤病毒(Mammalianreovirus)[23]的受体。大量研究表明,病原刺激会影响jam-a基因的表达[24-25]。Siglecs是一类包含Ig结构域的细胞表面受体,主要存在于脊椎动物的各种免疫细胞中,可以与含唾液酸的聚糖特异性结合,促进细胞粘附和细胞信号传导[11]。CD22作为Siglecs家族的一员,可以调控B细胞受体信号传导阈值,对B细胞的早期发育发挥重要的作用,且在小鼠中验证了这一功能[26]。据报道,JAM-A可以通过细胞介导的防御反应促进先天免疫[10],而CD22通过调控B细胞信号传导,活化B细胞促进体液免疫[11],这可能是sajam-a和sacd22在七彩神仙鱼感染病原菌后组织中表达模式不同的原因。七彩神仙鱼免疫组织中sajam-a和sacd22的变化,表明了sajam-a和sacd22均参与了抗菌免疫反应,推测在七彩神仙鱼亲代抚育阶段,sajam-a和sacd22分别对亲鱼先天免疫和体液免疫反应发挥了重要的作用,具体作用机制还需要进一步的研究。
猜你喜欢
科普童话·学霸日记(2021年2期)2021-09-05 09:28:47
快乐语文(2021年11期)2021-07-20 07:41:18
快乐语文(2019年12期)2019-06-12 08:41:44
中学生百科·大语文(2019年1期)2019-04-17 04:45:14
文学少年(绘本版)(2018年9期)2018-11-20 10:42:20
小学生导刊(2018年22期)2018-08-21 01:47:48
快乐语文(2018年18期)2018-08-18 03:00:46
临床医药文献杂志(电子版)(2017年11期)2017-05-17 04:48:47
当代医药论丛(2017年22期)2017-04-12 06:29:43
小布老虎(2016年4期)2016-12-01 05:46:09
