
唐古特白刺叶功能性状沿气候梯度的变异特征
2022-11-12 03:29:04魏海霞霍艳玲周忠科张治国
生态学报 2022年20期
魏海霞, 霍艳玲, 周忠科, 张治国,*
1 枣庄学院旅游与资源环境学院, 枣庄 277160 2 中国科学院青藏高原研究所高寒生态学与生物多样性重点实验室, 北京 100101 3 中国林业科学研究院荒漠化研究所, 北京 100091
升温和降水分配格局发生改变是气候变化的主要特征[1]。过去近60年的气象数据显示,我国西北干旱地区温度和降水量均呈现出明显的增加趋势[2]。荒漠生态系统是干旱区普遍存在的生态系统类型,与其它生态系统相比,由于其自身的脆弱性,决定了其对气候变化更为敏感[3—5]。叶功能性状是植物为适应环境而表现出来的叶片水平功能特征,与植物对资源的获取和利用密切相关,能够较好地反映植物对气候变化的响应与适应策略[6]。因此,系统研究我国西北荒漠区优势种叶功能性状及性状相关关系沿气候梯度的变异特征对理解该地区荒漠生态系统对未来气候变化的响应具有重要意义。
比叶面积(SLA)和叶氮含量是全球叶片经济型谱中最基础的叶功能性状,分别表征了植物对光能的捕获能力和同化CO2的能力[7]。叶氮含量依据表达单位的不同分为单位重量的叶氮含量(Nmass)和单位面积的叶氮含量(Narea),其中, Nmass高低直接决定植物叶片光合能力的大小[8], Narea为Nmass与SLA之比,反映了单位叶面积的潜在光合能力[9]。沿气候梯度,在低降水或低温环境下,植物叶片变小、变厚、SLA降低、叶氮含量增加[10—13]。一方面,低温环境下,植物SLA降低(叶片密度或叶厚度增加)和叶氮含量增加有利于叶片保持较高的温度,缓解低温对与温度相关的生理化学过程的限制[14—15];另一方面,在低降水环境下,SLA降低有利于增加叶片内部水分向叶片表面扩散的距离或阻力,减少植物内部水分散失[16],叶氮含量的增加能够增加叶片内部光合作用酶的数量,提高叶片光合速率[8],从而减弱SLA降低对植物叶片光合作用能力的影响,实现在减少单叶蒸发面积的同时提高光合速率,增加水分利用效率[17]。但是, 较小的 SLA 和/或较高的叶氮含量通常会导致新叶建成成本的增加(因其木质素、蛋白质等高能值组分较高)[18—19]。单位质量和单位面积叶建成成本(CCmass、CCarea)表征了构建单位质量和面积的叶片所需要的葡萄糖当量,能够反映植物的能量利用策略和对环境的适应能力[20]。具有较高叶建成成本的物种通常具有较低的能量利用效率和生长速率,不利于植物的竞争和生存[21—22]。有研究认为,沿气候梯度,随降水减少,植物在如何提高水分利用效率(较小的 SLA 和/或较高的叶氮含量)和降低叶建成成本之间存在权衡[23],植物多种性状权衡(或协同)变化是植物对环境响应与适应的重要途径。因此,越来越多的工作开始关注气候梯度变化对叶功能性状关系的影响。例如,Wright等[24]与张治国等[25]分别研究了降水对植物叶功能性状关系的影响,发现降水会影响SLA与Nmass、CCarea与Narea的相关关系,在相同SLA和CCarea下,降水量较少地区的植物具有较高的Nmass与Narea。Xiang等[26]发现不同的生长温度对植物SLA与Nmass的协同变化有一定影响,SLA相同的叶片,温度较低地区植物的Nmass更高。因此,相较于某一单一性状,植物往往通过调节叶功能性状之间的关系并形成多个性状间的最佳功能组合来适应环境的变化。但是,上述研究大多数都是基于单个环境因子对叶功能性状及性状关系的影响,沿着环境梯度,当降水、温度和土壤条件等共同发生变化时,上述植物叶功能性状及性状关系对环境因子的响应规律发生改变,从而无法应用单个环境因子解释植物的生存适应特征[27—29],而采用多个叶功能性状和环境因子相结合的方法更有助于揭示植物对环境变化的响应与适应。
唐古特白刺(Nitrariatangutorum)为蒺黎科白刺属灌木,多见于沙漠边缘及戈壁滩等干旱半干旱荒漠地区,是我国西北荒漠地区的主要建群种之一,固沙防风效果显著[30]。近年来,有关唐古特白刺的研究主要集中在模拟增雨对其叶片形态、光合特性的影响等方面[31—33],而系统地分析唐古特白刺叶功能性状沿着气候梯度变异特征的研究还比较缺乏,并且影响其叶功能性状的主导因子也不明确[17]。因此,本研究沿着唐古特白刺的地理分布范围,选择了3个温度和降水量明显不同的区域进行调查采样,分别测定其SLA、Nmass、Narea、CCmass和CCarea,系统分析了不同降水和温度条件下唐古特白刺叶功能性状及性状相关关系的变异特征,以期为揭示唐古特白刺对未来气候变化的响应与适应机制提供科学依据。
1 材料与方法
1.1 研究区概况与研究点选择
在唐古特白刺群落天然分布区,沿气候梯度,本研究分别选择内蒙古磴口(高温,低降水)、青海都兰(低温,低降水)和青海沙珠玉(低温,高降水)3个地点进行相关调查采样(表1)。其中,磴口属于温带荒漠大陆性气候,年平均气温8.5℃,年平均降水量151mm,磴口样地位于中国林业科学研究院沙漠林业实验中心二场附近。都兰属于高原干旱大陆性气候,年平均气温3.38℃,年平均降水量207mm,都兰样地位于都兰县城附近。沙珠玉具有显著的高原大陆性气候特征,年平均气温4.48℃,年平均降水量259 mm,沙珠玉样地位于青海省共和县沙珠玉治沙试验站内。磴口、都兰和沙珠玉3个研究地点的气象资料分别来自当地气象站的观测数据(表1)。三个研究点样地的地貌特征均以半固定沙地为主,土壤类型均为风沙土,唐古特白刺为样地内优势物种,伴生种有沙鞭、沙蓬、虫实等。
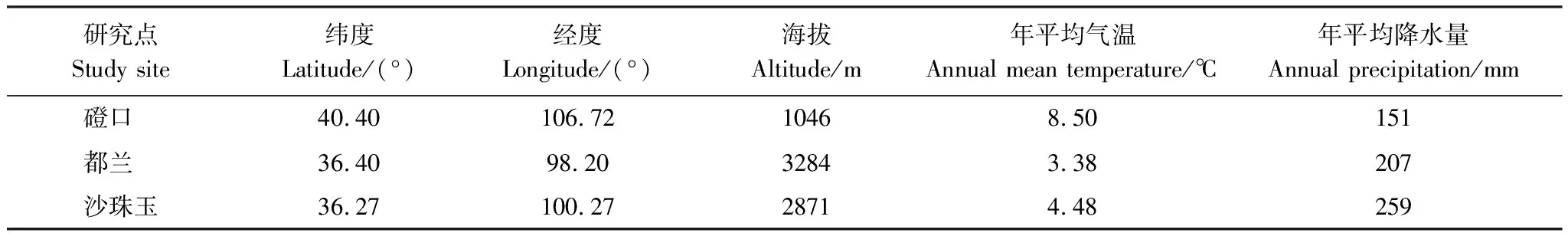
表1 采样点的地理位置及气候特征
1.2 植物与土壤样品的采集和测定
2008年7—8月,在3个研究点选择人类活动干扰较轻以及地势较为平坦的区域,分别设置3 个 10 m×10 m 调查样方,每个样方间隔 50 m 以上。在每个样方内选取4个唐古特白刺灌丛,3个研究点共计36丛。在每个唐古特白刺灌丛的冠层外部采集完全展开且保持完整的成熟叶片50—100g用于植物叶功能性状的分析。同时分别在每个样方的四角和中心位置进行土壤样品的采集,土壤样品通过土钻钻取获得,钻取深度为0—20 cm,将在同一样方中采集到的土壤均匀混合在一起作为样方水平上的土壤样品。将土壤样品自然风干碾碎后过2 mm筛,然后采用凯式定氮法测定全氮含量。
对于在3个研究点采集到的36份植物叶片样品,各随机选取50片新鲜叶进行叶面积的测定。利用扫描仪(HP Scanjet2400)扫描50片叶片的单面面积,之后采用Image Pro Plus 6.0软件对扫描图像进行处理以获得扫描叶片的实际叶面积。将扫描叶片在实验室中于70℃烘箱中烘干48h至恒重,之后用万分之一天平称重。SLA为样品叶面积总和与叶干重之比。
将采集到的每份植物叶片样品于70℃烘箱中烘干48h至恒重,用粉碎机粉碎,过80目筛,进行Nmass和CCmass的测定。以凯氏定氮法测定植物叶片样品的Nmass, Narea为Nmass与SLA的比值。CCmass采用Williams等[20]给出的计算公式进行计算:CCmass=[(0.06968Hc-0.065) × (1-A)+(kNmass/14.0067) × 180.15/24]/EG。其中Hc为热值(kJ/g);A为灰份含量(g/g);k为含氮化合物中氮的价态(硝态氮为+5, 铵态氮为-3);EG为生长效率。根据大量研究,EG一般为0.89[20]。 本研究假设植物的氮来源均为硝态氮,因为在大多数生境下硝态氮是高等植物最主要的氮源[34],而铵态氮仅在苔原地区是植物的主要氮来源[35]。公式中的灰分含量和热值分别通过直接灰化法和SDCM-Ⅲa氧弹式热量计测定。CCarea为CCmass与SLA的比值。
1.3 数据分析
通过单因素方差分析(One-way ANOVA)和均值的多重比较(Tukey)检验唐古特白刺各叶功能性状在不同研究点之间是否存在显著差异。然后通过多元线性回归的偏相关分析探讨气候因子和土壤全氮含量对唐古特白刺各叶功能性状的相对影响。采用简单线性回归方程分析SLA、Nmass、Narea、CCmass和CCarea的相关关系。上述叶功能性状关系在不同研究点之间的差异用协方差进行分析(ANCOVA)。协方差分析常用来检验几条回归线之间斜率和截距的差异。首先检验不同研究地区唐古特白刺叶功能性状关系斜率的相似性,之后分析截距的差异。
数据的显著水平均为P<0.05。所有分析均在SPSS 16.0软件中进行。
2 结果与分析
2.1 唐古特白刺叶功能性状沿气候梯度的变异特征
唐古特白刺SLA在温度较高的磴口显著高于其他两个温度较低的地区都兰和沙珠玉(磴口,(83.65±3.40) cm2/g;都兰,(69.01±3.80)cm2/g;沙珠玉,(72.75±4.68)cm2/g),而在温度差异不大、降水明显不同的都兰和沙珠玉之间差异不显著。Nmass在不同研究地区之间存在显著差异,其中Nmass在都兰最高(40.37±1.92)mg/g,其次为磴口(36.57±3.10)mg/g,最低的为沙珠玉(32.56±1.36)mg/g。唐古特白刺Narea在低温低降水的都兰最高(5.86±0.22)g/m2,在其他两个研究地区磴口和沙珠玉较低(4.37±0.25)g/m2和(4.48±0.16)g/m2。唐古特白刺CCmass在不同研究地区之间差异显著,其中,CCmass在3个研究地区从大到小依次为磴口(1.04±0.07)g/g >沙珠玉(0.96±0.05)g/g >都兰(0.86±0.05)g/g。与CCmass不同, CCarea在不同研究地区之间并没有表现出显著差异(图1)。
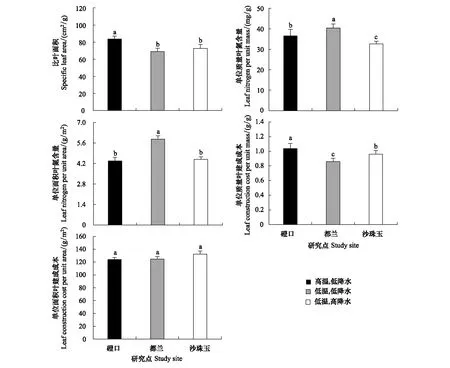
图1 不同研究地区唐古特白刺叶功能性状的比较Fig.1 Comparison of leaf functional traits of N. tangutorum among study sites不同字母表示叶功能性状在不同研究地区之间存在显著差异(P<0.05)
关于温度、降水和土壤全氮含量对唐古特白刺叶功能性状的相对影响,偏相关分析表明,年平均温度是决定唐古特白刺SLA变化的主要因子,二者之间呈极显著正相关关系,随着温度的增加,SLA增加(表2)。年平均温度和年平均降水量是决定唐古特白刺Nmass和Narea变化的主要因子,Nmass和Narea与温度和降水量均呈极显著负相关关系,即随着年平均温度或年平均降水量的减少,Nmass和Narea增加(表2)。唐古特白刺CCmass与年平均降水量呈显著正相关关系,与年平均气温呈极显著正相关关系,年平均气温是影响CCmass变化的主要因素,随着温度增加,唐古特白刺CCmass增加(表2)。与其它叶功能性状不同,年平均降水量、年平均气温和土壤全氮含量对唐古特白刺CCarea均没有显著影响(表2)。
2.2 唐古特白刺叶功能性状关系沿气候梯度的变异特征
在3个研究地区,唐古特白刺SLA与Nmass均呈极显著正相关关系。协方差分析结果表明,SLA与Nmass关系的斜率在3个研究地区之间差异不显著(P>0.05),但是,其截距在都兰和其他两个研究地区磴口和沙珠玉间有显著差异(P<0.05),而磴口和沙珠玉间截距差异不显著(P>0.05)。对于唐古特白刺来说,在给定的SLA下,低温低降水的都兰地区具有更高的Nmass,或在给定的Nmass下,都兰具有更小的SLA。SLA与Narea和CCarea在沙珠玉呈极显著负相关关系,但在其他两个研究地区相关性不显著。SLA与CCmass在3个研究地区均没有表现出显著相关关系(图2)。
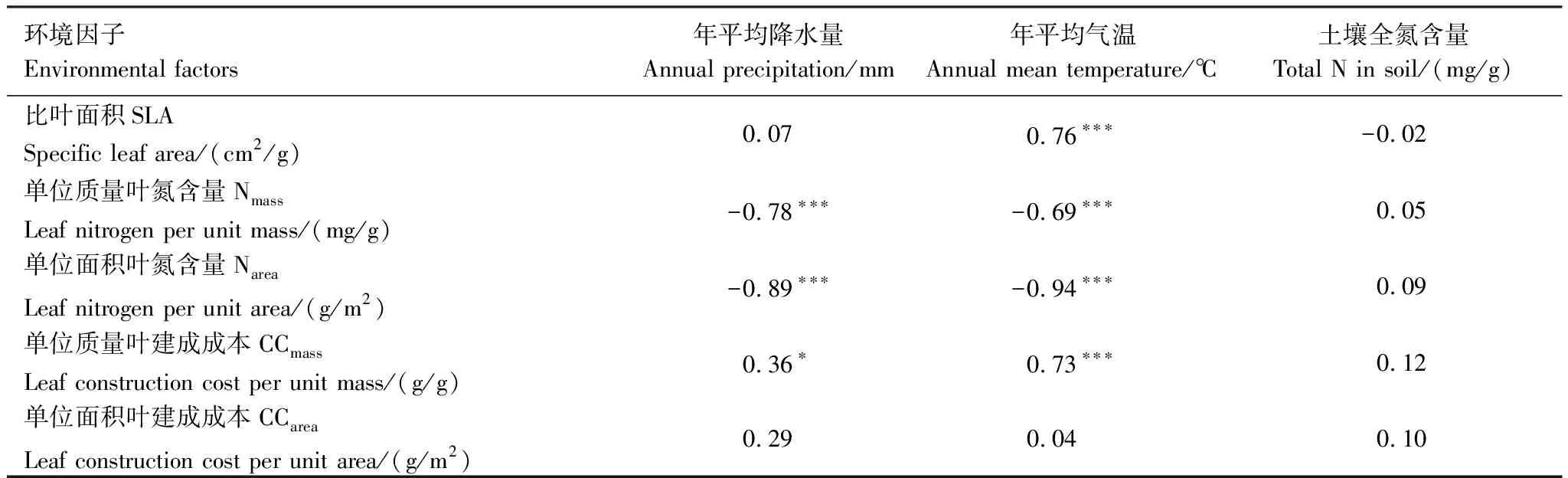
表2 不同研究地区唐古特白刺叶功能性状与环境因子的偏相关分析
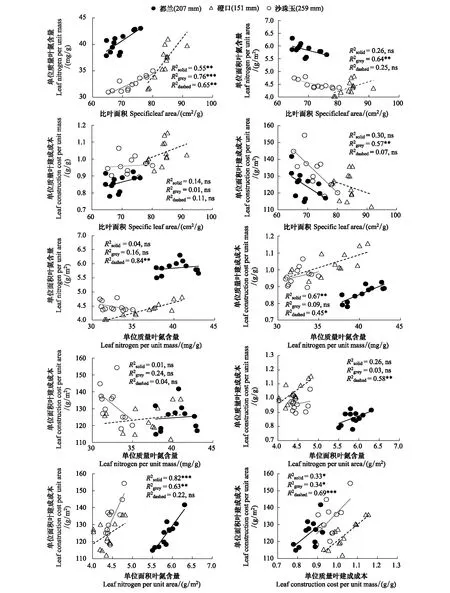
图2 不同研究地区唐古特白刺叶功能性状的相关关系Fig.2 Relationships of leaf functional traits within species of N. tangutorum under the three study sites
在都兰地区,Nmass与Narea、Narea与CCmass均呈极显著正相关关系,但两组叶性状关系在其他两个研究地区磴口和沙珠玉均不显著。CCmass与Nmass在都兰和磴口呈显著正相关关系,CCarea与Narea在都兰和沙珠玉呈极显著正相关关系。与SLA-Nmass关系在不同研究地区之间的变化相似,CCmass-Nmass和CCarea-Narea关系的斜率在不同研究地区之间差异不显著(P>0.05),但是,其截距在都兰和其他两个研究地区磴口和沙珠玉间有显著差异(P<0.05)。对于唐古特白刺来说,在给定的CCmass和CCarea下,低温低降水的都兰地区具有更高的Nmass和Narea。在3个研究地区,唐古特白刺CCmass与CCarea均呈显著正相关关系(图2)。
3 讨论与结论
3.1 降水和温度对唐古特白刺叶功能性状的相对影响
叶功能性状与植物的生长对策及资源利用效率紧密关联,研究叶性状沿环境梯度的变化特征能为理解植物对环境变化的响应与适应机制提供一种简便可行的测定指标[6—7]。在本研究中,沿气候梯度,唐古特白刺SLA在低温低降水的都兰研究点和低温高降水的沙珠玉研究点之间差异不显著,但都显著低于高温低降水的磴口研究点(图1)。偏相关分析进一步表明,唐古特白刺SLA主要受温度影响,降水没有成为其限制因子(表2)。因此, 在降雨不成为SLA变化的决定性因子时, 唐古特白刺SLA的变化主要是对温度变化的一种适应策略。较小的SLA意味着叶片会形成厚度较大而面积较小的叶片,一方面,低温环境下,小叶片的呼吸和蒸腾成本更低, 可以降低植物的维持消耗[36];其次,有研究发现大叶片与环境的温差比小叶片更大,从而使得大叶片在寒冷地区更容易遭受霜冻的破坏[37];此外,较厚的叶片具有保温和耐辐射的作用,能更好的适应寒冷气候和近日照的环境条件[14]。在本研究中,唐古特白刺Nmass和Narea在低温低降水的都兰研究点最高(图1)。偏相关分析表明,沿气候梯度,温度和降水对唐古特白刺Nmass和Narea均有显著影响,随着温度或降水量的减少,唐古特白刺Nmass和Narea增加(表2),这与以往诸多研究结果相同[10—13],是植物对低温和低降水环境的一种适应策略。叶片中的氮素主要分布于叶绿体当中,在低降水环境下,叶氮含量的增加直接提高叶片光合系统中酶的含量和活性,植物通过加强对光合酶的投资以促进光合速率(导致胞内二氧化碳浓度降低),提高植物的水分利用效率,有利于植物对水分胁迫的适应[8, 24]。此外,叶氮含量的增加也能够增加叶片内部非光合器官或组织氮的投入,如细胞壁中的氮含量,提高细胞内部的渗透压,因此较高的叶氮含量对干旱区植物光合作用中水分的保护具有重要意义[8]。在低温环境下,与温度相关的植物生理化学过程受温度的影响较大,如:高氮酶的生物化学效应会在低温时效率降低,而植物则可通过提高全氮含量来弥补效率的下降,缓解低温带来的负面影响[15]。
目前关于温度和降水对植物叶建成成本影响的研究较少,且没有一致的结论。有研究认为植物在受到干旱或低温胁迫时,植物叶建成成本增加[18—19];但也有研究认为,植物在受到环境因子胁迫时,植物叶建成成本会降低以提高自身对环境的适应性[38]。Villar & Merino[34]测定了14个不同生境下(从荒漠到雨林)162个物种的建成成本,发现冻原和荒漠植物分别具有最高和中间水平的建成成本。张治国等[25]分析了毛乌素沙地油蒿叶建成成本沿降水梯度的变化规律,结果却发现降水对植物叶建成成本并没有显著影响。在本研究中,唐古特白刺CCmass在不同研究地区之间差异显著,偏相关分析表明降水和年平均气温对CCmass均有显著影响,随着降水的减少和温度的降低,CCmass降低。CCmass主要由叶片化学特性决定,因此,不同降水和温度条件下植物CCmass的变化可能与叶片生化组分有关。理论上讲,干旱和低温可能会导致一些高能化合物的累积,这些化合物可使植物组织不易变形(木质素)或防止水分的散失(脂质)[19]。但也有研究发现,植物叶片中高能量投资组分与低能量投资组分呈正相关关系,如蛋白质和矿质元素,高能量投资组分之间呈负相关关系,如蛋白质和油脂等[21, 39]。在本研究中,降水和温度较低的都兰地区具有最低的热值与最高的灰份含量是导致该地区CCmass显著低于其他两个研究地区的主要原因。
3.2 唐古特白刺叶功能性状关系沿气候梯度的变异及其生理生态学意义
沿着降水梯度,Wright 等[24]和李永华等[40]发现 SLA-Nmass关系的斜率没有发生变化,而截距在低降水量地区发生上移, SLA-Nmass关系的截距表征植物的Narea,即低降水量地区植物具有更高的 Narea, 这是植物应对有限水分资源的一种保护策略。沿着降水梯度,Narea与叶 δ13C 值(通常与水分利用效率呈正相关)普遍存在正相关关系[41—42]。由于上述研究是跨越物种进行的,所以目前仍不清楚SLA-Nmass关系沿降水梯度的变化是由降水引起的还是由物种更替造成的。目前关于植物种内叶性状关系沿环境梯度变化的研究很少,而对这些关系的认识对于从生理生态角度方面深入理解植物对环境的适应性具有重要意义。在本研究中,沿气候梯度,唐古特白刺SLA-Nmass关系在低温低降水的都兰地区和其它两个低温高降水的沙珠玉地区以及高温低降水的磴口地区之间同样发生了平移,导致在相同SLA下,都兰地区具有更高的Nmass,这与以往研究结果一致。但是,胡梦瑶等[28]分析了青藏高原干旱、半干旱草地优势种紫花针茅叶功能性状沿着降水梯度的变化规律,发现紫花针茅SLA-Nmass关系在低降水量区和高降水量区之间并没有出现明显的平移,这可能与研究区域的综合环境因子有关,揭示了环境因子对植物叶功能性状的影响机制较为复杂。由于决定比叶面积大小的叶片厚度和叶片密度对Nmass的影响可能存在差异,SLA与Nmass的相关关系可能在很大程度上取决于叶厚度和叶密度的变化及其对SLA的贡献程度[17,28]。因此,进一步加强对植物解剖特征的研究可能有助于更好地理解SLA-Nmass关系沿着环境梯度变化的生理生态学意义。
SLA-Nmass关系沿气候梯度的位移导致降水和温度较低的地区具有较高的Narea。但是,较低的 SLA 和/或较高Nmass通常会导致新叶建成成本的增加(因其木质素、蛋白质等高能值组分较高),具有较高叶建成成本的物种通常具有较低的能量利用效率和生长速率,不利于植物的竞争和生存[21—22]。在本研究中,沿着气候梯度,SLA的降低以及Nmass和Narea的增加并没有导致唐古特白刺CCmass和CCarea的增加。CCmass-Nmass和CCarea-Narea关系在降水和温度较低的都兰地区和其它两个降水或温度较高的地区之间发生了位移,在相同的CCmass和CCarea下,都兰地区具有更高的Nmass和Narea。这一结果表明唐古特白刺能通过调节叶功能性状之间的关系来适应环境的变化,并形成性状间的最佳功能组合。
猜你喜欢
草原歌声(2019年1期)2019-07-25 07:30:50
农学学报(2018年10期)2018-11-01 01:19:30
中成药(2018年7期)2018-08-04 06:04:06
西夏学(2018年1期)2018-04-29 09:09:02
西夏研究(2017年1期)2017-07-10 08:16:55
西藏研究(2016年1期)2016-06-22 11:09:36
西藏研究(2016年4期)2016-06-05 11:31:19
西南农业学报(2016年5期)2016-05-17 05:42:28
海峡旅游(2015年2期)2015-08-17 20:04:21
天然产物研究与开发(2014年6期)2014-04-27 14:15:58
