
硝酸镧对脐橙叶片抗氧化酶活性的影响*
2022-11-12 00:41高柱陈璐毛积鹏卢玉鹏张小丽樊志鹏王小玲
中国果树 2022年10期
高柱,陈璐,毛积鹏,卢玉鹏,张小丽,樊志鹏,王小玲
(1 江西省科学院生物资源研究所,南昌330096)(2 井冈山生物技术研究院)
我国拥有丰富的稀土矿产资源,稀土元素齐全,成矿条件优越,储量居世界首位,是名副其实的稀土资源大国[1-2]。稀土农用是我国重要的创新之举,也是我国具有特色的研究成果之一,其施用方法简便且经济效益显著,在粮食[3]、水果[4]、蔬菜[5]等作物上得到了较为广泛的应用。稀土元素具有独特的理化性质和生理活性,可与质膜上的生物大分子如蛋白、磷脂等结合形成比较稳定的复合物,从而影响植物体内生理生化代谢过程[6]。
稀土元素在农业上的应用可归纳为以下3 个方面。一是促进植物生长发育。施用适当浓度的稀土元素可以打破种子休眠、促进种子萌发、促进植物芽和根系的生长;水稻[7]、小麦[8]、丹参[9]种子播种前经硝酸稀土浸泡处理后,显著提高了种子发芽率及发芽指数,同时促进了小麦苗期旺盛生长、丹参幼苗干物质积累与茎叶生长。二是减轻胁迫伤害,提高植物抗性。一方面,稀土离子能够提高渗透调节物质的积累和细胞膜的保护功能,维持细胞膜的通透性和稳定性,增强作物对逆境或胁迫的抵抗能力[10];另一方面稀土离子可以调节植物自身抗氧化酶活性,清除过多活性氧与自由基,缓解不良环境造成的损伤[11-12]。硝酸稀土处理藜麦幼苗后提高了抗氧化酶活性,调节了渗透物质含量,缓解了盐胁迫的损伤[13]。三是提高产量,改善品质。施用适当浓度的稀土元素可以提高植物对养分的吸收、转化和利用效率,还可促进植物自身叶绿素形成、增强光合效率,为提高产量与改善品质奠定物质基础。适宜浓度的硝酸稀土处理甜瓜[14]、草莓[15]后,明显提高了单果重和维生素C 含量,甜瓜果实可溶性固形物、可溶性糖含量也得到了提高。
镧与其他稀土元素相比,具有化学性质活泼、水合半径离子小、生物活性高等特性,被认为是作物的生理活性物质,在调节植物生理活动中起到重要作用[16]。植物在逆境胁迫条件下,增强酶促防御系统中保护酶的活性,如超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、谷胱甘肽还原酶(GR)、抗坏血酸氧化酶(APX)等抗氧化酶,可清除植物体内的活性氧及自由基,缓解胁迫伤害[2]。稀土元素镧不仅可以调节植物生长,还能促进逆境条件下保护酶活性的增加、降低细胞膜过氧化作用下丙二醛的产生,增强植物抵御逆境能力[12]。镧的合理使用对植物生长发育和品质改善具有明显促进作用,研究表明,稀土元素镧对植物生理调控表现出“低浓度硝酸镧促进、高浓度硝酸镧抑制”效应,高浓度硝酸镧处理时镧的重金属特性会抑制植物体内特定酶或结合蛋白的活性,甚至产生毒害作用[17]。
由于稀土元素镧在植物中的作用机理较为复杂,不同植物对稀土元素镧浓度的耐受性不尽相同,因此,不同植物及其组织、器官的最佳使用浓度还需要不断探索。本研究以2 年生纽荷尔脐橙嫁接苗为试验材料,采用不同浓度的硝酸镧溶液喷施叶片,通过测定喷施后24 h内不同时间段脐橙叶片中抗氧化酶(SOD、POD、CAT、GR 和APX)的活性,研究其活性变化规律,分析不同浓度稀土元素镧对脐橙叶片抗氧化酶活性的影响,为稀土元素的合理使用及其对脐橙生理代谢及品质调控提供科学依据。
1 材料与方法
1.1 试验材料
试验在江西省科学院园艺植物高值利用研究室奉新试验基地(海拔64.35 m,北纬28°40′17″,东经115°18′31″)进行。试验材料为2 年生纽荷尔脐橙嫁接苗,于2020 年2 月移栽至口径60 cm、底径31 cm、高35 cm 的花盆中,每盆栽植1 株嫁接苗。供试盆栽土取自基地耕作层,将土壤混匀之后,每盆装10 kg 土进行移栽定植。
1.2 试验方法
试验设置4 个处理:50、150、300 mg/L 硝酸镧处理,以喷施等量清水为对照(CK)。2020 年7月中旬,选择晴朗天气的8:00 喷施,每个处理喷施5 株幼树,3 次重复。于喷施当天8:00、10:00、12:00、16:00、20:00 和次日8:00(处理时间记录为0、2、4、8、12、24 h)采集植株中上部当年生嫩叶。将采集的叶片剪碎混匀,液氮冷冻后-80 ℃保存待测。
1.3 测定项目与方法
称取脐橙叶片样品1.0 g,液氮研磨,加入9 mL PBS 缓冲液(10 mol/L,pH 值7.2)匀浆抽提10 min。将制备好的10%匀浆液在4 ℃条件下离心20 min(8 000 r/min),取上清液置冰上待测[18]。叶片中抗氧化酶SOD、POD、CAT、GR、APX 活性均采用上海优选生物科技有限公司的ELISA 检测试剂盒进行测定,3 次重复。
1.4 数据分析
利用Microsoft Excel 2010 软件处理数据及制图,采用SPSS 20.0 软件进行数据统计分析和差异显著性检验。
2 结果与分析
2.1 不同浓度硝酸镧处理对脐橙叶片SOD 活性的影响
SOD 广泛存在于动植物和微生物中,能将超氧化物阴离子自由基O2-迅速歧化为过氧化氢和分子氧,是在清除活性氧过程中第一个发挥作用的抗氧化酶[19]。如图1 所示,CK 的脐橙叶片SOD 活性在24 h 内呈升高的趋势,而3 个硝酸镧处理的叶片SOD 活性均呈先升高后降低的趋势,且峰值均出现在处理后4 h,此时,50、150 mg/L 硝酸镧处理的叶片SOD 活性分别为13.85、14.23 U/g,均显著高于CK,300 mg/L 硝酸镧处理的叶片SOD 活性与CK 无显著差异。在处理后12 h,3 个硝酸镧处理的叶片SOD 活性均低于CK,其中300 mg/L 硝酸镧处理差异达显著水平。说明50、150 mg/L 硝酸镧处理前期可促进叶片SOD 活性的增加,加快活性氧的清除并降低自由基伤害,而300 mg/L 硝酸镧处理对叶片SOD 活性的调节作用不明显。

图1 不同浓度硝酸镧处理叶片SOD 活性的变化
2.2 不同浓度硝酸镧处理对脐橙叶片POD 活性的影响
POD 是以过氧化氢为电子受体催化底物氧化的酶,具有调节自由基和活性氧浓度而防止膜脂过氧化的作用,是一种重要的防御保护酶[20]。如图2所示,所有处理的叶片POD 活性在24 h 内均呈先升高后降低的趋势,3 个硝酸镧处理叶片POD 活性峰值与CK 相比均有所提前,150、300 mg/L 硝酸镧处理的峰值提前8 h,50 mg/L 硝酸镧处理的峰值提前4 h。处理后2、4 h,3 个硝酸镧处理叶片POD活性均显著高于CK,且以300 mg/L 硝酸镧处理最高;处理后8、12、24 h,50 mg/L 硝酸镧处理叶片POD 活性均高于其他处理,且均显著高于150、300 mg/L 硝酸镧处理。说明硝酸镧处理能改变脐橙叶片POD 活性变化规律,150 mg/L 硝酸镧处理的叶片POD 活性下降最快。

图2 不同浓度硝酸镧处理叶片POD 活性的变化
2.3 不同浓度硝酸镧处理对脐橙叶片CAT 活性的影响
CAT 存在于细胞的过氧化物体内,专一催化底物过氧化氢分解成氧和水[21],在清除活性氧、维持活性氧代谢平衡方面具有重要作用[22]。从图3 可以看出,50 mg/L 硝酸镧处理的叶片CAT 活性呈升高—降低—升高的趋势,150 mg/L 硝酸镧处理呈先降低后升高的趋势,而300 mg/L 硝酸镧处理呈降低—升高—降低的趋势。CK 的叶片CAT 活性最低值(1.28 U/g)出现在处理后8 h,分别是50、150、300 mg/L 硝酸镧处理的53.56%、61.24%和35.75%。50、150 mg/L 硝酸镧处理的最低值和300 mg/L 硝酸镧处理的峰值均出现在处理后12 h。处理后2、4 h,50 mg/L 硝酸镧处理的叶片CAT 活性显著高于其他处理;处理后8、12 h,300 mg/L 硝酸镧处理的叶片CAT 活性显著高于其他处理;处理后24 h,3 个硝酸镧处理的叶片CAT 活性均显著低于CK。表明50 mg/L 硝酸镧处理前期可促进CAT 活性的升高,而300 mg/L 硝酸镧处理后期可促进CAT 活性的升高。
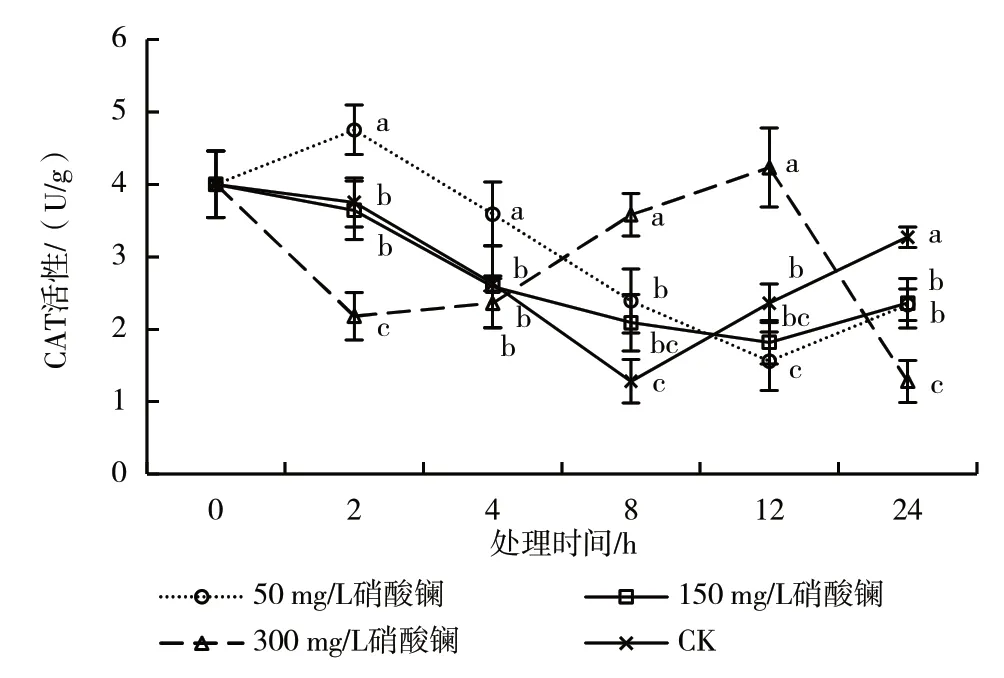
图3 不同浓度硝酸镧处理叶片CAT 活性的变化
2.4 不同浓度硝酸镧处理对脐橙叶片GR 活性的影响
由图4 可以看出,3 个硝酸镧处理均提高了叶片GR 活性,且随处理时间的延长均呈先升高后降低的趋势,峰值均出现在处理后4 h,其中150、300 mg/L 硝酸镧处理的叶片GR 活性始终显著高于CK。随着硝酸镧处理浓度的升高,叶片GR 活性呈升高趋势,最高值出现在300 mg/L 硝酸镧处理后4 h,达26.08 U/g,是CK 的2.45倍,此时,150、300 mg/L硝酸镧处理的叶片GR 活性均显著高于50 mg/L 硝酸镧处理。可见,硝酸镧具有提高叶片GR 活性的作用,且硝酸镧浓度越高其促进作用越显著。

图4 不同浓度硝酸镧处理叶片GR 活性的变化
2.5 不同浓度硝酸镧处理对脐橙叶片APX 活性的影响
由图5 可知,3 个硝酸镧处理均降低了叶片APX活性,其中50、150 mg/L 硝酸镧处理的降低效果更明显,且主要出现在处理后4~12 h;CK 和300 mg/L硝酸镧处理均呈先升高后降低的趋势,处理后4 h,50、150、300 mg/L 硝酸镧处理的叶片APX 活性分别为CK(17.25 U/g)的39.07%、39.13%和56.12%。硝酸镧处理浓度的升高对叶片APX 活性的影响表现为前期(处理后2 h)小、后期大,处理后4~24 h,3 个硝酸镧处理的叶片APX 活性均显著低于CK,其中,150 mg/L 硝酸镧处理对叶片APX 活性的抑制效果最为明显。

图5 不同浓度硝酸镧处理叶片APX 活性的变化
3 讨论与结论
植物长期进化形成一套成熟的抵御逆境胁迫的抗氧化系统[23],正常条件下,植物通过抗氧化剂与抗氧化酶系统共同维持细胞内活性氧代谢的动态平衡[24]。植物受到胁迫时,细胞稳态遭到破坏,诱导产生超氧阴离子、羟自由基、单线态氧、过氧化氢、脂类过氧化物等化学性质活泼的活性氧,当体内积累大量活性氧时,抗氧化酶系统启动自我防御功能,通过调节抗氧化酶活性清除过多的活性氧,以维持体内活性氧产生与清除的动态平衡,控制膜过氧化程度,增强胁迫抵御能力[25]。
3.1 硝酸镧处理浓度对抗氧化酶活性的影响
SOD、POD、CAT 的协同作用组成了植物防御系统中的一个抗氧化链条[26]。SOD 在活性氧清除过程中首先发挥作用,将超氧化物阴离子迅速歧化为过氧化氢和分子氧,而歧化反应产生的过氧化氢会立即被POD 和CAT 分解为完全无害的水,从而降低自由基对细胞造成的功能性伤害[27]。不同浓度硝酸镧处理在不同时间段对脐橙叶片中SOD、POD、CAT 活性的影响差异较大。叶片SOD 活性在处理后4 h 出现峰值,且随着处理浓度的升高呈先升高后降低的趋势,150 mg/L 硝酸镧处理的叶片SOD 活性最高。300 mg/L 硝酸镧处理的叶片POD 活性峰值出现在处理后4 h,而50 mg/L 硝酸镧处理的叶片POD活性峰值出现在处理后8 h,且均显著高于其他处理。叶片CAT 活性在处理后表现出显著差异,处理后2 h,50 mg/L 硝酸镧处理的叶片CAT 活性显著高于CK,300 mg/L 硝酸镧处理的叶片CAT 活性显著低于CK,而150 mg/L 硝酸镧处理的叶片CAT 活性与CK 无显著差异,说明硝酸镧对脐橙叶片中抗氧化酶活性同样显现了“低浓度硝酸镧促进、高浓度硝酸镧抑制”的效应[17]。硝酸镧处理能够提高抗氧化酶活性,加快活性氧与丙二醛的清除,从而降低膜脂的过氧化,起到保护细胞膜结构的作用。在本研究中,能够促进叶片SOD 活性增加的适宜硝酸镧浓度为150 mg/L,而促进叶片POD 和CAT 活性增加的适宜硝酸镧浓度为50 mg/L。
抗坏血酸-谷胱甘肽(AsA-GSH)循环作为植物清除活性氧的通路之一,通过调控抗氧化非酶物质含量和抗氧化酶活性来减轻氧化应激损伤,在植物应对胁迫时的防御体系中扮演重要角色[20]。GR 和APX 是AsA-GSH 循环中的主要酶系,维持氧化还原水平、保护叶绿体和其他细胞组分免受活性氧的毒害,从而提高植物氧化胁迫的耐受性[28]。本研究中,叶片GR 活性随硝酸镧处理浓度的升高而升高,且大多数时期显著高于CK,处理后4 h 出现峰值,其中300 mg/L 硝酸镧处理的叶片GR 活性最高达26.08 U/g。提高叶片GR 活性有助于维持或增加细胞内谷胱甘肽含量,以维系膜蛋白结构的稳定性[29]。本研究表明,不同浓度硝酸镧处理的叶片APX 活性大多数时期显著低于CK,50、150 mg/L 硝酸镧处理对叶片APX 活性的降低效果更明显。高活性APX会降低抗坏血酸含量,在AsA-GSH 循环中APX 以抗坏血酸为电子供体,还原过量的过氧化氢,硝酸镧处理后降低了脐橙叶片APX 活性,低活性APX更有利于维持细胞内抗坏血酸含量及氧化还原的平衡[30]。硝酸镧处理可降低脐橙叶片APX 活性、提高叶片GR 活性,该结论与Shan等[15,31]、李进等[29]的研究结论一致。抗坏血酸、谷胱甘肽等非酶促抗氧化剂直接与活性氧接触发生反应,同时还可作为活性氧清除的催化剂,加速活性氧清除过程[31],平衡细胞内AsA/DHA、GSH/GSSG 氧化还原水平,低活性APX 可减少抗坏血酸降解,高活性GR 可促进谷胱甘肽合成,能够减轻不利环境对脐橙叶片造成的伤害。因此,能够提高脐橙叶片GR 活性的适宜硝酸镧浓度为300 mg/L,而降低APX 活性的适宜硝酸镧浓度为150 mg/L。
研究表明,稀土元素镧对植物生长发育的影响取决于浓度,过量的镧将抑制植物生长、破坏细胞结构和扰乱生理代谢;低浓度的镧可以促进根系生长、种子萌发、茎叶生长以及介导次生代谢、调节氮同化等,同时可以缓解植物环境胁迫下的氧化损伤[32]。本研究结果显示,300 mg/L 硝酸镧处理促进了叶片GR 活性升高,抑制了SOD 活性增加;150 mg/L 硝酸镧处理提高SOD 活性的效果最佳,同时对叶片APX 活性的抑制效果最好;50 mg/L 硝酸镧处理提高了叶片POD 活性,对叶片APX 活性的抑制效果明显优于300 mg/L 硝酸镧处理,同时在处理前期提高了叶片CAT 活性,这可能与抗氧化酶参与的生理代谢调控途径不同有关。
3.2 硝酸镧处理时间对抗氧化酶活性的影响
硝酸镧处理脐橙叶片的时间长短对SOD、POD、CAT、GR 活性的影响差异较大,3 个硝酸镧处理的叶片SOD 活性均在处理后4 h 出现峰值;300 mg/L硝酸镧处理的叶片POD 活性在处理后4 h 出现峰值,而50 mg/L 硝酸镧处理POD 活性峰值出现在处理后8 h;50 mg/L 硝酸镧处理的叶片CAT 活性峰值出现在处理后2 h,300 mg/L 硝酸镧处理的峰值出现在处理后12 h;3 个硝酸镧处理的叶片GR 活性峰值出现在处理后4 h。
综上所述,硝酸镧处理能够促进脐橙叶片抗氧化能力的提高,通过抗氧化酶和抗氧化剂共同作用,维持细胞内活性氧产生与消除的动态平衡,增强脐橙叶片对环境胁迫的耐受性。
猜你喜欢
今日农业(2022年3期)2022-11-16
科学导报(2022年11期)2022-03-03
中学化学(2019年2期)2019-07-08
小猕猴学习画刊(2017年5期)2017-05-25
天津诗人(2017年2期)2017-03-16
江苏农业科学(2016年8期)2017-02-15
风湿病与关节炎(2016年12期)2017-01-14
江苏农业科学(2016年7期)2016-10-20
中学生数理化·高三版(2016年9期)2016-05-14
农产品市场周刊(2015年4期)2015-04-03
