
GA3和6-BA 对柿砧木种子破眠促萌效果研究*
2022-11-11 12:23:22曹璐瑶于晓丽宋来庆赵玲玲王彦波张青林刘彬徐君驰赵建伟杜晓云
中国果树 2022年11期
曹璐瑶,于晓丽,宋来庆,赵玲玲,王彦波,张青林,刘彬,徐君驰,赵建伟,杜晓云
(1 烟台大学生命科学学院,山东364003)(2 山东省烟台市农业科学研究院)(3 华中农业大学园艺植物生物学教育部重点实验室)(4 黄冈师范学院经济林木种质改良与资源综合利用湖北省重点实验室大别山特色资源开发湖北省协同创新中心)
“三当”育苗是多年生木本果树快速高效育苗的一种方式,即当年播种、当年嫁接、当年出圃,使育苗时间较常规育苗缩短1 年,可减少投入时间和成本,提高经济效益,市场前景良好,然而对育苗技术也提出了更高的要求。冬季提早播种解除种子休眠、促进种子萌发是育苗关键技术环节之一。研究表明,植物生长调节剂处理可有效解除种子休眠促进萌发[1-3],其中以GA3和6-BA 的应用较为广泛,在多种植物上取得了良好的效果[4-9]。
近年来,甜柿产业飞速发展,苗木需求剧增,对高效快速的育苗技术提出迫切需求,“三当”育苗技术体系的建立,将为甜柿苗木产业提供有力保障。但目前对甜柿育苗过程中每个环节的关键技术尚处于探索阶段,文献报道主要集中于田间的农事操作要领[10-11],而对提早播种解除种子休眠和促进萌发的技术尚未见专门研究。孙鹏等[12]、徐长宝等[13]和姬孝忠[14]利用500 mg/L GA3处理种子均取得较好的促萌效果,但仅针对低温层积后春季播种前的种子,是否适用于冬季提早播种尚不得而知。GA3和6-BA 对解除种子休眠和促进萌发有效,但在柿砧木种子上的研究较少,为此,以当前生产上较多使用的君迁子(Diospyros lotusLinn.)和小果甜柿(Diospyros kakiThunb.)2 种柿砧木作为研究对象,开展GA3和6-BA 对2 种柿砧木种子破眠促萌试验,以期为柿苗木高效生产提供技术参考。
1 材料与方法
1.1 试验材料
于2020 年11 月下旬在烟台市农业科学研究院柿圃(北纬37°29′9″,东经121°16′28″)采集君迁子和小果甜柿果实,取种、清洗、阴干,保存于4 ℃冰箱。于2020 年12 月上旬开展种子破眠促萌试验。
1.2 种子基本性状测定
1.2.1 种子形态特征观察
随机选取100 粒种子,种子形状和颜色采用目测法进行观察,参照《柿种质资源描述规范和数据标准》[15]进行描述;采用游标卡尺测量种子长度、宽度、厚度,取其平均值(精度0.01 mm)。
1.2.2 种子千粒重测定
参照《林木种子检验规程》(GB 2772—1999),随机数取8 组种子,每组100 粒,分别称重,计算种子千粒重。
1.2.3 种子含水量测定
参照《林木种子检验规程》(GB 2772—1999),将种子切成小块,均匀放置在样品盒中,置于(103±2)℃的烘箱中进行烘干处理,分别称取烘干前和烘干24 h 后的种子重量。
种子含水量(%)=[(烘干前种子重量-烘干后种子重量)/烘干前种子重量]×100
1.2.4 胚根及幼苗根系结构观察
种子萌发后,留少量种子在培养皿内继续培养,15 d 后记录胚根伸长情况,其余点播于50 孔深穴育苗盘中,置于有供暖设施的日光温室内生长,育苗基质为商道商业化专用育苗基质,日常肥水管理,50 d 后观察幼苗根系结构,拍照记录。
1.3 研究方法
1.3.1 试验设计
使用GA3和6-BA 2 种植物生长调节剂,每种植物生长调节剂设置6 个浓度,GA3浓度分别为0、250、500、750、1 000、2 000 mg/L,6-BA 浓度分别为0、5、10、20、50、100 mg/L。采用二因素六水平正交试验设计,共设36 个处理,每个处理使用种子数量为30 粒,设3 次重复,取平均值。
1.3.2 试验操作
使用0.2%KMnO4溶液进行种子表面消毒,室温浸泡2 h,流动纯水充分清洗,用滤纸吸干种子表面水分。将目的数量的种子放入提前配好的植物生长调节剂溶液中,55 ℃恒温水浴浸泡16 h,纯水冲洗数次。采用培养皿滤纸法,将处理后的种子平铺于具有2 层无菌滤纸、直径为9 cm 的培养皿内,15 mL 纯水润湿,培养皿加盖放置在温度28 ℃、相对湿度75%的光照培养箱内恒温培养,保持滤纸湿润。为防止发霉,定时更换培养皿中纯水。从第2 d 开始统计萌发数量,发芽期限设为30 d。以胚根伸长2 mm 为标准,计算发芽率和发芽势,从发芽试验开始至第1 粒种子萌发所用天数计为发芽时滞天数。
发芽率(%)=(种子萌发总数/供试种子总数)×100
发芽势(%)=(日平均发芽数达到最高那天为止正常发芽的种子总数/供试种子总数)×100
1.4 数据处理
采用Excel 2019 软件处理数据,利用SPSS 25.0软件邓肯氏新复极差法进行多重比较和方差分析。
2 结果与分析
2.1 种子性状及其相关形态特征
君迁子种子红褐色,宽半月形,饱满,种壳坚硬,种皮光滑,横断面长形,长度11.61 mm,宽度5.86 mm,厚度2.70 mm,千粒重146.3 g,含水量21.27%;胚芽宽大,呈阔卵形,胚轴略粗,横断面近倒楔形;幼苗根系发达,易分支,须根繁茂,主根明显;叶先端狭急状,基部宽楔形,整体呈梭形;果实近圆形,果皮橙黄色,果个小,单果种子数7~8 粒。
小果甜柿种子棕褐色,窄半月形,略扁,种壳坚硬,种皮粗糙,横断面长形,长度14.79 mm,宽度8.44 mm,厚度3.90 mm,千粒重318.2 g,含水量13.72%;胚芽狭长,似柳叶形,胚轴细长,横断面近长形;幼苗根系稀疏,须根少,主根粗壮;叶先端阔急状,基部圆形,整体呈阔椭圆形;果实扁圆形,果皮橙红色,果个比君迁子大1 倍之多,单果种子数6~8 粒。
总体看来,君迁子和小果甜柿在种子大小、种胚结构、萌发后的胚根结构、幼苗的叶型、根系结构、果实形态等方面差异明显(图版1)。
2.2 不同植物生长调节剂对2 种柿砧木种子发芽率和发芽势的作用效果
不同植物生长调节剂对君迁子和小果甜柿砧木种子发芽率和发芽势的作用效果不同(表1)。在36 个处理作用下,君迁子砧木种子的发芽率范围为16.67%~83.33%、发芽势范围为8.89%~53.33%,其中处理32 的发芽率和发芽势均最高,处理3 的发芽率和发芽势均最低;小果甜柿砧木种子在36 个处理作用下的发芽率范围为23.33%~54.44%、发芽势范围为12.22%~33.33%,其中处理35 的发芽率和发芽势均最高,处理7、8 的发芽率最低,处理12 的发芽势最低。

表1 君迁子和小果甜柿砧木种子在36 个处理作用下的发芽率和发芽势

续表1
通过进一步计算GA3、6-BA 及其组合对2 种柿砧木种子发芽率和发芽势的变异系数可以得出(表2),GA3和6-BA 组合作用产生的君迁子和小果甜柿砧木种子发芽率和发芽势的变异系数差异较大,君迁子砧木种子发芽率和发芽势的变异系数分别为33.75%和41.35%,小果甜柿砧木种子发芽率和发芽势的变异系数分别为23.89%和27.52%。总体看来,GA3和6-BA 组合对君迁子砧木种子发芽率和发芽势产生的变异系数均高于小果甜柿,说明GA3和6-BA 组合对君迁子砧木种子的破眠促萌作用大于小果甜柿。对单独使用GA3和6-BA 对2 种柿砧木种子发芽率和发芽势的影响程度进行分析可以得出,6-BA 对2 种柿砧木种子发芽率和发芽势的变异系数为1.78%~12.82%,平均值为6.13%,其中6-BA 对小果甜柿砧木种子发芽率的影响最弱;而GA3对2 种柿砧木种子的发芽率和发芽势的变异系数均明显大于6-BA,GA3对2 种柿砧木种子发芽率和发芽势的变异系数为24.31%~33.81%,平均值为29.02%。可见,GA3对君迁子和小果甜柿砧木种子的破眠促萌作用大于6-BA。

表2 2 种植物生长调节剂及其组合对2 种柿砧木种子作用数据的分组统计 %
发芽时滞天数越短意味着响应速度可能越快,由图1 可知,36 个处理作用于君迁子砧木种子的发芽时滞天数低于小果甜柿,且在0.01水平差异显著,表明君迁子砧木种子对植物生长调节剂的作用可能更加敏感;君迁子砧木种子最高发芽率天数也低于小果甜柿,且在0.001 水平差异显著;君迁子砧木种子的发芽率在0.01 水平显著高于小果甜柿;君迁子砧木种子的发芽势与小果甜柿无显著差异。总体看来,在发芽率、发芽势、发芽时滞天数、最高发芽率天数4 个指标上,君迁子和小果甜柿均显示出明显不同的数据分布样式,表明2 种柿砧木种子对2 种植物生长调节剂的作用反应不同,君迁子砧木种子对2 种植物生长调节剂的作用可能更加敏感。

图1 2 种植物生长调节剂对2 种柿砧木种子发芽率、发芽势、发芽时滞天数、最高发芽率天数的数据分布及差异显著性
2.3 不同植物生长调节剂浓度对君迁子砧木种子破眠促萌的影响
如图2 所示,在GA3浓度为0~750 mg/L 范围内,君迁子砧木种子发芽率均值和发芽势均值的变化差异均不显著;在GA3浓度为750 mg/L 时,君迁子砧木种子发芽率均值最高,为39.26%;在GA3浓度为0 mg/L 时发芽率均值最低,为32.41%;GA3浓度为750 mg/L 和500 mg/L 时分别产生最高发芽势均值(19.82%)和最低发芽势均值(16.29%)。当GA3浓度提高到1 000 mg/L 时,与前4 种浓度相比,君迁子砧木种子发芽率均值和发芽势均值均显著提高,由36.48%(0~750 mg/L 发芽率均值)和17.92%(0~750 mg/L 发芽势均值)提高到52.78%(1 000 mg/L 发芽率均值)和28.52%(1 000 mg/L发芽势均值),分别提高了16.30、10.60 个百分点;当GA3浓度为2 000 mg/L 时,君迁子砧木种子发芽率均值和发芽势均值均达到最高值,与1 000 mg/L浓度相比,发芽率均值显著提高了17.78 个百分点,而发芽势均值的变化差异不显著。

图2 不同植物生长调节剂浓度对君迁子砧木种子破眠促萌的影响
不同6-BA 浓度处理下的君迁子砧木种子发芽率均值为42.78%~46.67%,发芽势均值为19.63%~27.04%,发芽率均值和发芽势均值在不同6-BA 浓度处理下均无显著差异(图2)。
总的看来,以6 个GA3处理浓度对君迁子砧木种子破眠促萌的效果进行分组,发芽率水平可以归为3 个组:0~750 mg/L、1 000 mg/L、2 000 mg/L作用组;发芽势水平可以归为2 个组:0~750 mg/L、1 000~2 000 mg/L 作用组;6-BA 处理浓度对君迁子砧木种子破眠促萌作用不明显。以GA3、6-BA 及其组合作用为自变量,君迁子砧木种子发芽率和发芽势为因变量进行多因素方差分析,结果显示,GA3处理对君迁子砧木种子的发芽率和发芽势均产生极显著影响,而6-BA 处理影响不显著。
2.4 不同植物生长调节剂浓度对小果甜柿砧木种子破眠促萌的影响
如图3 所示,小果甜柿砧木种子在GA3浓度为0~750 mg/L 范围内的发芽率均值为28.70%~31.85%,且各浓度处理间差异不显著;GA3浓度为1 000 mg/L 和2 000 mg/L 时,发芽率均值分别为40.37%和49.82%,与0~750 mg/L 浓度相比均存在显著性差异。小果甜柿砧木种子在GA3浓度为0~750 mg/L 范围内的发芽势均值为15.93%~16.30%,各浓度处理间无显著差异;在GA3浓度为1 000 mg/L时,发芽势均值提高到23.15%;在GA3浓度为2 000 mg/L 时,发芽势均值达到最高值,为27.59%。发芽率和发芽势在总体变化趋势上基本一致。
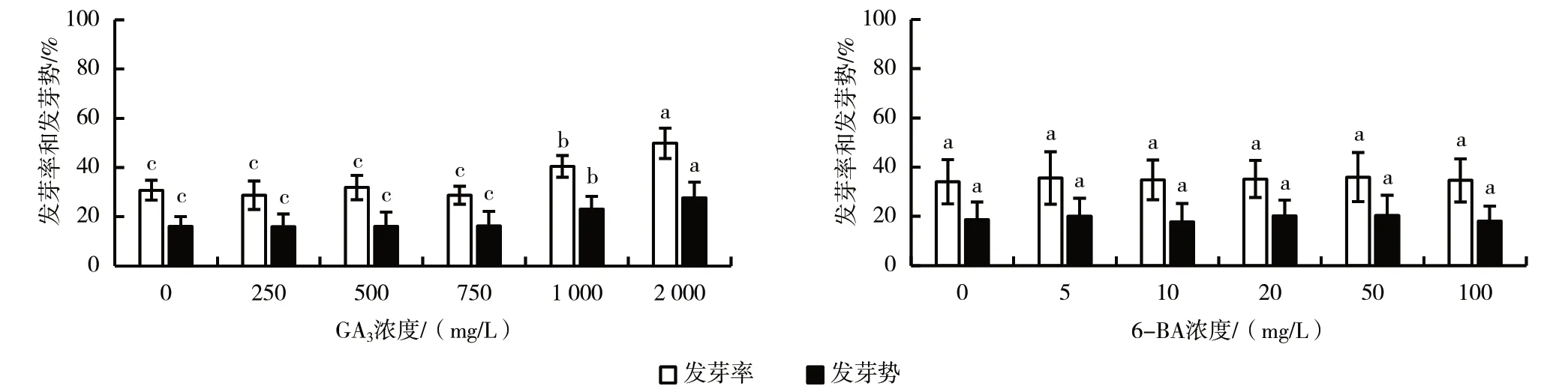
图3 不同植物生长调节剂浓度对小果甜柿砧木种子破眠促萌的影响
不同6-BA 浓度处理对小果甜柿砧木种子发芽率均值和发芽势均值均无显著性差异,发芽率均值为34.07%~35.93%,平均值为35.03%;发芽势均值为17.78%~20.37%,平均值为19.20%(图3)。
总的看来,以6 个GA3处理浓度对小果甜柿砧木种子破眠促萌的效果进行分组,发芽率和发芽势水平均可以归为3 个组:0~750 mg/L、1 000 mg/L、2 000 mg/L 作用组;6-BA 处理浓度对小果甜柿砧木种子破眠促萌作用不明显。以GA3、6-BA 及其组合作用为自变量,小果甜柿砧木种子发芽率和发芽势为因变量进行多因素方差分析,结果显示,GA3处理对小果甜柿砧木种子的发芽率和发芽势的影响均达到极显著水平,而6-BA 处理影响不显著。
3 讨论
种子休眠是植物在长期进化过程中形成的抵御外界恶劣环境的自我保护机制[16],种子在非适宜季节萌发需要采取一定措施打破休眠。使用植物生长调节剂是打破种子休眠的重要手段之一,其中GA3和6-BA 是较常用的2 种植物生长调节剂[17-18]。现有研究认为,赤霉素通过影响激素间的平衡,对打破种子休眠促进萌发具有一定作用[17]。本研究通过不同浓度的GA3处理,在2 种柿砧木种子上均获得萌发率显著提高的效果,表明GA3对打破柿种子休眠促进萌发具有明显效果,与前人结论一致[4-6]。对于6-BA 的作用效果存在分歧,较多结论认为其具有促进种子萌发的作用[7-9],但也有研究发现其作用效果并不明显[19-20]。本研究通过计算不同浓度植物生长调节剂及其组合处理对2 种柿砧木种子发芽率和发芽势的变异系数、主体间效应检验以及多因素方差分析,研究结果均表明6-BA 对君迁子和小果甜柿砧木种子破眠促萌的效果并不明显。该结果说明,GA3对打破柿种子休眠至关重要,外施GA3可以在一定程度上解除种子休眠,而6-BA 可能并非决定其破眠特性的关键影响因素,具体原因尚需进一步深入研究。
植物生长调节剂对种子破眠促萌效果具有剂量效应,在适宜浓度下才能起到促进作用[21-22],已有研究表明,GA3适宜浓度为1 000 mg/L 以下,6-BA适宜浓度为50 mg/L 以下。本研究结果表明,GA3浓度在1 000 mg/L 时对2 种柿砧木种子均开始发挥破眠促萌作用,其后浓度再增加,发芽率和发芽势也得到明显提高,在所设最大浓度为2 000 mg/L 时还未到拐点;而2 种柿砧木种子在各种6-BA 浓度处理下的效果均不明显。植物生长调节剂对种子破眠促萌效果因植物种类、种子形态、休眠程度等不同而存在很大差别[23-25],而本研究植物生长调节剂对种子萌发效应与其他文献报道中的区别可能与物种差异有关。姬孝忠[14]、徐长宝等[13]、孙鹏等[12]针对植物生长调节剂对柿种子萌发效应进行研究认为,GA3对种子的萌发具有显著影响,支持本研究的结论,但其研究结果表明,GA3最佳使用浓度为500 mg/L,与本研究结果不一致。500 mg/L GA3浓度对本研究当季冬播的君迁子和小果甜柿砧木种子的萌发基本不起作用,原因可能与研究对象的休眠程度、浸种时间、植物生长调节剂种类等因素有关。后续研究可以从GA3单一处理并增加GA3处理浓度,也可尝试使用其他植物生长调节剂种类进行组合处理,以及其他辅助的物理或化学处理,以进一步获得更理想的萌发率。
君迁子和小果甜柿是柿常用的2 种砧木,分属于不同种,君迁子为我国习用砧木,适合于中国传统柿品种,与近年兴起的日本甜柿亲和性较差;而最近发现的小果甜柿与日本甜柿亲和性良好。因此,本研究选用君迁子和小果甜柿砧木作为研究对象,以满足产业上育苗的多元需求。生产上发现,君迁子根系发达,容易分支,细根多,移栽易成活,缓苗也快;而柿砧木种子普遍发芽率低,主根发达,分支少,根系深,移栽后成活较难[26]。本研究根据2 种柿砧木种子的形态数据、萌发后胚根上主侧根分化状态、幼苗期地上部和地下部形态的观察结果均吻合成年后的形态差异,充分表明2 种柿砧木的遗传差异很大。试验结果所得植物生长调节剂对君迁子和小果甜柿砧木种子破眠促萌迥异的作用效果,可以用植物生长调节剂对差异较大的基因型间差别作用来解释,也间接反映了君迁子和小果甜柿砧木种子的休眠机制可能不同。
种子休眠的原因除与体内激素有关外,还受种皮、种胚发育成熟度、各种内含物等多种因素的影响,提前对具有坚硬种皮的种子采取划破、干燥、提前热层积、低温层积等方法,对种子生理休眠的解除具有一定的促进作用。本研究发现,小果甜柿砧木种子的促早萌芽方法有待进一步提高,因此,今后试验可以考虑在植物生长调节剂作用的同时,再辅以一定的前处理,以寻求对小果甜柿砧木种子更高效的破眠促萌方法。
4 结论
综上可知,一定浓度的GA3对君迁子和小果甜柿砧木种子的休眠解除及促进萌发具有显著的促进作用,而6-BA 在各浓度水平之间无显著影响,也无和GA3的叠加效应;君迁子和小果甜柿砧木种子对2 种植物生长调节剂的反应不同,前者相对而言更加敏感;在本试验设定条件下,GA3浓度≤750 mg/L 时对柿砧木种子破眠促萌的作用效果不明显,而1 000 mg/L 是GA3发挥作用的起始浓度,2 000 mg/L 为最佳浓度,但由于相关种子萌发试验普遍认为GA3的最佳浓度低于1 000 mg/L,因此推测GA3浓度在高于2 000 mg/L 时可能对种子的破眠促萌作用呈下降趋势,而实际情况还有待进行相关的补充试验。
猜你喜欢
作文周刊·小学二年级版(2023年48期)2024-01-01 21:25:24
农业科技通讯(2023年1期)2023-02-12 07:09:08
现代农业科技(2022年2期)2022-12-17 01:36:52
花火B(2021年3期)2021-06-25 02:38:31
现代园艺(2020年19期)2020-10-02 03:15:48
中国果树(2020年2期)2020-07-25 02:14:22
种子(2019年6期)2019-07-13 07:45:16
种子(2019年4期)2019-05-28 02:04:14
现代农村科技(2019年4期)2019-01-06 14:13:19
上海农业学报(2017年3期)2017-04-10 12:39:22
