
Oscillating field stimulation promotes axon regeneration and locomotor recovery after spinal cord injury
2022-11-11 13:46YiXinWangJinZhuBaiZhenLyuGuangHaoZhangXiaoLinHuo
中国神经再生研究(英文版) 2022年6期
Yi-Xin Wang, Jin-Zhu Bai,, Zhen Lyu, Guang-Hao Zhang, Xiao-Lin Huo
Abstract Oscillating field stimulation (OFS) is a potential method for treating spinal cord injury. Although it has been used in spinal cord injury (SCI) therapy in basic and clinical studies, its underlying mechanism and the correlation between its duration and nerve injury repair remain poorly understood. In this study, we established rat models of spinal cord contusion at T10 and then administered 12 weeks of OFS. The results revealed that effectively promotes the recovery of motor function required continuous OFS for more than 6 weeks. The underlying mechanism may be related to the effects of OFS on promoting axon regeneration, inhibiting astrocyte proliferation, and improving the linear arrangement of astrocytes. This study was approved by the Animal Experiments and Experimental Animal Welfare Committee of Capital Medical University (supplemental approval No. AEEI-2021-204) on July 26, 2021.
Key Words: astrocyte orientation; astrocyte proliferation; axonal regeneration; locomotor recovery; neural regeneration; neural repair; oscillating field stimulation; spinal cord injury; stimulus duration
Introduction
Treatment of spinal cord injury (SCI) that promotes nerve regeneration remains an uncertain clinical goal. A previous study has found that axonal regeneration is associated with externally applied electric fields, with newer fibers growing along the long axis of the voltage gradient (Jaffe and Poo, 1979). In another study, neurites preferentially grew towards the cathode and were resorbed from the anode in an applied electric field (Borgens et al., 1993). Growth along the voltage gradient axis is three times faster towards the cathode than towards the anode. This characteristic has led to the development of a treatment for SCI called oscillating field stimulation (OFS).
OFS imposes a weak voltage gradient across the lesion site and reverses its polarity every 15 minutes to promote the growth of nerve fibers in both directions (Borgens et al., 1999). Although OFS has the potential to be a treatment for SCI (Shapiro, 2014; Li, 2019), the underlying mechanism is not well illustrated. Previous studies indicated that OFS can promote remyelination (Zhang et al., 2014) and attenuate secondary apoptotic responses within 4 weeks in rats with SCI (Zhang et al., 2015a). Although axonal regeneration is an important outcome of OFS, the temporal characteristic OFS-induced action has not been studied in detail. The aim of this study was to investigate the effect of OFS on axonal regeneration, astrocyte proliferation, and astrocyte reorientation after SCI, as well as the relationship between these effects and stimulation duration.
Materials and Methods
Animals and grouping
Healthy 6-month-old adult clean-grade Sprague-Dawley rats, weighing 220 ± 10 g were provided by the Experimental Animal Center of the Chinese Academy of Military Medical Sciences (Beijing, China). This study was approved by the Animal Experiments and Experimental Animal Welfare Committee of Capital Medical University (approval No. AEEI-2021-204). All surgical procedures were conducted according to the guidelines of the National Guidelines for Experimental Animal Welfare. The animal feeding and experimental surgeries were completed in the Animal Laboratory of the China Rehabilitation Research Center. Pentobarbital sodium (1%, Chinese Institute of Rehabilitation Science) was injected intraperitoneally at 30 mg/kg of body weight for anesthesia.The rats had free access to food and water throughout the study. The rats (n= 120) were randomly divided into OFS + SCI and SCI groups (n= 60 per group). The OFS + SCI group received the OFS intervention, while the SCI group received sham stimulation (Figure 1A).
Surgical procedures
Rat models of spinal cord contusion were made according to the modified Allen method at T10 (Perot et al., 1987).After anesthesia and being fixed on the NYU impactor (New York University, USA), an SCI model was created using 50 g-cm of potential energy. After spinal cord impingement, the stimulator electrodes were implanted rostral and caudal to the injury site by suturing one to the bilateral facet capsule and the other to the interspinous ligament (Figure 1BandC). The OFS + SCI rats were implanted with a working stimulator and the SCI rats received a sham stimulator. The positions of the electrodes were confirmed by X-ray.
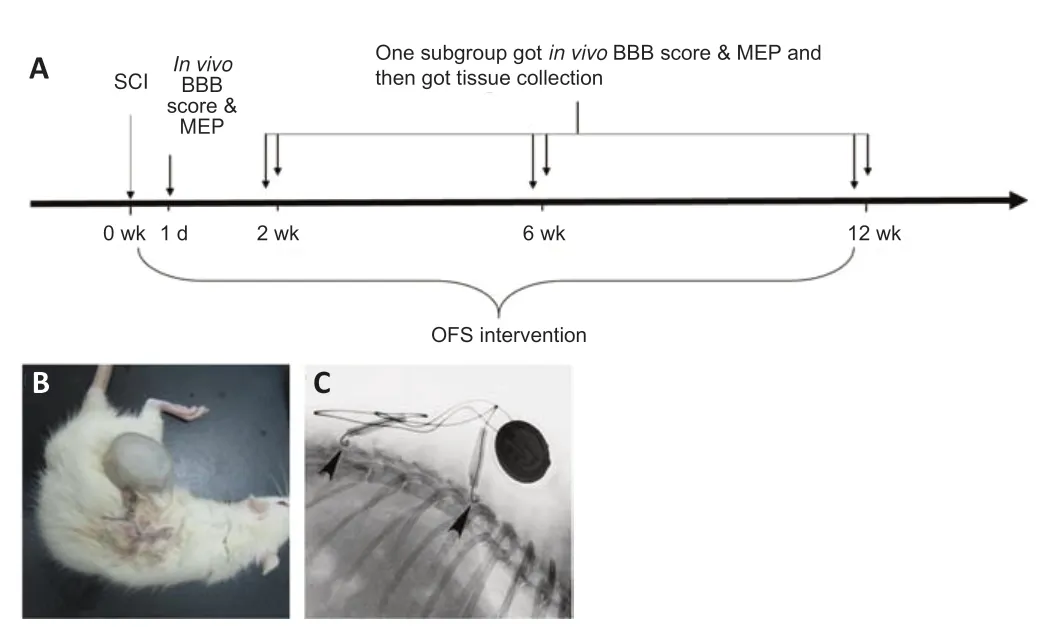
Figure 1|Experimental procedure and the OFS implantation.
Stimulator fabrication
The oscillating field stimulator was designed by the School of Electronics, Electrical and Communication Engineering, University of Chinese Academy of Sciences. The stimulator uses a 3.3 V battery (CR2032, 210 mA, Panasonic, Jakarta, Indonesia) as the power unit. An oscillating voltage with an interval of 15 minutes is generated by a binary ripple counter (74HC4060, Texas Instruments, Dallas, TX, USA) with a 47 nF capacitor and two resistors (910 k and 6.2 M). The oscillating voltage is converted to three 12-μA oscillating currents through three constant current circuits. Using Pt-Ir electrodes, the stimulating currents are delivered into the biological tissue after the oscillating field stimulator is implanted into an animal’s body. Oscillating electric field stimulation (electric field intensity, 500 V/mm; polarity alternated every 15 minutes) began immediately after surgery and the stimulation was continued uninterrupted for 12 weeks. Sham stimulators were fashioned in exactly the same way, but current was blocked by attaching insulated rubber tape to the battery.
Behavioral assessment
Basso-Beattie-Bresnahan (BBB) scores (Basso et al., 1995) were used to evaluate locomotor function in the rats. The maximum score is 21 points, with lower scores indicating worse motor function. BBB scores were assessed on Day 1, and on Weeks 2, 6, and 12 after SCI (n= 20 per assessment). To decrease errors, the assessments were independently performed by two of the authors (YXW and ZL) who were blinded to rat grouping.
Motor evoked potential
At 2, 6 and 12 weeks after SCI (n= 20 per group), motor evoked potentials (MEPs) were evaluated by neuroelectrophysiological stimulation (Medelec Synergy, Berlin, Germany). Two pairs of stimulating needle electrodes were separately placed on the spinous process intervals: on the rostral side at T7/8 and on the caudal side at L1/2. Negative electrodes were put at the caudal side. The distance between the two electrodes was 0.5-0.8 cm. The stimulation was a 0.05-ms wave-width rectangular pulse with an intensity of 100-150 V. A pair of recording needle electrodes was placed bilaterally in the gastrocnemius. MEPs were the compound muscle action potentials (CAMPs) recorded in the target muscle after stimulation of the spinal cord. The latency and amplitude of the CAMPs were recorded. The differences in CAMP latencies and amplitudes between the rostral and caudal sides were evaluated. Smaller differences indicate better functional recovery (Tian et al., 2016).
Hematoxylin-eosin staining
Hematoxylin-eosin staining was performed (n= 20 per group) at 2, 6, and 12 weeks. After rats were perfused with 4% paraformaldehyde and fixed, the injured spinal cord segments, along with 2 cm of adjacent tissue, were harvested and fixed in the 4% paraformaldehyde for 12 hours. Paraffin-embedded tissue sections (10 μm thick) were sliced consecutively. After dewaxing and rehydration, sections were stained with hematoxylin, differentiated with 1% hydrochloric acid, and stained with eosin. Tissue integrity, including defects and cystic changes, was observed under a light microscope (Olympus Corporation, Tokyo, Japan).
Immunohistochemical and immunofluorescence staining
Immunohistochemistry and immunofluorescence staining at 2, 6, and 12 weeks after SCI (n= 20 per group) were used to determine the extent of axon regeneration and astrocyte proliferation in the injured area. The injured spinal cord samples first underwent antigen retrieval and block. The samples were incubated with mouse anti-rat NF200 monoclonal antibody (1:1000, Sigma, St. Louis, MO, USA, Cat# WH0004744M1, RRID No. AB_1842647) or rabbit anti-rat glial fiber-acid protein (GFAP) polyclonal antibody (1:500, Sigma, Cat# G4546, RRID No. AB_1840895) at 37°C for 1 hour. They were then rinsed with phosphate buffered saline (PBS) three times for 2 minutes each. For NF200 immunohistochemical staining, the samples were incubated with goat anti-mouse IgG (1:200, Sigma, Cat# SAB4600066, RRID No. AB_2336060) for 1 hour at 37°C. The samples were then rinsed three times for 5 minutes, stained with diaminobenzidine for 5-10 minutes, and counterstained with hematoxylin for 2 minutes. After differentiation in acid-alcohol solution, dehydration, rinsing, and mounting, the samples were observed with a light microscope. For GFAP immunofluorescence staining, the samples were rinsed and incubated with Alexa Fluor 568 conjugated goat anti-rabbit IgG (1:200; Zymed, San Diego, SC, USA, Cat# A-11036, RRID No. AB_10563566) for 2 hours at room temperature. After rinsing and mounting, the samples were observed by fluorescence microscope (DMLA 4000B, Leica, Solms, Germany). GFAP integrated optical density (IOD) and the angle of the astrocyte processes (0-90°, with smaller angles indicating a more linear cell orientation) at the injury site were measured using Image-Pro Plus 6.0 software (Media Cybernetics, Rockville, MD, USA).
Statistical analysis
Statistical analysis was performed with SPSS 18.0 software (SPSS Inc. Chicago, IL, USA). All data are expressed as the mean ± standard deviation (SD). Two-way (group × time) completely random effects analyses of variance (ANOVAs) were used to investigate the differences in BBB scores, MEP latency, and MEP amplitude between groups (OFS/sham) and over time (day 1, weeks 2, 6, or 12).Post hoctests were performed using the least significant difference test. Simpleeffect analyses were performed to compare each covariance between the two groups. Independent samplest-tests were performed to compare the numbers of regenerated axons at the injury site, IOD values, and astrocyte process angles across time points. APvalue < 0.05 was considered statistically significant.
Results
OFS improves the locomotor function of bilateral hind limbs in SCI rats
After OFS, BBB scores increased gradually in the OFS + SCI group and were higher than those in the SCI group. The twoway ANOVA revealed significant main effects of time (F(3,312)= 148.516,P< 0.001) and group (F(1,312)= 16.477,P< 0.001) on BBB score, as well as a significant group × time interaction (F(3,312)= 12.583,P< 0.001). The least significant difference post hoc test showed that the BBB score increased over time. The simple effects analysis showed statistically significant differences in BBB score at each time point for the OFS + SCI group (P< 0.001). BBB score was the highest at 12 weeks. In the SCI group, BBB score increased gradually, and differed significantly between day 1 and week 2 (P< 0.001), and between weeks 2 and 6 (P< 0.01) but did not differ between weeks 6 and 12 (P= 0.675). Additionally, although BBB score did not differ significantly between the two groups at 1 day or at 2 weeks (day 1:P= 0.648; week 2:P= 0.209), scores were significantly higher in the OFS + SCI group than in the SCI group at 6 and 12 weeks (week 6:P< 0.01, week 12:P< 0.001;Figure 2A).
OFS improves the MEP of bilateral hind limbs in SCI rats
As with BBB score, two-way ANOVAs were used to assess group and time differences in the latencies and amplitudes of evoked MEPs in the electrophysiological experiment. first, we found main effects of time (F(3,312)= 70.443,P< 0.001) and group (F(1,312)= 20.101,P< 0.001) on MEP latency, as well as a significant group × time interaction (F(3,312)= 6.428,P< 0.001). Post hoc analysis showed that latencies differed significantly between day 1 and week 2 (P< 0.001), between weeks 2 and 6 (P< 0.001), and between weeks 6 and 12 (P< 0.05;Figure 2B). The simple effects analysis showed that latencies for the OFS + SCI group differed significantly between different time points (day 1vs. week 2:P< 0.001; week 2vs. week 6:P< 0.001; week 6vs. week 12:P< 0.05). Conversely, latencies for the SCI group only differed significantly between day 1 and week 2 (P< 0.001) (week 2vs. week 6:P= 0.868; week 2vs. week 12:P= 0.615; week 6vs.week 12:P= 0.999). Significant differences in latency were observed between the two groups at 6 weeks (P < 0.01) and at 12 weeks (P< 0.001;Figure 2B).
As with latency, we found significant main effects of time (F(3,312)= 79.692,P< 0.001) and group (F(1,312)= 14.928,P< 0.001) on MEP amplitude, as well as a significant time × group interaction (F(3,312)= 9.606,P< 0.001). The least significant difference test showed that amplitudes differed significantly across time points (P< 0.001). The simple effects analysis showed that MEP amplitudes in the OFS + SCI group differed significantly different at certain time points (day 1vs. week 2:P< 0.001; week 2vs. week 6:P< 0.01; week 6vs. week 12:P< 0.001). In the SCI group, statistically significant differences were observed between day 1 and week 2 (P< 0.01), and between weeks 2 and 12 (P< 0.01) but not between weeks 2 and 6 (P= 0.160) or between weeks 6 and 12 (P= 0.696). No differences in amplitude were observed between the two groups at 1 day (P= 0.602), 2 weeks (P= 0.943), or 6 weeks (P= 0.055). However, differences were statistically significant between the two groups at 12 weeks (P< 0.001;Figure 2C).
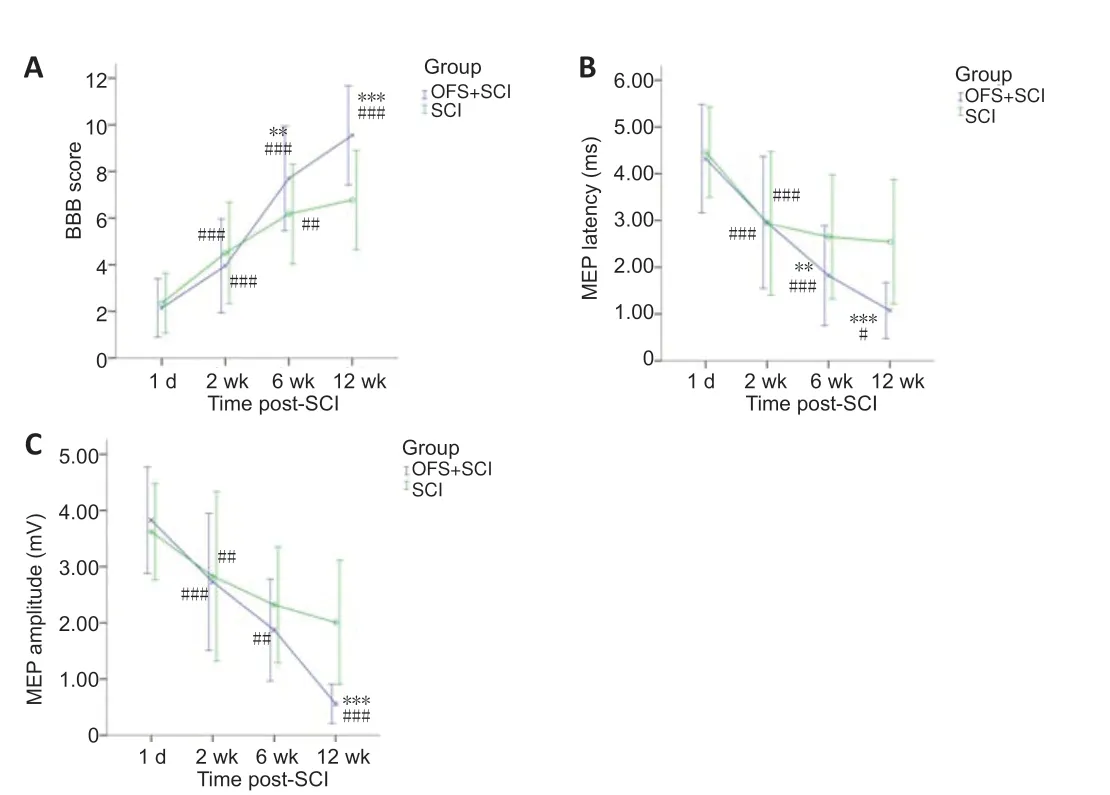
Figure 2|Effect of OFS on locomotor function (BBB score) and neural conduction (MEP).
OFS improves the histological morphology of injured spinal cords in SCI rats Histological morphology of hematoxylin-eosin staining
No significant differences were observed between the two groups under hematoxylin-eosin staining. The border between white and gray matter was unclear at 2 weeks (Figure 3AandB). A tissue defect was observed surrounded by a small amount of white matter at 6 weeks (Figure 3CandD). A glial scar was found at the site of injury at 12 weeks (Figure 3EandF).
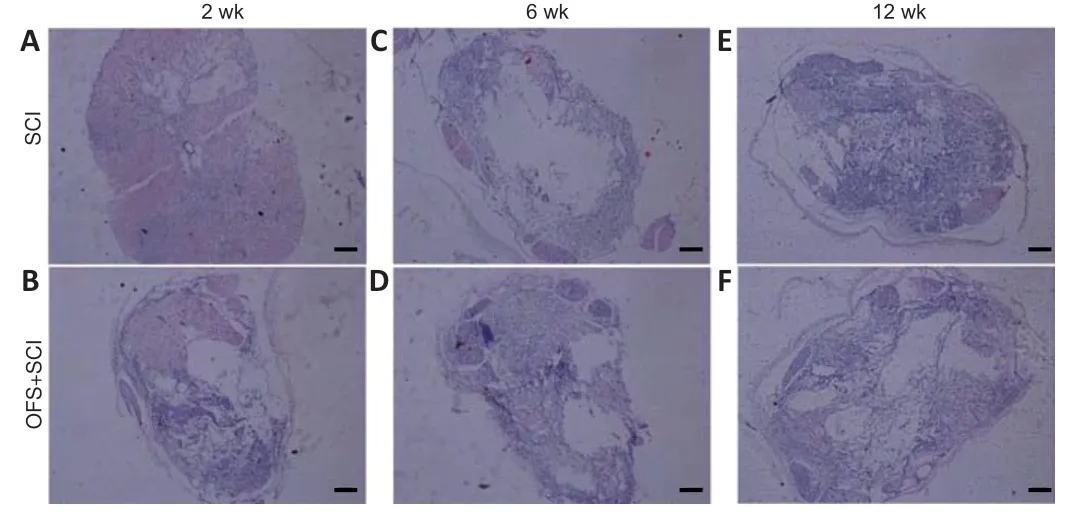
Figure 3|Effect of OFS on the histological morphology of injured spinal cords in SCI rats (hematoxylin and eosin staining).
NF200 immunohistochemical staining and axon counting
NF200-positive nerve fibers in the white matter were observed in both groups at different time points. The NF200 positive nerve-fiber density was higher in the OFS + SCI group than in the SCI group at 12 weeks (Figure 4AandB). In the independent samplest-test, the number of axons did not differ significantly at 2 weeks (P= 0.206) or 6 weeks (P= 0.278). However, at 12 weeks, axon count was significantly higher in the OFS + SCI group than in the SCI group (P< 0.001;Figure 4C).
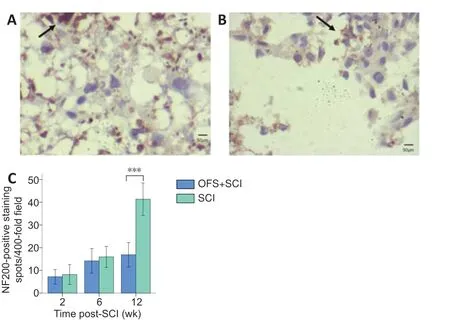
Figure 4|Effect of OFS on NF200-positive nerve fibers in the injured spinal cord of SCI rats (immunohistochemical staining).
GFAP immunofluorescence staining
Using immunofluorescence staining, we determined the IOD value for GFAP (Figure 5AandB). An independent samplest-test showed that IOD did not differ significant at 2 weeks (P= 0.736) or at 6 weeks (P= 0.067). However, at 12 weeks, IOD was significantly lower in the OFS + SCI group than in the SCI group (P< 0.05), which indicates less astrocyte proliferation (Moriarty and Borgens, 2001) (Figure 5C).
Astrocyte process angle
We measured the angles of the astrocyte processes on the GFAP immunofluorescence-stained photos and compared them to evaluate the level of linear orientation in the astrocytes. An independent samplest-test showed that the angles in the OFS + SCI group were smaller than those in the SCI group at 12 weeks (P< 0.001;Figure 5D), which indicates more linear cell orientation.
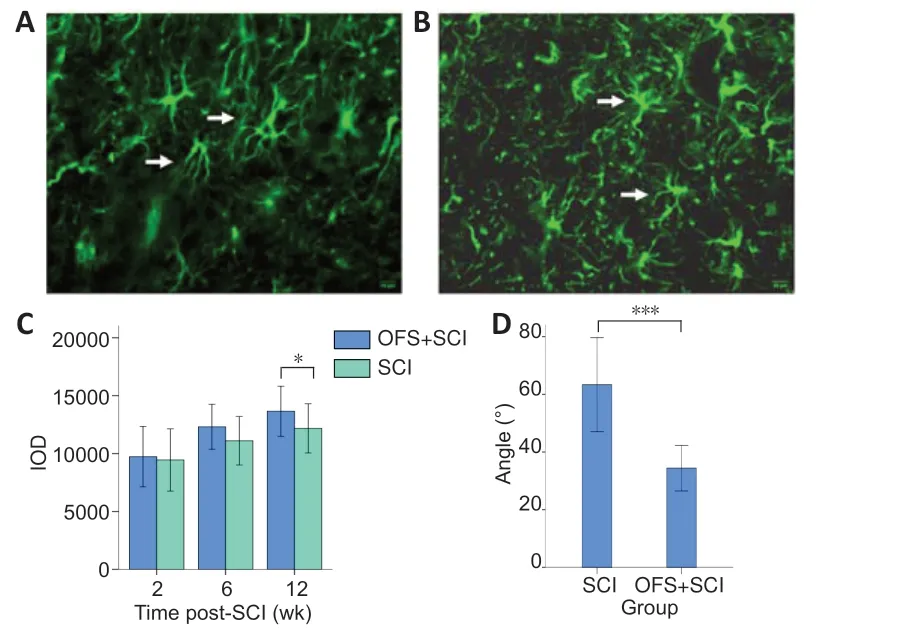
Figure 5|Effect of OFS on GFAP-positive astrocytes in injured spinal cords of SCI rats.
Discussion
Peripheral axons react poorly following SCI. Although some research has shown that many axons attempt regeneration within 6 to 24 hours after injury, they often grow in the wrong directions (Kerschensteiner et al., 2005). Studies have reported that a weak direct current electric field can induce directional nerve growth (Jaffe and Poo, 1979; Borgens et al., 1993). An oscillating field imposes stimulation across a lesion site and because it reverses its polarity every 15 minutes, it can promote the growth of nerve fibers in both directions (Borgens et al., 1999; Huang et al., 2016). There is evidence that OFS can promote neural repair and motor function recovery in SCI (Shapiro, 2014; Jing et al., 2015; Zhang et al., 2015a; Tian et al., 2016; Bacova et al., 2019; Li, 2019). Therefore, OFS has a therapeutic potential in basic and clinical SCI research. The mechanism though which OFS facilitates the repair of spinal cord injury is not yet clear at the cellular level. A previous study indicated that OFS can promote locomotor recovery and remyelination in rats with SCI, which might have been related to the improved differentiation of oligodendrocyte precursor cells in the spinal cord (Zhang et al., 2014). Another study showed that early application of electric field stimulation attenuates secondary apoptotic responses and exerts neuroprotective effects within 4 weeks in rats with acute SCI (Zhang et al., 2015b). Axonal regeneration is the most important facet of neural repair after SCI. However, how OFS influences axon growth and astrocyte orientation and what the most effective stimulation duration still need to be explored.
In the current study, both groups had incomplete SCI. The OFS + SCI group received the OFS intervention immediately. The BBB score and the MEP showed that OFS could promote motor function recovery. When we consider the significant interaction between group and time, we can see that the speed of nerve function recovery was greater in the group that received the OFS intervention. We first observed a difference between the two groups at 6 weeks. Similarly, MEP analysis showed that the motor conduction function of rats with SCI improved to a certain degree after 6 weeks of continuous OFS. Simple effects analysis showed that when only time was considered, motor function in the SCI group spontaneously recovered without intervention at an early stage, and then stagnated over time. This result is consistent with a previous study (You et al., 2003). Conversely, motor function and MEP of the hind limbs improved gradually at each time point in the OFS + SCI group. These findings indicate that OFS needed a certain amount of time to take effect, but once it did, it persisted beyond 6 weeks. The effect of OFS on motor function recovery might be related to improved spinal cord conduction.
A previous study induced regeneration by applying an electric field after axon transection of the dorsolateral spinal cord in adult pigs (Borgens and Bohnert, 1997). New axons traversing the glial scar were usually less than 1 μm and were mainly distributed in the white matter surrounding the injury site, affected by normal axons, and were difficult to identify in the axial immunohistochemical section. In the current study, analysis of the number of axons showed that axon regeneration after 2 weeks remained insufficient to generate statistically significant differences with control rats. At the same time, we did not observe any obvious functional recovery. The short OFS duration and the lack of axon regeneration can explain the low functional recovery. After 6 weeks, even though we saw functional recovery in the intervention group, we still did not see any differences in axon number between groups. This indicates that better motor function recovery in the SCI + OFS group might be related to the early neuroprotective effects of OFS rather than its effects on axonal regeneration. After 12 weeks, we observed more axons in the OFS + SCI group than in the SCI group. Additionally, the number of axons was significantly greater after 12 weeks than after 6 weeks. This indicates that prolonged OFS might be one way to promote axon regeneration and the associated subsequent locomotor recovery after 6 weeks.
Glial scar formation is a primary factor that inhibits axon regeneration and neural functional recovery (Gao et al., 2021; Wang et al., 2021). A previous study found that an electrical field can reduce glial proliferation at the site of injury (Hamid and Hayek, 2008). A higher IOD value for GFAP indicates a more serious glial scar. In the current study, IOD value was lower in the OFS + SCI group than in the SCI group after 12 weeks. Thus, OFS inhibited glial proliferation, and this effect took approximately 12 weeks.
Some cystic cavities form and are surrounded by thick astrocytic scars after SCI. Subsequently, lesions evolved into multilobular cystic structures whose walls were formed by glia (Yokota et al., 2017). Axons cannot grow across cavities, and the scar imposes a physical and chemical barrier to regenerating axons. Several studies have analyzed the directionality of axonal growth in rats with SCI by measuring the angle of axon segments in cellular grafts or implanted biomaterials to evaluate the efficacy of various techniques (Francis et al., 2013; Tuft et al., 2014). A previous study used OFS to suppress astrocyte processes extending at the area of injury and to guide its reorientation along the electric field direction (Moriarty and Borgens, 2001). In the current study, the astrocytic process angles were measured to evaluate the linearity of cell orientation. After 12 weeks, the angles for the OFS + SCI group were significantly lower than those of the SCI group, which indicates greater linearity. Thus, OFS might facilitate axon regeneration by providing space for axon growth. Reduced interweaving of astrocytes might help inhibit glial scar formation and facilitate regenerated axons to cross the scar in the injury site.
This study had some limitations. Because only the 12-week OFS and control groups were included, we cannot be sure what the optimal OFS duration is. It could be that the effects observed at 12 weeks would have been obtained even if the duration of OFS had been stopped earlier. In future studies, we will include two additional groups that receive 2 weeks and 6 weeks of OFS but are assessed at 2, 6, and 12 weeks respectively. This will allow us to determine whether 12 continuous weeks of OFS is necessary for recovery.
In conclusion, our results showed that OFS continues to improve locomotion and MEP conduction in rats with SCI after more than 6 weeks. The mechanisms underlying this effect might involve the promotion of axonal regeneration and the inhibition of astrocyte proliferation. Astrocyte linear reorientation may be helpful for axon directional growth. In this study, we only measured locomotor function, electrophysiology, and histomorphology. Assessment of oligodendrocyte markers (such as MBP) should be included in the future because of their important role in axonal myelination and proper electrophysiological function. Further studies aimed at determining the molecular mechanisms should also be performed. Moreover, we chose only one stimulation pattern, which is not enough to obtain a definite conclusion regarding optimal stimulation parameters. More stimulation patterns, such as different frequencies and stimulus intensities, should thus be attempted.
Acknowledgments:The authors would like to express their thanks to Professor Yan-Xia Luo from the Department of Statistics, Capital Medical University for her statistical suggestion in this research.
Author contributions:Study design: JZB, XLH; study implementation and paper preparation: YXW, ZL; data analysis: JZB, YXW, ZL; oscillating field stimulator fabrication: GHZ, XLH. All authors approved the final version of this manuscript for publication.
Conflicts of interest:There are no conflicts of interest.
Financial support:This study was supported by the National Natural Science Foundation of China, No. 30801222 (to JZB). The funder had no roles in the study design, conduction of experiment, data collection and analysis, decision to publish, or preparation of the manuscript.
Institutional review board statement:This study was approved by the Animal Experiments and Experimental Animal Welfare Committee of Capital Medical University (supplemental approval No. AEEI-2021-204) on July 26, 2021.
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewers:Rodolfo Gabriel Gatto, University of Illinois at Chicago, USA; Yona Goldshmit, Monash University, Australia.
Additional file:Open peer review reports 1 and 2.

- 中国神经再生研究(英文版)的其它文章
- The importance of fasciculation and elongation protein zeta-1 in neural circuit establishment and neurological disorders
- Promoting axon regeneration in the central nervous system by increasing PI3-kinase signaling
- Microglial voltage-gated proton channel Hv1 in spinal cord injury
- Liposome based drug delivery as a potential treatment option for Alzheimer’s disease
- Retinal regeneration requires dynamic Notch signaling
- All roads lead to Rome — a review of the potential mechanisms by which exerkines exhibit neuroprotective effects in Alzheimer’s disease