
基于线粒体Cytb基因的拟裸蝗属部分种类的系统发育关系
2022-11-16 13:33熊忠平吴培福
保山学院学报 2022年5期
熊忠平 杨 晨 吴培福 柳 青
(1.西南林业大学 生物多样性保护学院,云南省森林灾害与控制重点实验室,昆明 650224;2.保山学院 资源环境学院,云南省高校滇西昆虫资源保护与利用重点实验室,云南 保山 678000)
利用现代分子生物学技术对昆虫遗传物质的研究和分析,探讨昆虫各类群间的系统发育、遗传进化、物种形成和分化等问题是当前昆虫分子系统学研究的热点[1]。线粒体基因(mitochondrial DNA,mtDNA)由于具有严格的母系遗传、缺少基因重组以及较快的进化速率等特点,而被广泛用于昆虫系统发育、种群遗传变异和分化以及近缘种、种下分类阶元分子鉴定等方面[2-4]。其中,线粒体Cytb(Cytochrome b,Cytb)基因是目前昆虫线粒体基因组13个编码蛋白基因中结构与功能研究得最清楚的基因之一[5-6]。Cytb基因进化速率适中,较短的一个片段就能包含种下、种级、属级乃至科级水平的系统信号,并且该基因中不同进化速率的密码子位点以及保守和突变的区域同时存在,使得该基因广泛应用于昆虫系统进化研究中[6-7]。有学者应用Cytb基因和16S rRNA基因序列的联合数据分析了北美田间蟋蟀Gryllus属11种13个种群的系统演化关系,指出欧洲的Gryllus属与北美洲的Gryllus属间具有明显的分枝,枝间分歧大于枝内分歧[8]。也有学者通过对我国蝗总科8科10种蝗虫的Cytb基因序列测定和构建分子系统树,发现槌角蝗科Gomphoceridae、网翅蝗科Arcypteridae和剑角蝗科Acrididae 3科关系相对较近,它们是较为进化的类群,这与传统的分类观点基本一致[9];利用Cytb基因对我国斑翅蝗科l4种的系统发育关系研究表明,14种蝗虫可以明显的分为4个支系[10];对黄胫小车蝗Oedaleus infernalisSaussure线粒体Cytb研究表明,其种群间具有较高的遗传多样性,可能由于其迁移能力较强,弱化了其种群间的遗传分化,使其种群间基因交流频繁,地理距离并未对其种群间的基因交流产生影响,也不是造成其种群间遗传分化的主导因素[11];基于线粒体Cytb基因对新疆荒漠半荒漠草原意大利蝗不同地理种群的遗传多样性分析表明,意大利蝗群体稳定,历史上未出现群体扩展,地理距离可能不是影响种群间遗传距离的重要因素[3]。
蝗虫是直翅目Othorptera蝗总科Acridoidea的统称,也是昆虫纲中的重要类群之一,许多种类是农、林、园艺作物的重要害虫[12]。目前,全世界已知直翅目有效种(亚种)为31 819种,我国直翅目物种记录有3 633种[13]。拟裸蝗属Conophymacris是蝗虫的代表性类群,隶属蝗总科斑腿蝗科Catantopidae,是Willemse于1933年建立[14],属模式种Conophymacris chinensis标本采自于云南昆明[15]。拟裸蝗属也是我国特有的短翅型蝗虫类群,目前已知有10种,分别为中华拟裸蝗C.chinensisWillems、楚雄拟裸蝗C.chuxiongensisWang、锥尾拟裸蝗C.conicercaBi et Xia、云南拟裸蝗C.yunnanensisCheng、香格里拉拟裸蝗C.xianggelilaensisNiu et Zheng、黑股拟裸蝗C.nigrofemuraLiang、苍山拟裸蝗C.cangshanensisZheng et Mao、绿拟裸蝗C.viridisZheng、九龙拟裸蝗C.jiulongensisZheng et Shi和四川拟裸蝗C.szechwanensisChang[15],均分布在我国云南省中部及北部到四川西南部的横断山东部地区,并且大部分种类呈岛屿状分布[16]。经作者前期的观察,拟裸蝗属部分种间的差异并不明显,部分种类的确定目前还存在着分歧,有必要进一步厘定其属内的种间关系。因此,本研究利用线粒体Cytb基因测序分析拟裸蝗属部分种类的遗传特征,构建该属昆虫的系统发育关系,为该属昆虫的进化及分类研究提供基础资料。
1 材料与方法
1.1 实验材料
本研究野外调查期间共采集到拟裸蝗属昆虫6种,具体标本采集信息见表1。蝗虫标本的鉴定主要以郑哲民《蝗虫分类学》[17]为依据。

表1 六种拟裸蝗野外采集信息
1.2 实验仪器和试剂
高速冷冻离心机(Eppendorf AG Germany),微型离心机(BIO-RAD韩国分公司),PCR仪(BIO-RAD),微量移液枪(Eppendorf),DYY-Ⅲ-6B型稳压稳流电泳仪(北京市六一仪器厂),凝胶成像仪(GEL Japan),恒温水浴振荡摇床(天津奥特赛斯仪器有限公司),全自动立式压力蒸汽灭菌锅(上海博讯实业有限公司)。
动物组织DNA提取试剂盒(Animal tissue DNA Kit,Omega),蛋白酶K(Omega)、上样缓冲液(双染料6×loading buffer)、BM2000 DNA Marker、琼脂糖(昆明硕阳科技有限公司)、50×TAE电泳缓冲母液(昆明硕阳科技有限公司)、核酸染料(昆明硕阳科技有限公司)、2×Power Taq PCR MasterMix(昆明硕阳科技有限公司)、酚:氯仿:异戊醇(25∶24∶1)(昆明硕阳科技有限公司)、氯仿:异戊醇(24∶1)(昆明硕阳科技有限公司)、SDS(天津市科密欧化学试剂开发中心)。
1.3 试验方法
1.3.1 蝗虫基因组DNA提取
蝗虫基因组DNA的提取按照试剂盒操作步骤进行。具体如下:
(1)用镊子取一只拟裸蝗的胸部肌肉及腿部肌肉组织,加入到1.5 mL高温灭菌的离心管中,用高温灭菌的研磨棒将样本研磨粉碎;
(2)用微量移液枪向离心管中再加入600 μL DNA提取缓冲液和10 μL蛋白酶K(20 mg/mL),充分混匀混合,使蛋白酶K与组织细胞充分接触;
(3)将离心管放置在预先加热到65℃的恒温水浴振荡摇床上,震荡消化3 h,每隔0.5 h人工震荡混匀一次;
(4)将水浴消化的离心管上清液转入到另一离心管中,加入600 μL已制备的酚:氯仿:异戊醇(25∶24∶1)充分混匀;
(5)将离心管置于高速冷冻离心机12 000 rpm/min离心10 min;
(6)将离心管中的上清液转移至另一支1.5 mL离心管中,加入600 μL氯仿:异戊醇(24∶1)充分混匀;
(7)将离心管置于高速冷冻离心机12 000 rpm/min离心10 min;
(8)再将离心管中的上清液转移至另一支1.5 mL离心管中,加入无水乙醇1 200 μL,轻柔混匀后置入冰箱(-20℃)中10 min;
(9)将离心管置于高速冷冻离心机12 500 rpm/min离心15 min,弃上清液(注意不要把DNA倒掉);
(10)在离心管中加入220 μL Buffer BL,充分混匀,在70℃水浴锅中孵化10 min;
(11)加入220 μL无水乙醇,在漩涡混合器中充分震荡混匀后于微型离心机中离心;
(12)将试剂盒中的圆柱套在收集管上,把离心管中的上清液转移到圆柱中,将收集管置于高速冷冻离心机8 000 rpm/min离心1 min,后倒掉收集管中液体;
(13)加入500 μL Buffer HB于圆柱中,将收集管置于高速冷冻离心机8 000 rpm/min离心1 min,后倒掉收集管中液体;
(14)加入650 μL DNA Wash Buffer于圆柱中,将收集管置于高速冷冻离心机8 000 rpm/min离心1 min,后倒掉收集管中液体;
(15)加入650 μL DNA Wash Buffer于圆柱中,将收集管置于高速冷冻离心机8 000 rpm/min离心1 min,后倒掉收集管中液体;
(16)将收集管置于高速冷冻离心机12 000 rpm/min离心3 min,后倒掉收集管中液体;
(17)圆柱下换一个新的收集管,加入水浴锅预热70℃的Elution Buffer 50 μL,室温放置3 min;
(18)将收集管置于高速冷冻离心机12 500 rpm/min离心3 min,收集管中即为所提DNA。
1.3.2 引物设计与PCR扩增
PCR引物根据NCBI GenBank中飞蝗及其他鞘翅目昆虫的线粒体核苷酸数据,自行设计扩增拟裸蝗属线粒体Cytb基因的引物(见表2),预计扩增大小约为690 bp。引物由上海生工生物工程有限公司合成。

表2 拟裸蝗属6种蝗虫Cytb基因PCR扩增引物序列
PCR总反应体系为25 μL,包括PCR mix 12.5 μL,上游引物CBJ 0.5 μL,下游引物CBN 0.5 μL,模板5 μL,灭菌水6.5 μL。扩增程序为:94℃预变性4 min;94 ℃变性40 s,44℃退火35 s,72℃延伸50 s,30个循环;循环结束后于72℃下总延伸6 min。PCR产物用1%的琼脂糖凝胶电泳检测,以BM2000 DNA Marker作为参照标准,在120 V稳定电压条件下电泳30 min,凝胶成像并记录结果。经检测合格的PCR产物委托上海生工生物工程有限公司进行样品的纯化和双向测序。
1.3.3 数据统计和分析
采用DnaMan 6.0软件对所测序列进行比对分析,采用DNAStar V7.1.0软件MegAlign程序中的Clustal W方法来比对核苷酸和推导氨基酸序列,分析氨基酸组成。应用MEGA(Version 6.0)软件分析拟裸蝗属昆虫的碱基组成百分比、密码子第三位G+C百分含量(GC3S),种间和种内进化距离,基因突变情况以及构建系统发育树。利用DNAsp 5.0和MEGA(Version 6.0)程序来分析密码子使用情况。
2 结果与分析
2.1 拟裸蝗属昆虫线粒体Cytb基因的碱基组成
拟裸蝗属6种蝗虫线粒体Cytb基因序列中A、T、G、C碱基的平均含量分别为34.04%、39.32%、10.67%、15.97%;A+T平均含量(73.36%)明显高于G+C平均含量(26.64%)。6种蝗虫的碱基组成百分比基本一致,但亦存在差异;绿拟裸蝗和九龙拟裸蝗Cytb基因的T碱基平均含量高于其他各种,苍山拟裸蝗的C碱基平均含量高于其他各种(见表3)。

表3 拟裸蝗属6种蝗虫Cytb基因的碱基组成特征
拟裸蝗属6种蝗虫的AT斜率和GC斜率均为负值,进一步表明各种类碱基组成中A、G碱基含量低于T、C碱基含量;结合6种蝗虫的AT斜率和GC斜率的绝对值,可以看出各种类中A碱基含量略低于T碱基含量,而G碱基含量明显低于C碱基含量。此外,6种蝗虫的GC3S平均含量存在一定的差异,其中香格里拉拟裸蝗GC3S平均含量(13.33%)最大,九龙拟裸蝗GC3S平均含量(10.83%)最小(见表3)。
2.2 拟裸蝗属昆虫线粒体Cytb基因的碱基替换
拟裸蝗属6种蝗虫的线粒体Cytb基因的碱基替换主要是发生碱基的颠换和转换,且碱基替换大都发生在密码子的第三位点。拟裸蝗属6种蝗虫整体的碱基颠换率为1.42,转换率为22.15,表明转换率均大于颠换率(见表4)。

表4 拟裸蝗属6种蝗虫Cytb基因的碱基替换
2.3 拟裸蝗属昆虫线粒体Cytb基因的种间进化距离
拟裸蝗属6种蝗虫中,以云南拟裸蝗和绿拟裸蝗的种间进化距离最大(0.068±0.014),九龙拟裸蝗和中华拟裸蝗的种间进化距离最小(0.017±0.007)。整体而言,绿拟裸蝗与其他拟裸蝗属种间的进化距离均较大,但与香格里拉拟裸蝗种间进化距离较小(见表5)。

表5 拟裸蝗属6种蝗虫Cytb基因的种间进化距离
2.4 拟裸蝗属昆虫线粒体Cytb基因系统发育关系
以太白秦岭蝗Qinlingacris taibaiensisYin et Chou和日本条螽Ducetia japonica(Thunberg)为外群,基于最大似然法(ML)和最大简约法(MP)构建拟裸蝗属6种蝗虫的系统发育树,结果表明,最大似然法和最大简约法两种方法建树结果基本一致,6种蝗虫大体可以分为两个分支,其中绿拟裸蝗、香格里拉拟裸蝗和苍山拟裸蝗构成一个大分支,中华拟裸蝗、九龙拟裸蝗和云南拟裸蝗3种构成另外一个分支;绿拟裸蝗和香格里拉拟裸蝗聚在一个小分支上,说明两者之间关系相对较近,与苍山拟裸蝗的关系较远;云南拟裸蝗和九龙拟裸蝗聚在一个小分支上,同样说明两者具有较近的亲缘关系,而与中华拟裸蝗关系较远(见图1)。
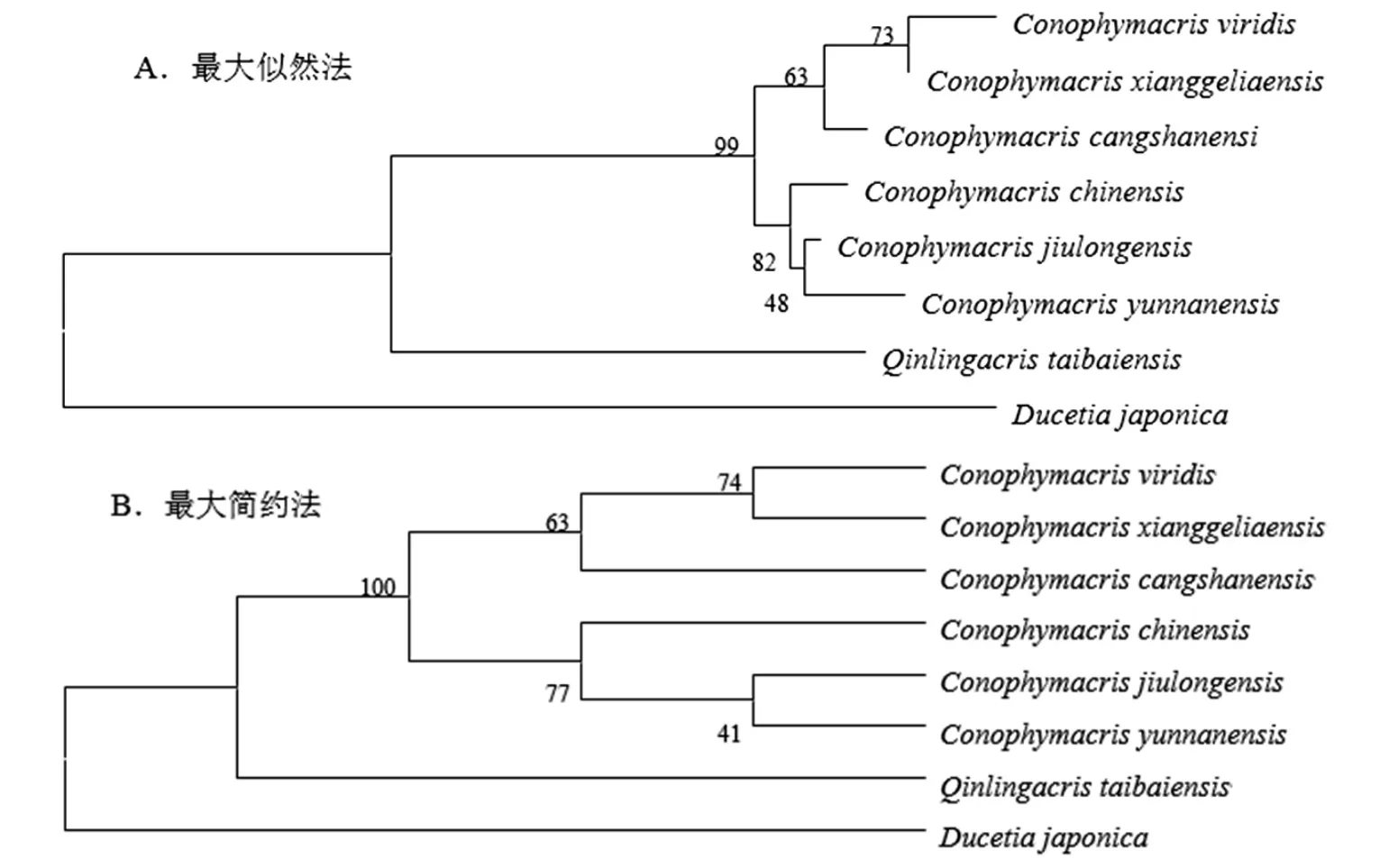
图1 拟裸蝗属6种蝗虫的系统发育关系
3 结论与讨论
自1989年Kocher等设计了第一对脊椎动物Cytb基因部分片段的扩增引物后,该基因立即被广泛应用于动物系统学研究中,目前被认为是对动物种上和种下阶元进行系统进化研究最好的分子标记之一[6,18]。本研究采用Cytb基因对拟裸蝗属6种蝗虫的遗传特征和系统发育关系进行了研究,结果表明,拟裸蝗属6种蝗虫的线粒体Cytb基因的碱基组成百分比基本一致,序列中A+T平均含量为73.36%,明显高于G+C平均含量26.64%,表现出强烈的AT偏向性,这一结果与前人对其他许多昆虫的线粒体Cytb基因研究结果类似,普遍认为在昆虫的Cytb基因中,A+T含量为明显高于G+C(A+T达到80%,G+C仅为20%),尤其是在密码子第三位点,这种偏向更加明显(A+T达到95%,G+C仅为5%)[19];此结果也与前人等对我国蝗总科昆虫的Cytb基因测序研究结果一致[8,20],但与绢蝶(75.4%)[21]、果蝇(78.6%)[22]和意大利蜜蜂(84.9%)[23]相比则偏低。
拟裸蝗属6种蝗虫线粒体Cytb基因的碱基替换主要是发生颠换和转换,且转换率均大于颠换率,这与其他直翅目昆虫相一致[9-10]。有研究认为,与其他昆虫相比,蝗总科昆虫每个氨基酸密码子第三位的A+T含量较高[9],而较高的A+T含量增加了AT颠换的可能性,导致了氨基酸密码子第3位点颠换速率的增加,进而加速氨基酸的变异[24-25]。本研究发现的拟裸蝗属6种蝗虫Cytb基因碱基替换与A+T含量及氨基酸变异之间的关系还有待进一步研究。
本研究发现,6种蝗虫间以云南拟裸蝗和绿拟裸蝗的种间进化距离最大,九龙拟裸蝗和中华拟裸蝗的种间进化距离最小。这与6种蝗虫的野外分布格局较为吻合(表1),中华拟裸蝗是6种蝗虫中分布最为广泛的种类,在云南昆明、丽江、中甸及四川泸定等地均有分布,云南拟裸蝗主要分布在云南个旧和昭通等地区,而绿拟裸蝗和九龙拟裸蝗分布范围则十分狭窄[16],加之拟裸蝗属是典型的短翅型种类,无飞翔能力,其活动和扩散区域十分有限,因此推测地理隔离可能是导致拟裸蝗属6种蝗虫种间进化的主要因素。
拟裸蝗属目前记载种类较少,但少数物种包括种下阶元的归隶仍具有一定的争议,因此,及时厘清其属内种间系统关系就显得十分必要。本研究表明,采用最大似然法和最大简约法构建的拟裸蝗属6种蝗虫的系统发育树基本一致,6种蝗虫大体可以分为两个分支,其中绿拟裸蝗、香格里拉拟裸蝗和苍山拟裸蝗构成一个大分支,中华拟裸蝗、九龙拟裸蝗和云南拟裸蝗3种构成另外一个分支。易传辉等采用支序生物地理学的方法研究了拟裸蝗属8种蝗虫的系统发育关系,认为拟裸蝗属不是一个严格的单系群,8种蝗虫中四川拟裸蝗单独聚为一支,与其他拟裸蝗的亲缘关系较远;其余的7种拟裸蝗又分为两个大的分支,其中云南拟裸蝗、中华拟裸蝗和绿拟裸蝗具有较近的亲缘关系而聚为一支,锥尾拟裸蝗、苍山拟裸蝗、楚雄拟裸蝗和黑股拟裸蝗4种具有较近的亲缘关系而聚为一支[16]。这与本研究结果并不一致,本研究中绿拟裸蝗与苍山拟裸蝗因具有较近的亲缘关系而聚为一个大的分支。当然,本研究涉及种类和方法与易传辉等人的研究不尽相同,有待今后更全面的标本采集,对该属所有种类进行线粒体基因组测序并结合外部形态特征进一步确证该属种间的系统发育关系。
猜你喜欢
小哥白尼(野生动物)(2022年5期)2022-08-15
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
科技视界(2020年26期)2020-09-24
科技视界(2020年17期)2020-07-30
科学大众(2020年10期)2020-07-24
故事作文·低年级(2019年6期)2019-06-20
蔬菜(2018年5期)2018-05-17
化学教学(2018年1期)2018-02-28
幼儿教育·父母孩子版(2017年10期)2017-12-13
杂草学报(2015年2期)2016-01-04
