
岷江干旱河谷两种主要人工林林下豆科物种组成及植被多样性
2022-11-11 03:59贺维杨育林高鹏张炜尤继勇张好
四川林业科技 2022年5期
贺维, 杨育林, 高鹏, 张炜, 尤继勇, 张好
1. 四川省林业科学研究院,四川 成都 610081;2. 理塘县林业和草原局,四川 理塘 624300
干旱河谷是青藏高原南、东、北边缘高山峡谷地带植被退化严重形成的独特生态景观[1],主要分布于长江上游主要支流的中上游,生态地位突出,是保障长江上游生态屏障的关键[2]。干旱河谷独特的环境条件形成了独特的植被,主要为灌丛、草丛,植物呈多毛、具刺、叶小、质厚、低矮或匍匐生长等典型旱生特征[3]。20世纪80年代以来,随着天然林保护、防护林、退耕还林、干旱河谷治理等工程的实施[4],营造了成片的岷江柏(Cupressus chengiana)和辐射松(Pinus radiata)人工林,有效恢复了部分地区的植被覆盖,总体缓解了干旱河谷植被稀疏且退化严重的现状。过去已有学者对干旱河谷造林树种选择、林地生产力、物种多样性、造林成效等进行了一系列研究[5-10],发现由于对干旱河谷植被演替规律认识不足、植被恢复多以营造人工林为主等,部分人工林不仅没达到预期效果,甚至有加剧生态系统退化的趋势[11,12]。研究认为,在干旱河谷恶劣的环境条件下,以保护植被更新能力为前提的恢复措施,才可能真正有效[13]。
林下植被作为森林生态系统的一个重要组成部分,在维护森林的多样性、生态功能稳定性和持续立地生产力方面具有独特的功能和作用[14,15]。人工林森林群落结构、生物种类简单,生态系统十分脆弱,维护地力的能力下降,促进林下植被的发育,对整个人工林生态系统的稳定性发展意义重大[16]。其中,豆科植物是干旱河谷植被的特殊功能群[3],具有较强的生存能力,同时能够为其他植物生长创造有利的微生境条件,在保持植被结构完整性、防止水土流失等方面都具有不可替代的作用[17]。因此,本研究以造林20年后的岷江柏和辐射松人工林为研究对象,通过分析豆科物种组成及植被多样性,对比评价两种人工林,为干旱河谷的生态恢复提供理论依据。
1 材料与方法
1.1 研究区概况
研究区域位于汶川、理县,属于岷江流域干暖河谷区。该区域风速大、“焚风效应”显著;光照充足,年日照时数多在2 000 h以上;温差大,1月和7月平均温差20℃以上;气候干燥,年均降雨量大多不足600 mm,年均蒸发量是降水量的3~4倍;地形切割强烈,山高坡陡、河谷深邃、相对高差大多超过1 000 m;地质破碎,滑坡、泥石流、崩塌等地质灾害频发;土壤类型主要为山地褐土,土壤贫瘠,土层薄、石粒含量高、有机质含量低;植被盖度极低,主要植被类型为小叶灌丛等。
1.2 调查方法
样地设置见下表,以造林20年后的岷江柏和辐射松人工纯林为研究对象,按照样地选择标准,分别设置实验样地,调查其豆科植物组成及林下植被多样性。样地为阴坡,海拔2 000 m以下,包括下、中、上不同坡位,坡度25~30°。每个样地设置乔木调查样方3个,每个样方大小为20 m×20 m;沿样方四个顶点方向设置4个灌木调查样方,大小为3 m×3 m;在每个灌木样方内,沿对角线设置5个草本调查样方,大小为1 m×1 m。在植物生长季的6—8月份开展样地调查,测定和统计内容有灌木层、草本层和林下植被生物量[8],并统计样方中出现的所有豆科植物,分别记录株数、高度、盖度,每种豆科挖取能够代表平均长势的植株3株,观察根瘤情况,分别将地上和地下部分取样带回实验室,于80℃烘干,称重测定生物量。
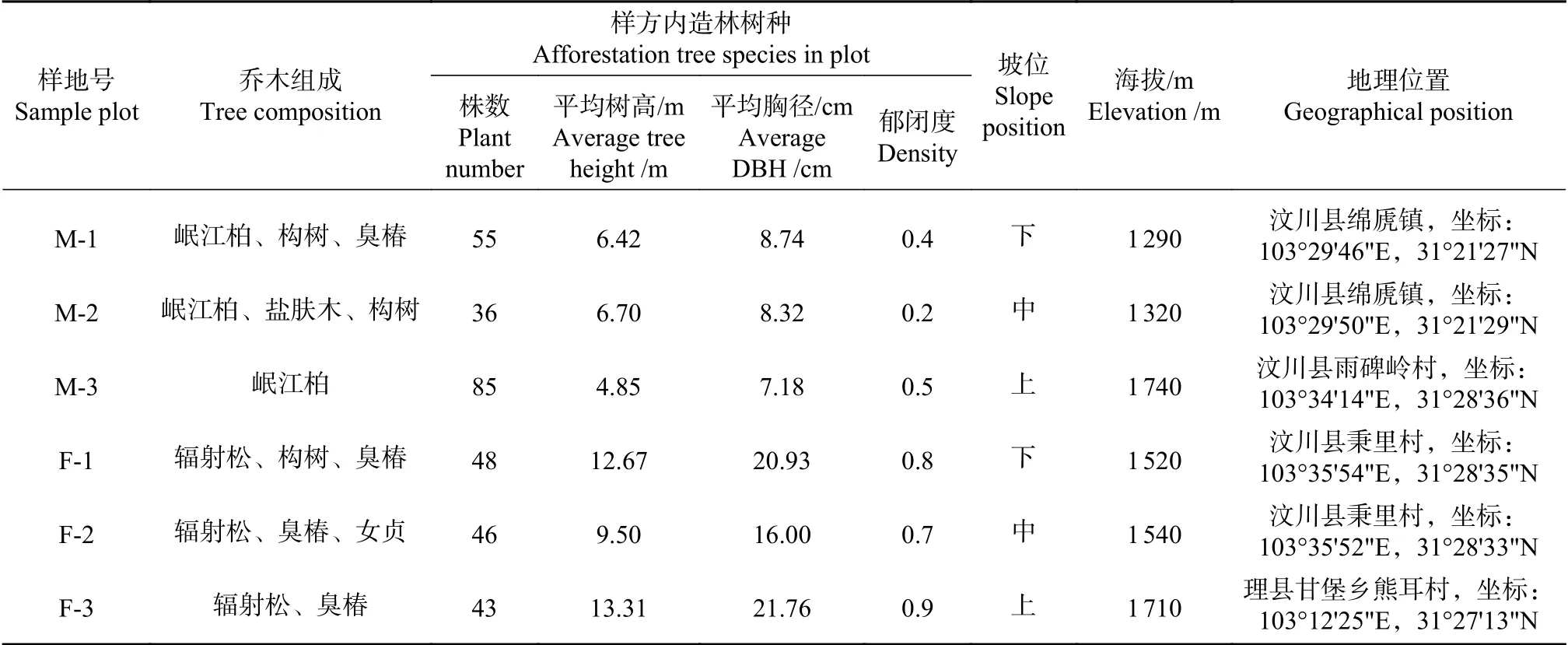
表 1 岷江柏和辐射松人工林样地情况表Tab. 1 Sample plots of Cupressus chengiana and Pinups radiata plantations
1.3 物种多样性计算
根据样地资料,计算各物种的相对密度、相对频度和相对显著度(相对盖度)和重要值(important value,IV)。采用Margalef丰富度指数、Simpson多样性指数、Shannon-wienner信息指数、Pielou均匀度指数来综合评价人工林群落的物种多样性[8]。
用Excel软件进行基础数据统计;单因素方差分析(one-way ANOVA)和最小差异显著法(LSD)进行比较分析;SPSS 19.0统计软件完成统计分析。
2 结果与分析
2.1 两种人工林豆科植物组成及重要值
表2显示,岷江柏林下物种总数为41种。灌木层植物15种,包括豆科、漆树科、蔷薇科、芸香科、冬青科、忍冬科、小檗科、白花丹科、杜鹃花科、鼠李科,其中豆科植物种类最多,共5种,分别为岷谷木蓝(Indigofera lenticellata)、鞍叶羊蹄甲(Bauhinia brachycarpa)、胡 枝 子(Lespedeza bicolor)、白刺花(Sophora davidii)、锦鸡儿(Caragana sinica),豆科灌木平均重要值达43.99%。草本层植物26种,包括菊科(8种)、禾本科(5种)、蔷薇科(2种)、毛茛科(2种)、豆科(2种)、蓼科(2种)、莎草科(1种)、千屈菜科(1种)、紫葳科(1种)、龙胆科(1种)、玄参科(1种),其中豆科平均重要值为5.91%,主要是绣球小冠花(Securigera varia)、地八角(Astragalus bhotanensis)。随着海拔升高(1 290~1 740 m),豆科灌木重要值呈下降趋势,但岷谷木蓝重要值无明显变化,维持在10%以上;豆科草本逐渐出现且重要值增加。

表 2 两种人工林豆科植物组成及重要值Tab. 2 Species composition and important value of Leguminous plants in two plantations
辐射松林下仅有21个物种,是岷江柏物种总数的50%。灌木层植物6种,包括豆科、蔷薇科、芸香科、萝摩科、卫矛科,其中豆科有胡枝子、白刺花2种,豆科灌木平均重要值为42.57%。草本层植物15种,包括菊科(6种)、禾本科(2种)、蓼科(2种)、堇菜科(1种)、毛茛科(1种)、茄科(1种)、莎草科(1种)、马鞭草科(1种),无豆科植物。随着海拔升高(1 520~1 710 m),豆科灌木重要值逐渐降低,但胡枝子重要值仍然能维持在30%以上。
2.2 两种人工林豆科植物生物量
调查结果显示,干旱河谷豆科灌木植株平均高度:胡枝子>锦鸡儿>白刺花>鞍叶羊蹄甲>岷谷木蓝;平均盖度:鞍叶羊蹄甲、胡枝子、岷谷木蓝>白刺花>锦鸡儿;单株生物量:胡枝子>锦鸡儿>白刺花>岷谷木蓝>鞍叶羊蹄甲。豆科草本植株平均盖度:地八角>绣球小冠花,单株生物量排序为:绣球小冠花>地八角。调查只发现岷谷木蓝和锦鸡儿有根瘤,其中岷谷木蓝根瘤数量相对多一些,其他植物均无根瘤。
岷江柏人工林鞍叶羊蹄甲和岷谷木蓝的平均高度低于1 m,鞍叶羊蹄甲全株生物量低于10 g,岷谷木蓝单株生物量在10~20 g之间;胡枝子、白刺花、锦鸡儿的平均高度大于1m,3种灌木全株生物量均在20 g以上;草本层绣球小冠花、地八角平均高度低于50 cm,全株生物量均在10 g以下。随着海拔升高(1 290~1 740 m),豆科灌木总盖度呈下降趋势,逐渐出现了豆科草本且盖度有所提高;豆科植物总生物量在减少,但岷谷木蓝单株生物量积累增加。此外,豆科植物地下部分生物量占全株生物量的比例为45%,部分豆科植物有根瘤。
辐射松人工林灌木层胡枝子、白刺花平均高度大于1 m,全株生物量均在20 g以上,无豆科草本分布。随着海拔升高(1 520~1 710 m),豆科灌木总盖度变化不大;豆科植物总生物量变化不明显,胡枝子单株生物量总体呈上升趋势。豆科植物地下部分生物量占全株生物量的比例为38%,所有豆科植物均未发现根瘤。
总体来看,岷江柏人工林豆科植物盖度、生物量积累远远高于辐射松人工林,且部分豆科植物有根瘤,地下部分生物量占全株生物量的比例较高。
2.3 两种人工林林下植被生物量及物种多样性
从表4可以看出,岷江柏林下灌木平均盖度30%,生物量2.40 kg;草本盖度43%,生物量1.01 kg;枯落物生物量0.63 kg。随着海拔升高(1 290~1 740 m),灌草植被盖度、灌草生物量以及枯落物生物量逐渐降低。辐射松林下灌木平均盖度12%,生物量0.20 kg;草本盖度15%,生物量0.34 kg;枯落物生物量0.32 kg。随着海拔升高(1 290~1 740 m),灌草植被盖度、灌草生物量以及枯落物生物量呈先增加后减少的趋势,但变化幅度不大。总体而言,岷江柏林下灌草植被盖度、灌草生物量以及枯落物生物量远远高于辐射松,这与豆科植物的表现一致。

表 3 两种人工林林下豆科植物生物量Tab. 3 Biomass of Leguminous species in two plantations
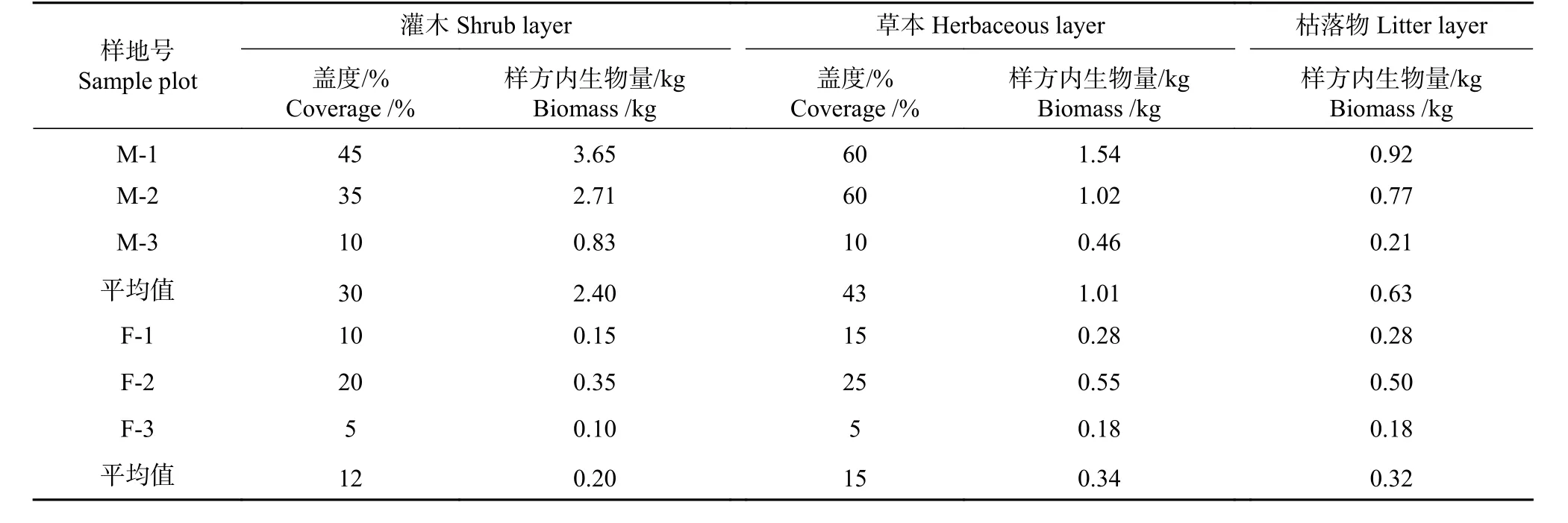
表 4 两种人工林林下植被生物量Tab. 4 Biomass of understory vegetation in two plantations
物种多样性一般采用Margalef丰富度指数、Simpson多样性指数、Shannon-wienner信息指数、Pielou均匀度指数来综合评价。从表5可知,总体来看,岷江柏人工林灌木和草本植被各多样性指数均高于辐射松。其中,岷江柏和辐射松林下灌木的丰富度指数(R)均低于草本,其中岷江柏灌木植被R值约为辐射松的2倍,与上文物种组成的结果一致。而两种人工林林下灌木的均匀度指数(J)均高于草本,表明草本物种相对较多且分布不均匀,结构复杂性和多样性较高[18]。

表 5 两种人工林林下植被物种多样性Tab. 5 Species diversity of understory vegetation in two plantations
3 结论与讨论
豆科植物组成。调查结果显示,岷江柏林下41种灌草植物中,有豆科灌木5种(岷谷木蓝、鞍叶羊蹄甲、胡枝子、白刺花、锦鸡儿)、草本2种(绣球小冠花、地八角),其中岷谷木蓝是较稳定的伴生灌木植物;随着海拔升高,豆科灌木减少,豆科草本逐渐出现。辐射松林下灌草21种,其中豆科灌木2种(胡枝子、白刺花),胡枝子在灌木层优势明显,无豆科草本;随着海拔升高,豆科灌木减少。研究证实,干旱河谷地区主要生长着大量的喜阳、耐旱乡土豆科灌木和草本植物,这与该地区独特的气候类型相符[19];同时,豆科植物也是该区域植被恢复中应用最广、适用性强的物种[3,20]。此外,与豆科草本植物相比,豆科灌木对干旱河谷核心区严酷的环境胁迫适应性更强[19]。相关研究表明,豆科灌木的一级根较细,比根长较大,对土壤资源的获取效率较高,在养分吸收上更具有优势,环境适应性更强,这类灌木对人工林林下灌木层的构建具有重要意义[21-23]。可见,岷江柏人工林豆科植物种类更丰富,对干旱胁迫、外界干扰的抵抗能力相对更强。
豆科植物生物量。随着海拔升高,岷江柏林下豆科灌木盖度下降,豆科植物总生物量在减少。其他学者也得出了一致结果,认为在干旱河谷低海拔地段,干旱、贫瘠环境胁迫更加严重,豆科灌木物种丰富度却较高[19],而随着海拔升高、干旱减轻,其他适应物种逐渐增加,豆科植物盖度、生物量就相应地慢慢降低。然而,岷谷木蓝的地上生物量、地下生物量及总生物量显著增大,这与李芳兰的研究一致[17]。辐射松人工林豆科植被灌木总盖度、豆科植物总生物量变化不大,这可能与海拔垂直高度差不大,干旱环境水分差异不明显有一定关系。与辐射松相比,岷江柏豆科植物地下部分生物量占比较大,分析认为岷江柏人工林郁闭度低、林隙大,水分蒸发大导致干旱程度高,地下生物量分配比例增大,根系更发达;而辐射松人工林郁闭度太高、光照少,在弱光环境中则表现出高生长速率和比叶面积增大,地上生物量积累更多[24]。本次研究发现岷江柏林下的岷谷木蓝和锦鸡儿根系中有少量根瘤,可以发挥固氮作用,在干旱河谷退化土壤改良中具有更好的应用潜力[17]。而其他灌木的根系微生物活性可能受干旱环境影响太大,根瘤形成受到限制,从而丧失了结瘤固氮机制,可见岷谷木蓝对环境胁迫的适应性更强[25]。在干旱河谷植被恢复中,应重视并提倡栽种岷谷木蓝这类具有结瘤固氮特征的植物[26,27]。
林下植被生物量及物种多样性。生长季高温、风频、风急等引起的过量蒸发是导致岷江流域严重干旱的主要原因,有效减少地表蒸发是该地区植被恢复面临的巨大挑战[28]。岷江柏林下灌草植被盖度、生物量以及生物多样性指数均远高于辐射松,灌木生物量能达到辐射松的20倍。分析认为,由于岷江柏植株矮小、冠幅更窄,郁闭度低、林隙更大,地表环境变化更剧烈,各要素在林窗内部的空间异质性越强,从而丰富了灌草物种多样性,促进了生物量积累[29-31],提高了植被覆盖率,进而增强生态系统稳定性[32]。而在降雨过程中,凋落物堆积较多,可以机械阻拦减少地表径流,从而增加土壤的持水性能[33]。随着海拔升高,岷江柏灌草植被盖度、灌草生物量以及枯落物生物量逐渐降低,这与高海拔人工林的林分密度大有很大关系[34]。而辐射松林下植被变化幅度不大,可能因为冠层结构、林分密度相似,所以林下植被的生长、生物量差异不明显[35,36]。此外,林下草本层物种相对较多且分布不均匀,结构复杂性和生物多样性高于灌木层。
对比研究结论。总体而言,岷江柏人工林林下植被盖度大、物种多样性高,且豆科植物种类丰富、生物量积累多,还分布有发挥固氮作用的岷谷木蓝,对干旱胁迫、外界干扰的抵抗能力相对更强。
猜你喜欢
散文诗世界(2021年12期)2021-12-17
科学中国人·上半月(2021年4期)2021-07-20
科学导报(2020年82期)2020-12-23
科学导报(2020年83期)2020-01-13
岷峨诗稿(2019年4期)2019-04-20
阿来研究(2018年1期)2018-07-10
现代园艺(2017年21期)2018-01-03
农业环境科学学报(2017年2期)2017-03-20
廉政瞭望(2016年13期)2016-08-11
天津科技大学学报(2015年3期)2015-04-16
