
都匀毛尖本地种茶树响应假眼小绿叶蝉侵害的代谢组学研究
2022-11-16 06:10王芬木仁李国栋喻佩李静崔宝禄吕立堂
湖南农业大学学报(自然科学版) 2022年5期
王芬,木仁,李国栋,喻佩,李静,崔宝禄,吕立堂*
都匀毛尖本地种茶树响应假眼小绿叶蝉侵害的代谢组学研究
王芬1,木仁1,李国栋2,喻佩1,李静1,崔宝禄1,吕立堂2*
(1.黔南民族师范学院生物科学与农学院,贵州 都匀 558000;2.贵州大学茶学院,贵州 贵阳 550025)
以被假眼小绿叶蝉侵染0、12、24 h的都匀毛尖本地种茶树叶片为试验材料,采用超高效液相色谱–串联四级杆飞行时间质谱技术对试验材料的代谢产物进行定性和定量分析,正离子模式下鉴定出1094个代谢物。采用主成分分析、正交偏最小二乘法判别分析,根据差异倍数筛选出0 h与12 h处理有20个差异代谢物,其中6个差异代谢物的含量上调,14个下调;0 h与24 h处理之间有41个差异代谢物,其中26个上调,15个下调;12 h与24 h处理有59个差异代谢物,其中45个上调,14个下调。0 h与12 h处理有2个差异代谢物注释到KEGG数据库中,其中芥子酸注释到苯丙素生物合成途径中,与木质素形成有关;0 h与24 h处理有6个差异代谢物注释到KEGG数据库中,其中6,9,12,15十八碳四烯酸乙酯注释到亚麻酸代谢通路,没食子儿茶素注释到类黄酮生物合成,苯乙醛注释到苯丙氨酸代谢通路,推测苯乙醛会对周围的假眼小绿叶蝉造成其行为或生理的影响,而苯丙氨酸经苯丙氨酸解氨酶等酶催化合成木质素和类黄酮等;12 h与24 h处理有9个差异代谢物注释到9条通路中,马钱苷酸注释到单萜生物合成通路中,相比侵染12 h处理,24 h处理的马钱苷酸含量上调,表明茶树被假眼小绿叶蝉为害后可能通过释放萜类物质来抵御害虫。以上结果表明,木质素、萜类物质和黄酮类化合物在茶树抵御小绿叶蝉的侵害具有重要作用。
茶叶;假眼小绿叶蝉;代谢组学;超高效液相色谱–串联四级杆飞行时间质谱;差异代谢物
茶树的叶片和叶梢是茶叶加工的原料,但茶树的叶片和嫩梢常易被小绿叶蝉取食,进而影响茶叶品质[1]。在茶树生长过程中,假眼小绿叶蝉对茶树的侵害会引起一系列生理生化的防御反应[2–3],且会激活一系列激素调节的信号通路(如激活水杨酸、茉莉酸、脱落酸、乙烯等的信号通路)[4–6]来进行害虫侵染的防御反应。在小绿叶蝉侵染茶树的过程中其代谢产物会受到严重影响,导致茶树鲜叶的氨基酸、多酚类、生物碱、萜类等含量发生变化[7]。CHO等[8]研究表明,小绿叶蝉侵染后的茶叶可以制成具有独特香气的东方美人茶。LIAO等[9]研究发现小绿叶蝉侵害增加了茶叶中茉莉酸和水杨酸的含量。MEI等[10]发现通过小绿叶蝉侵染茶叶可诱导茶叶中芳樟醇的释放。MU等[11]的研究结果表明,小绿叶蝉侵染的茶树可诱导绿叶释放挥发物,增强对害虫的抗性。ZENG等[4]研究发现小绿叶蝉取食不但可以提高香叶醇和芳樟醇的释放,还可增强萜烯合酶基因的表达。可见,研究茶树对假眼小绿叶蝉取食的应激防御机制具有重要意义。
代谢组学是研究生物体代谢途径的一门学科。采用代谢组学技术对生物体受外界的刺激时代谢产物的变化情况进行定性与定量分析[12],可发现生命活动的动态变化规律。已有研究人员应用代谢组学技术对茶叶加工过程中儿茶素等代谢产物的变化、不同茶叶的化学成分以及茶叶的抗逆胁迫等进行了研究[13–15]。王春波等[16]通过超高效液相色谱–串联四级杆飞行时间质谱(UPLC–Q–TOF/MS)研究发现,NW34品系比NW48品系的表没食子儿茶素没食子酸酯、表儿茶素等代谢物的含量高。本研究以假眼小绿叶蝉吸食0、12、24 h的都匀毛尖本地种茶树叶片为试验材料,运用UPLC–Q–TOF/MS代谢组学技术,研究茶鲜叶被假眼小绿叶蝉取食后代谢产物的变化,旨在为都匀毛尖本地种茶树抗虫良种的选育提供理论参考。
1 材料与方法
1.1 植株材料
2018年11月,从贵州省都匀市红敏茶园选取1株枝叶茂盛、长势良好的茶树作为母株,剪取健康的茎或叶在生根粉水溶液中浸泡2 h,然后移入花盆中进行扦插,在温室大棚中进行精心培养。2021年6月,选取9株长势良好并且株高基本一致的茶苗放入纱网笼中进行假眼小绿叶蝉侵染,每3株放入1个笼中,分为3组:第1组侵染0 h;第2组侵染12 h;第3组侵染24 h。分别在每组3株茶苗上剪取3~5片健康嫩叶作为侵染0、12、24 h的3个生物学重复。共9个样本。固氮处理后放入–80 ℃的冰箱中保存。
1.2 主要试剂与仪器
Merck甲醇、Merck乙腈、Merck甲酸均为色谱纯。主要仪器有SHIMADZU出产的Nexera X2型超高效液相色谱仪;Scientz出产的Scientz–100F型冻干机;Retsch出产的MM400型研磨仪;ABI出产的Applied Biosystems QTRAP series型串联质谱;Agilent出产的Agilent SB–C18型色谱柱。
1.3 方法
1.3.1样本制备
将茶叶样品放置于冻干机中真空冷冻干燥后研磨至粉末状,称取100 mg粉末,溶解于1.2 mL 70%甲醇提取液中,每30 min涡旋1次,每次持续30 s。共涡旋6次。样本放于4 ℃冰箱过夜,12 000 r/min离心10 min,吸取上清液,用微孔滤膜过滤样品,并保存于进样瓶中,以用于UPLC–MS/MS分析[17–18]。
1.3.2液相条件
采用Agilent SB–C18色谱柱(1.8 μm、2.1 mm×100 mm)。流动相:A相为超纯水(加入0.1%的甲酸),B相为乙腈(加入0.1%的甲酸)。洗脱梯度:0 min(5%B),9 min(95%B),10~11 min(5%B),11~14 min(5%B)。流速为0.35 mL/min,柱温为40 ℃,进样量为4 μL。
1.3.3质谱条件
AB6500三重四级杆线性离子阱质谱仪UPLC/MS/MS系统由Analyst1.6.3软件(AB Sciex)控制运行正负2种离子模式。ESI源操作参数设置为:离子源,涡轮喷雾;源温度550 ℃;离子喷雾电压5500 V(正离子模式)/–4500 V(负离子模式);离子源气体I(GSI)、气体II(GSII)和帘气(CUR)分别设置为50、60和25 Psi。在三重四级杆(QQQ)和线性离子阱(LIT)模式下分别用10、100 μmol/L聚丙二醇溶液进行仪器调谐和质量校准。
1.3.4数据处理
基于标准品上机建库的信息和二级谱信息对9个样本的代谢物进行定性,并利用三重四级杆质谱的多反应监测模式进行定量分析。用MultiaQuant软件打开样本下机质谱文件,进行色谱峰的积分和校正工作,计算峰面积,然后进行数据归一化,对代谢物进行定量,随后对9个样本进行主成分分析,并利用R包ropls进行计算[19]。不同处理代谢产物的差异用检验和OPLS–DA模型的VIP值进行分析,最后根据代谢物名称在对应数据库中进行查找,对代谢物进行KEGG、HMDB和LIPID MAPS注释。
2 结果与分析
2.1 重复相关性评估
对假眼小绿叶蝉感染0、12、24 h的茶树叶片共9个样本进行重复相关性评估。将斯皮尔曼相关系数()作为生物学重复相关性的评估指标,2越接近1,说明每个时间点内重复样品的相关性越强。结果(图1)表明,除了12 h第1个重复和第3个重复之间的相关性较低外,其他2个时间点内的相关性均接近0.8或超过0.8。而一般认为样本间的相关性大于0.75时,数据质量较为可靠。可见,本研究中的差异代谢物的数据整体上比较可靠。
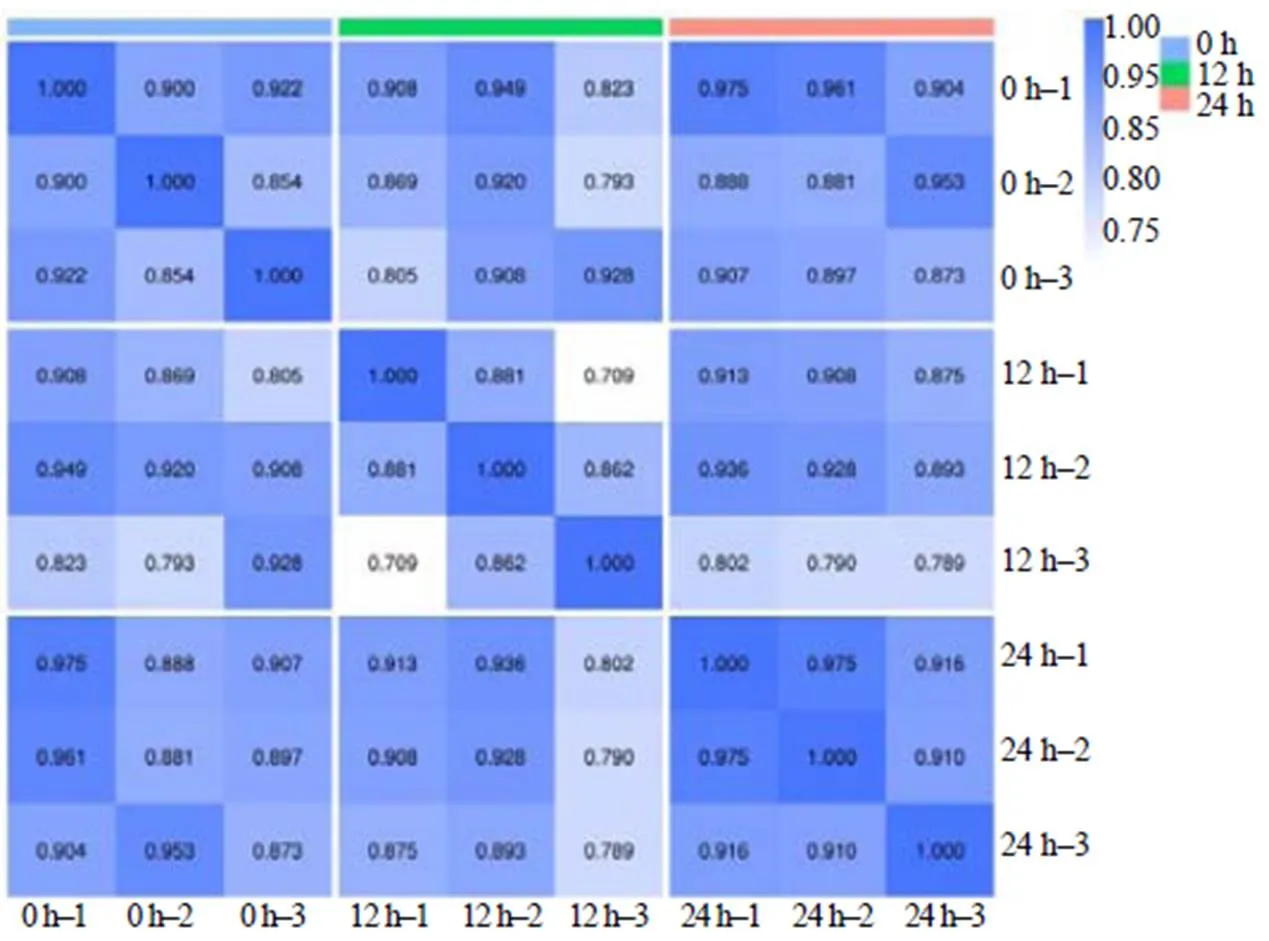
图1 9个样本间的相关性评估结果
2.2 主成分分析
采用UPLC–QTOF–MS对被假眼小绿叶蝉侵染0、12、24 h后的9个茶树叶片样品进行主成分分析(PCA)。从图2可以看出,除12 h的第1个重复与另外2个重复组内代谢物差异较大外,其他重复组内代谢物差异均较小,表明在试验过程中数据的重现性较好。在0、12、24 h的主成分分析中,主成分1和2贡献值分别为53.15%和22.05%,样品累积贡献值为75.20%,说明3组样品在代谢产物的种类、含量等方面存在差异,并且在其余的主成分中还存在一定的贡献值。在0 h和12 h的主成分分析中,主成分1和2的贡献值分别为62.60%和18.74%,累积贡献值为81.34%,说明代表性较好。在0 h和24 h的主成分分析中,主成分1和2的贡献值分别为49.27%和31.45%,累积贡献值为80.72%,说明这2个主成分解释了80.72%的原始变量信息。在12 h和24 h的主成分分析中,主成分1和2的贡献值分别是60.41%和24.26%,累积贡献值超过84%,说明主成分分析可为鉴别侵染12 h和24 h差异代谢物提供足够的可信度。
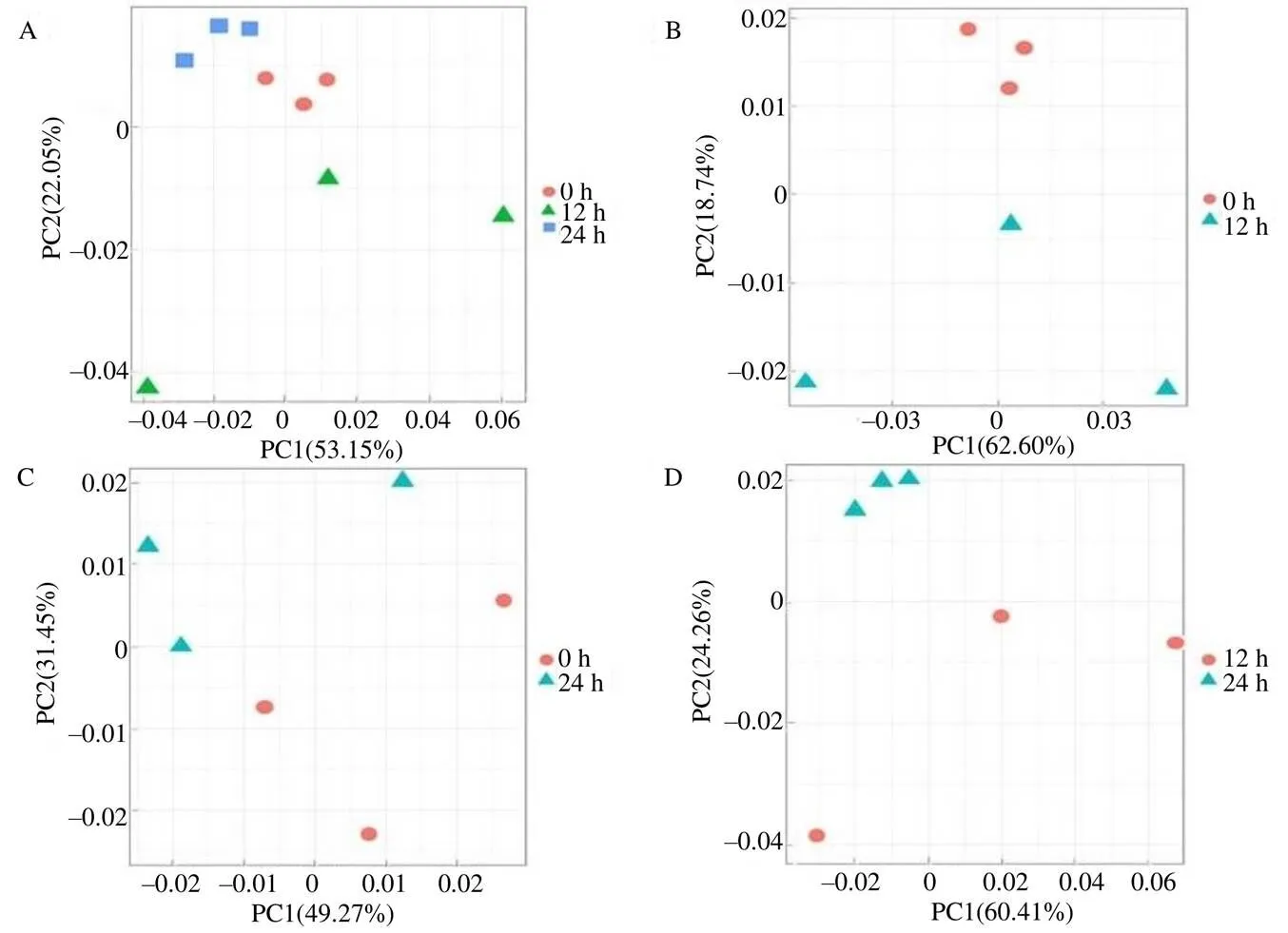
A 0、12、24 h处理代谢物主成分分析结果;B 0 h和12 h处理主成分分析结果;C 0 h和24 h处理主成分分析结果;D 12 h和24 h处理主成分分析结果。
2.3 正交偏最小二乘法判别分析
PCA分析对相关性较小的变量不敏感,而正交偏最小二乘法判别分析(OPLS–DA)可滤除与分类信息无关的噪音,从而更好地凸显组间的差异。OPLS–DA 分析(图3)中,0 h处理的3个重复分布于横坐标的负轴,12 h的3个重复分布于横坐标的正轴,区分效果明显,说明假眼小绿叶蝉侵染都匀毛尖本地种茶树叶片0 h和12 h在代谢成分上存在明显的差异。模型验证后参数值2=0.753,2=0.998,2=0.416,虽然2偏小,但能较好地对0 h和12 h处理的代谢物进行区分(图3–A)。在0 h和24 h处理的OPLS–DA模型分析(图3–B)中,2=0.659,2=0.996,2=0.604,说明模型符合样本数据真实情况。在12 h与24 h处理的OPLS–DA模型分析(图3–C)中,2组样本的重复点相距较近,表明数据重复性好,并且12 h和24 h处理的结果分布于两侧,差异显著。模型验证后参数值2=0.753,2=0.999,2=0.746,说明本研究中所建立的OPLS–DA模型能够有效揭示12 h和24 h处理的代谢物的差异。
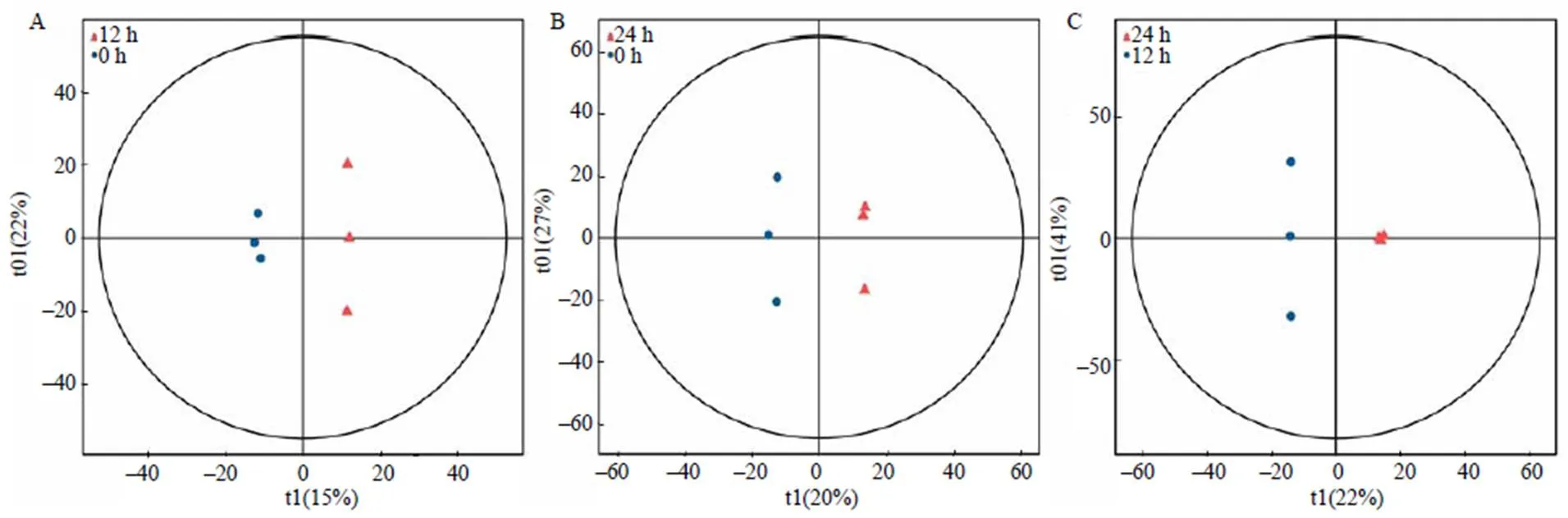
A 0 h与12 h正交偏最小二乘法判别分析结果;B 0 h与24 h正交偏最小二乘法判别分析结果;C 12 h与24 h正交偏最小二乘法判别分析结果。
2.4 差异代谢物筛选
对于有生物学重复的样本,按照VIP≥1和≤0.05来筛选茶树被假眼小绿叶蝉侵染0 h和12 h、0 h和24 h,12 h和24 h后的差异代谢物。正离子模式下共鉴定出1094个差异代谢物。0 h与12 h处理之间有20个差异代谢物,其中6个差异代谢物(环苜蓿烯、环氧石竹烯、二乙茎甲苯酰胺、丁香酸乙酯、甲基多巴酸盐、d–葡萄糖醛酸乙酯)的含量上调,14个差异代谢物(3–羟基2456–四甲氧基查尔酮、6–羟基高良姜素、3–甲硫基–1–丙醇、异胸腺素、O–甲基灵芝酸、7,4,5–三羟基–5,2–氧化–4–苯基香豆素、单酰甘油、丙氨酸丝氨酸络氨酸、前列腺素E2甲酯、红紫素、壬苯醇醚–9、芥子酸、6–O乙酰基大豆异黄酮、麦芽三糖)的含量下调。0 h与24 h处理有41个差异代谢物,其中26个差异代谢物(没食子儿茶素、3–羟基2456–四甲氧基查尔酮、苯乙醛等)的含量上调,15个差异代谢物(6,9,12,15十八碳四烯酸乙酯、四氢皮质酮、单酰甘油等)下调。12 h与24 h处理有59个差异代谢物,其中45个差异代谢物(没食子儿茶素、表没食子儿茶素、马钱苷酸等)的含量上调,14个(己二酸二环己酯、单酰甘油、环氧石竹烯等)下调(图4)。这些差异代谢物可能与响应假眼小绿叶蝉的侵染有关。对检测到的代谢物进行以2为底的对数处理,在正离子模式下列出了0 h与12 h、0 h与24 h、12 h和24 h处理之间变化最明显的5种差异代谢物(表1)。
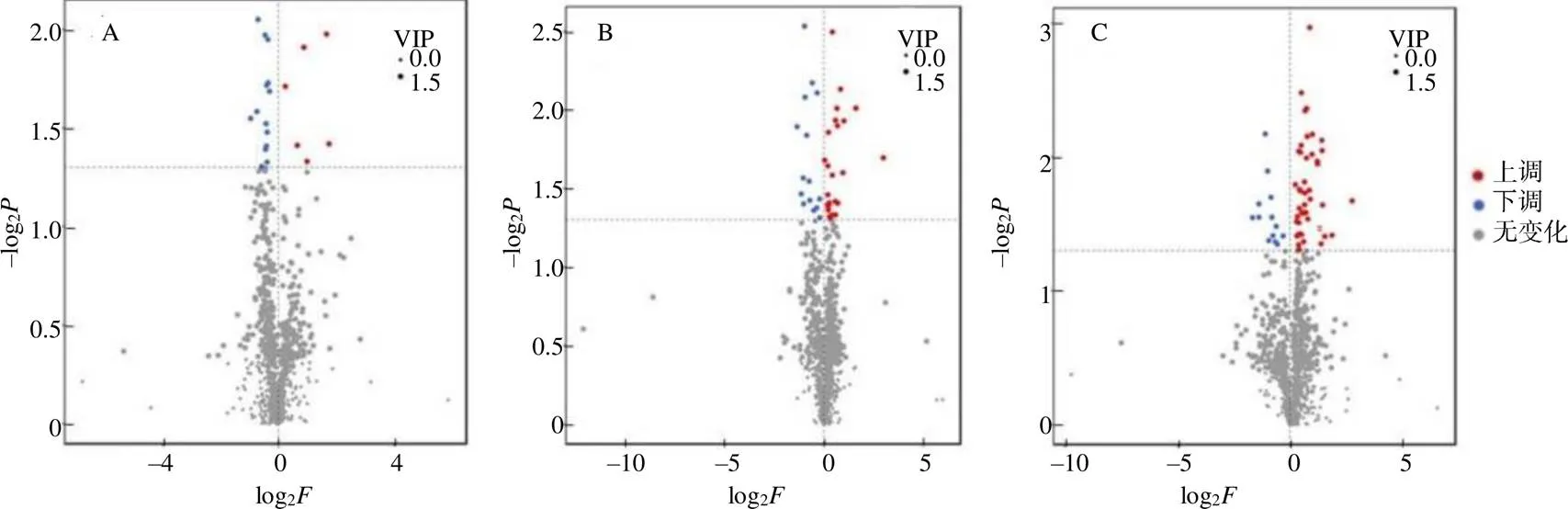
A 0 h与12 h差异代谢物火山图;B 0 h与24 h差异代谢物火山图;C 12 h与24 h差异代谢物火山图;红点代表上调差异代谢物,绿点代表下调,灰点代表差异不显著。P代表P值,F代表差异倍数。

表1 3个时间点间变化最明显的5种差异代谢物
2.5 差异代谢物注释
根据代谢物名称在数据库中进行查找,对代谢物进行注释,有215个代谢物注释到HMDB数据库中,278个代谢物注释到LIPID MAPS数据库中,151个代谢物注释到KEGG数据库中。将代谢物在KEGG COMPOUND数据库的注释信息匹配到对应的通路,发现0 h与12 h处理有2个差异代谢物注释到KEGG数据库中,芥子酸(pos_754)注释到苯丙素生物合成和次级代谢生物合成,麦芽三糖(pos_1052)注释到ABC转运蛋白通路中。0 h与24 h处理有6个差异代谢物注释到KEGG数据库中,其中6,9,12,15十八碳四烯酸乙酯(pos_153)注释到亚麻酸代谢通路;没食子儿茶素(pos_65)注释到类黄酮生物合成通路, 苯乙醛(pos_237)注释到苯丙氨酸代谢通路。12 h与24 h处理有9个差异代谢物注释到9条通路中,其中磷脂酰丝氨酸(pos_1090)(上调)注释到甘氨酸、丝氨酸和苏氨酸代谢通路中;没食子儿茶素(pos_65)、表没食子儿茶素(pos_157)和白杨素(pos_756)注释到类黄酮生物合成途径;马钱苷酸(pos_906)注释到单萜生物合成和次级代谢生物合成通路;十八碳炔烯醇酸(pos_258)和己二酸二环己酯(pos_405)注释到亚麻酸代谢。Cluster Profiler[20]是一个用于KEGG富集分析的R包,利用Cluster Profiler,选用超几何检验的方法对差异代谢物KEGG的注释结果进行富集分析,并利用Cytoscape[21]软件绘制富集网络图(图5)。芥子酸富集到苯丙素生物合成;没食子儿茶素、表没食子儿茶素、白杨素富集到类黄酮生物合成;6,9,12,15十八碳四烯酸乙酯、己二酸二环己酯、十八碳炔烯醇酸富集到亚麻酸代谢;马钱苷酸富集到单萜生物合成。
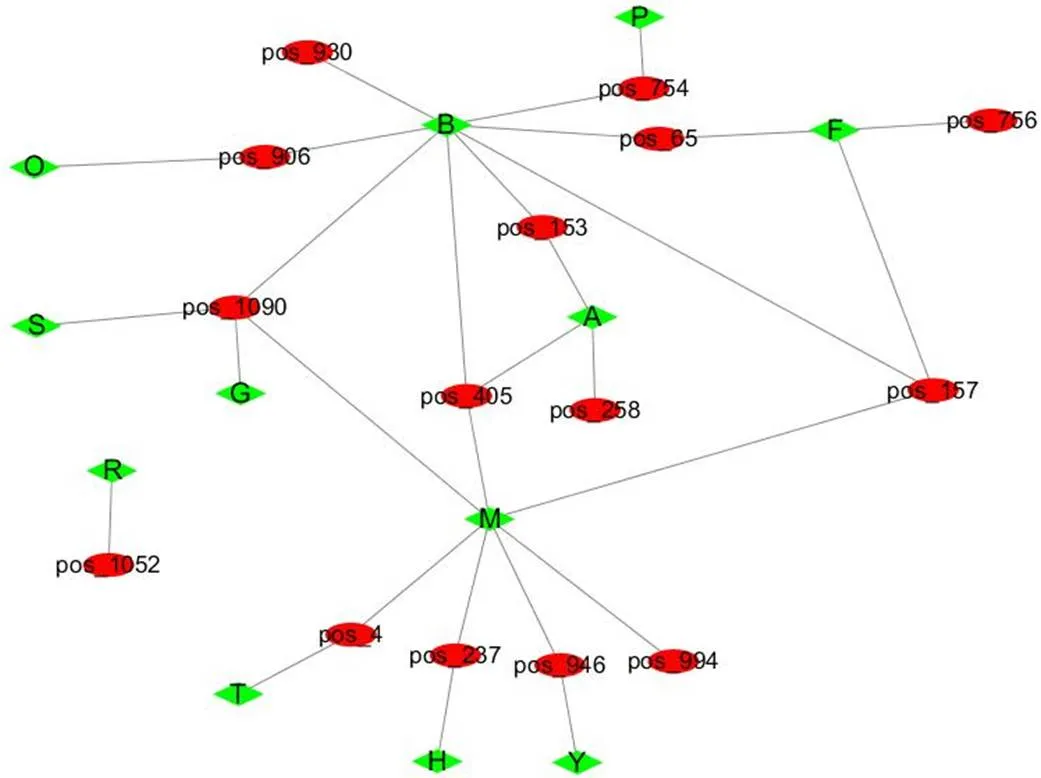
红色的椭圆形代表差异代谢物,绿色的菱形代表通路。pos_754为芥子酸;pos_65为没食子儿茶素;pos_930为道诺霉素;pos_906为马钱苷酸;pos_153为6,9,12,15十八碳四烯酸乙酯;pos_405为己二酸二环己酯;pos_1090为磷脂酰丝氨酸;pos_258为十八碳炔烯醇酸;pos_756为白杨素;pos_4为癸酸;pos_946为假尿苷;pos_994为去氢表雄酮;pos_1052为麦芽三糖;pos_157为表没食子儿茶素;pos_237为苯乙醛。B为次级代谢生物合成;P为苯丙素生物合成;F为类黄酮生物合成;A为亚麻酸代谢;M为代谢通路;G为甘油磷脂代谢;O为单萜生物合成;S为甘氨酸、丝氨酸、苏氨酸代谢;H为苯丙氨酸代谢;Y为嘧啶代谢;T为脂肪酸代谢;R为ABC转运蛋白。
3 结论与讨论
本研究利用UPLC–Q–TOF/MS代谢组技术对被假眼小绿叶蝉侵染0、12、24 h的都匀毛尖本地种茶树9个样品1094个代谢物进行了分析,结果显示:0 h与12 h处理有20个差异代谢物,6个上调,14个下调;0 h与24 h处理之间有41个差异代谢物,26个上调,15个下调;12 h与24 h处理之间有59个差异代谢物,45个上调,14个下调。通过主成分分析、正交偏最小二乘法判别分析和重复相关性评估证明获得的差异代谢物比较可靠。
正离子模式下假眼小绿叶蝉侵染0 h与12 h都匀毛尖本地种茶树有2个差异代谢物下调注释到KEGG数据库中,芥子酸(pos_754)注释到苯丙素生物合成途径中,麦芽三糖(pos_1052)注释到ABC转运蛋白通路中。芥子酸通过特异途径生成木质素单体,最终聚合形成木质素,在抵御植物抗虫等非生物胁迫中起重要作用[22]。
0 h与24 h处理有6个差异代谢物注释到KEGG数据库中,6,9,12,15十八碳四烯酸乙酯(下调)注释到亚麻酸代谢途径。茉莉酸由亚麻酸通过各种酶合成,当植物遭受害虫取食危害时,亚油酸和亚麻酸途径被激活,植物迅速合成大量的茉莉酸并激活下游信号转导,进而诱导大量抗虫次级代谢产物的合成,以此来抵御昆虫的取食。没食子儿茶素(上调)注释到类黄酮生物合成,苯乙醛(上调)注释到苯丙氨酸代谢通路。苯乙醛具有很强的香味[23],推测苯乙醛会对周围的假眼小绿叶蝉造成其行为或生理的影响,从而达到对害虫的抵抗。苯丙氨酸经苯丙氨酸解氨酶等酶催化合成木质素和类黄酮等,以达到茶树对假眼小绿叶蝉的抵御[24]。没食子儿茶素上调也与植物抗虫性密切相关[24]。以上研究表明,6,9,12,15十八碳四烯酸乙酯、苯乙醛、没食子儿茶素等物质在茶树响应假眼小绿叶蝉胁迫方面具有重要作用,可激活亚麻酸代谢途径、类黄酮生物合成途径、苯丙氨酸代谢途径等,以防御假眼小绿叶蝉的侵染。
12 h与24 h处理有9个差异代谢物注释到9个代谢通路中,磷脂酰丝氨酸(上调)注释到甘氨酸、丝氨酸和苏氨酸代谢通路中。十八碳炔烯醇酸(下调)和己二酸二环己酯(下调)注释到亚麻酸代谢通路中。马钱苷酸(上调)注释到单萜生物合成通路中,相比侵染12 h处理,24 h处理的马钱苷酸含量上调,表明茶树被假眼小绿叶蝉为害后可能通过释放萜类物质来达到对害虫的抵御;白杨素(上调)、没食子儿茶素(上调)和表没食子儿茶素(上调)注释到类黄酮生物合成通路中,表明黄酮等多酚类物质与茶树抗虫性密切相关。以上结果表明,萜类物质和黄酮类化合物在茶树响应小绿叶蝉吸食的应激防御方面起着重要作用,与王杰等[7]的研究一致,为选育抗虫良种并最终实现茶叶增产提供理论依据。
[1] YANG Z Y,BALDERMANN S,WATANABE N.Recent studies of the volatile compounds in tea[J].Food Research International,2013,53(2):585–599.
[2] 任倩倩.抗、感茶树品种对茶小绿叶蝉取食诱导的防御反应[D].福州:福建农林大学,2020.
[3] 赵小嫚.多组学技术探究茶树响应小绿叶蝉吸食的防御反应[D].福州:福建农林大学,2019.
[4] ZENG L T,WATANABE N,YANG Z Y.Understanding the biosyntheses and stress response mechanisms of aroma compounds in tea() to safely and effectively improve tea aroma[J].Critical Reviews in Food Science and Nutrition,2019,59(14):2321–2334.
[5] ARIMURA G I,OZAWA R,MAFFEI M E.Recent advances in plant early signaling in response to herbivory[J].International Journal of Molecular Sciences,2011,12(6):3723–3739.
[6] 张明菊,朱莉,夏启中.植物激素对胁迫反应调控的研究进展[J].湖北大学学报(自然科学版),2021,43(3):242–253.
[7] 王杰,童璐,巨学阳,等.茶树对茶小绿叶蝉为害的响应及其机制研究进展[J].福建农林大学学报(自然科学版),2021,50(2):145–154.
[8] CHO J Y,MIZUTANI M,SHIMIZU B I,et al.Chemical profiling and gene expression profiling during the manufacturing process of Taiwan oolong tea “Oriental Beauty”[J].Bioscience,Biotechnology,and Biochemistry,2007,71(6):1476–1486.
[9] LIAO Y Y,YU Z M,LIU X Y,et al.Effect of major tea insect attack on formation of quality-related nonvolatile specialized metabolites in tea() leaves[J]. Journal of Agricultural and Food Chemistry,2019,67(24):6716–6724.
[10] MEI X,LIU X Y,ZHOU Y,et al.Formation and emission of linalool in tea() leaves infested by tea green leafhopper(()Matsuda)[J].Food Chemistry,2017,237:356–363.
[11] MU D,CUI L,GE J,et al.Behavioral responses for evaluating the attractiveness of specific tea shoot volatiles to the tea green leafhopper,[J].Insect Science,2012,19(2):229–238.
[12] 董登峰.代谢物组学方法及其在植物学研究中的应用[J].广西植物,2007,27(5):765–769.
[13] 宁井铭,方骏婷,朱小元,等.基于代谢谱分析的祁门红茶加工过程中儿茶素及芳香类物质变化[J].食品工业科技,2016,37(9):127–133,138.
[14] 吕海鹏,林智,张悦,等.不同等级普洱茶的化学成分及抗氧化活性比较[J].茶叶科学,2013,33(4):386–395.
[15] 张玉叶,陈志丹,孙威江,等.茶树对小贯小绿叶蝉组成抗性的研究进展[J].茶叶通讯,2021,48(3):392–398.
[16] 王春波,吕辉,韦玲冬,等.贵定雪芽茶树高EGCG品系代谢组学的比较研究[J].湖南农业大学学报(自然科学版),2021,47(4):412–418.
[17] 周坤能,夏加发,王元垒,等.水稻恢复系M630稻瘟病抗性改良及其代谢组研究[J].植物遗传资源学报,2021,22(2):427–437.
[18] 刘馨蕴,刘年元,尚海,等.油桐花、叶、果壳代谢组成分鉴定与差异分析[J].植物生理学报,2021,57(12):2366–2378.
[19] THÉVENOT E A,ROUX A,XU Y,et al.Analysis of the human adult urinary metabolome variations with age,body mass index,and gender by implementing a comprehensive workflow for univariate and OPLS statistical analyses[J].Journal of Proteome Research,2015,14(8):3322–3335.
[20] YU G C,WANG L G,HAN Y Y,et al.ClusterProfiler:an R package for comparing biological themes among gene clusters[J].Omics:A Journal of Integrative Biology,2012,16(5):284–287.
[21] RAGUENEAU E,SHRIVASTAVA A,MORRIS J H,et al.IntAct App:a cytoscape application for molecular interaction network visualization and analysis[J]. Bioinformatics,2021,37(20):3684–3685.
[22] GUO W F,JIN L,MIAO Y H,et al.An ethylene response-related factor,GbERF-,fromimproves resistance tovia activating lignin synthesis[J].Plant Molecular Biology,2016,91(3):305–318.
[23] 王绍梅,宋文明,冷燕,等.花香型古树滇红茶挥发物质气相色谱分析[J].江苏农业学报,2021,37(4):1010–1015.
[24] ZHONG R Q,YE Z H.Transcriptional regulation of lignin biosynthesis[J].Plant Signaling & Behavior,2009,4(11):1028–1034.
Metabolomics study on the response of Duyun Maojian native tea plant to the infestation ofGothe
WANG Fen1,MU Ren1,LI Guodong2,YU Pei1,LI Jing1,CUI Baolu1,LYU Litang2*
(1.The Department of Life Science and Agriculture, Qiannan Normal University for Nationalities, Duyun, Guizhou 558000, China; 2.College of Tea Science, Guizhou University, Guiyang , Guizhou 550025, China)
To investigate how tea response toinfection, we used ultra performance liquid chromatography-quadrupole time-of-flight mass spectrometry(UPLC-QTOF-MS) to measure the profiles of 1094 metabolites in tea leaves at 0 h, 12 h and 24 h after infection. With principal component analysis(PCA), orthogonal partial least squares discrimination analysis(OPLS-DA), and fold change(FC), 6 metabolites were upregulated and 14 downregulated at 12 h comparing to 0 h, 26 metabolites wereupregulated and 15 were downregulated at 24 h compared to 0 h, and 45 metabolites were upregulated and 14 were downregulated at 24 h compared to 12 h. Two differential metabolites in phenylpropanoid biosynthesis pathway with sinapic acid related to lignin formation at 0 h and 12 h were annotated to KEGG database . Six differential metabolites whose phenylacetaldehyde and gallocatechin were annotated to phenylalanine metabolic pathway and biosynthesis of flavonoids, respectively at 0 h and 24 h were also annotated to KEGG database. It was speculated that the fragrance of phenylacetaldehyde would affect the behavior or physiology ofGothe. There were 9 differential metabolites at 12 h and 24 h annotated to 9 metabolic pathways, and loganic acid annotated to monoterpenoid biosynthesis pathway. Compared with 12 h, the increase of loganic acidin in 24 h indicated that tea plants might release terpenoids to resist theGothe. The results indicated that lignin, terpenoids, and flavonoids played important roles in the response toinfection.
tea leaf;Gothe; metabolomics; ultra performance liquid chromatography-quadrupole time-of-flight mass spectrometry; differential metabolite
王芬,木仁,李国栋,喻佩,李静,崔宝禄,吕立堂.都匀毛尖本地种茶树响应假眼小绿叶蝉侵害的代谢组学研究[J].湖南农业大学学报(自然科学版),2022,48(5):543–549.
WANG F,MU R,LI G D,YU P,LI J,CUI B L,LYU L T.Metabolomics study on the response of Duyun Maojian native tea plant to the infestation ofGothe[J].Journal of Hunan Agricultural University(Natural Sciences),2022,48(5):543–549.
http://xb.hunau.edu.cn

S571.101; S435.711
A
1007-1032(2022)05-0543-07
10.13331/j.cnki.jhau.2022.05.006
2021–11–25
2022–09–13
国家自然科学基金项目(32260080、31900486);贵州省教育厅植物学重点支持学科项目(黔学位合字ZDXK[2016]23号);贵州省科技计划项目(黔科合基础[2019]1298);贵州省教育厅项目([2015]68、QNYSKYTD2018006);黔南民族师范学院科研创新团队项目(Qnsyk201605、QNYSKYTD2018011);黔南民族师范学院项目(2019xjg0303、2018xjg0520、QNSY2018ZJ006、QNYSXXK2018005)
王芬(1984—),女,山西运城人,博士,副教授,主要从事茶树组学研究,fenmin521@163.com;*通信作者,吕立堂,教授,主要从事茶学研究,ltlv@gzu.edu.cn
责任编辑:毛友纯
英文编辑:柳正
猜你喜欢
现代临床医学(2022年4期)2022-09-29
食品安全导刊(2021年20期)2021-08-30
理化检验-化学分册(2020年5期)2020-06-15
农药科学与管理(2019年5期)2019-08-13
特别文摘(2019年3期)2019-02-28
中成药(2017年7期)2017-11-22
中成药(2017年3期)2017-05-17
质谱学报(2015年5期)2015-03-01
蚕桑茶叶通讯(2015年1期)2015-02-22
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
