
Coordination of Schwann cell myelination and node formation at the transcriptional level
2022-11-11 13:46FranziskaFrMichaelWegner
中国神经再生研究(英文版) 2022年6期
Franziska Fröb, Michael Wegner
Formation of the node of Ranvier as a highly coordinated event:Saltatory conduction ensures that information in the vertebrate nervous system is rapidly transmitted over large distances and efficiently processed in complex networks. It requires the insulation of axonal segments by myelin and the formation of highly structured nodes of Ranvier that are interspersed at regular intervals between successive myelin sheaths and regenerate the action potential as it propagates along the nerve (Rasband and Peles, 2021). To be functional, nodes of Ranvier and the adjacent paranodal and juxtaparanodal regions (jointly referred to as nodal complex) contain ordered arrays of voltage-gated ion channels. The nodal complex additionally contains a host of adhesion molecules that are either supplied by the neuron or by the axon-contacting glial cells. Formation, organization, structural stability, and maintenance of the node depend on multiple fine-tuned molecular interactions among these neuronal and glial adhesion molecules (Faivre-Sarrailh and Devaux, 2013; Rasband and Peles, 2021).
In the peripheral nervous system (PNS), Schwann cells are the glial cells involved in node formation as well as in myelin sheath production. Major Schwann cell contributions to the nodal complex include the L1-type cell adhesion molecule Nrcam, the extracellular matrix component gliomedin and the NF155 isoform of neurofascin. Nrcam and gliomedin are mainly found at the node and NF155 is predominantly localized to paranodal regions (Faivre-Sarrailh and Devaux, 2013; Rasband and Peles, 2021). For proper node formation, the production of these proteins in Schwann cells has to be coordinated with the generation of their interaction partners on the axonal side in the neuron. Additionally, a synchronization is required with the manufacturing of the various proteinaceous and lipid components of the myelin sheath in the glial cells themselves (Srinivasan et al., 2012). How this is achieved is poorly understood on the molecular level and in part addressed by a recent study that analyzed the transcriptional regulation of genes with relevance for node formation in Schwann cells (Saur et al., 2021).
Sox10 and Krox20 as regulators of node formation and myelination in the PNS:By transcriptomic comparison of the MSC80 Schwann cell line and genomeedited Sox10-deficient clonal derivatives, Nrcam, gliomedin and neurofascin were identified as differentially expressed genes. A strongly reduced expression in peripheral nerves of mice with Schwann cell-specific Sox10 deletion further substantiated the assumption that nodal genes in Schwann cells are under control of Sox10, a transcription factor with essential functions in most stages of Schwann cell development (Finzsch et al., 2010). Transient transfections identified at least one putative regulatory region within each of the three genes or in their vicinity that strongly activated reporter gene expression in response to Sox10. These evolutionary conserved regulatory regions contained functionally relevant binding sites for Sox10 and thus resembled previously identified Sox10-responsive regulatory regions from genes that code for structural components of the myelin sheath in Schwann cells such asMbp,MpzandGjb1(henceforth referred to as myelin genes) (Peirano et al., 2000; Bondurand et al., 2001).
Genome wide studies had furthermore shown that many of the Sox10-bound regions in the vicinity of myelin genes or genes with relevance for lipid biosynthesis (referred to as lipid biosynthetic genes) strongly overlap with regions that are occupied by Krox20 (also known as Egr2) in Schwann cells (Srinivasan et al., 2012). Considering the fact that loss-of-function experiments in the mouse established essential roles for both Sox10 and Krox20 in myelin formation, these data point to a cooperative transcriptional activation of myelin and lipid biosynthetic genes by Sox10 and Krox20 as a precondition for PNS myelination (Figure 1).
Interestingly, Sox10-responsive regulatory regions that were associated with the genes for Nrcam, gliomedin and neurofascin were also activated by Krox20. In the joint presence of both Sox10 and Krox20, reporter gene activation was even higher than in the presence of only one. Analysis of nodal gene expression levels and nodal morphology in mice heterozygous for both Sox10 and Krox20 furthermore confirmedin vivothe cooperation between the two transcription factors during node formation in the PNS. This argues that the same two transcription factors are major regulators of myelin and node formation in developing Schwann cells (Figure 1). Sox10 and Krox20 thereby ensure that both processes are already coordinated on the transcriptional level. In transiently transfected cells, regulatory regions of myelin genes were activated at higher levels by Sox10 than by Krox20, whereas regulatory regions from nodal genes relied more strongly on Krox20. While these findings should be cautiously interpreted because of the artificial experimental setting, they may be indicative of a fine-tuning mechanism. Krox20 is itself a target gene of Sox10 (Figure 1) so that its levels peak later during Schwann cell differentiation than those of Sox10. A preferential reliance on Krox20 may therefore ensure that nodal gene expression peaks slightly later than myelin gene expression.
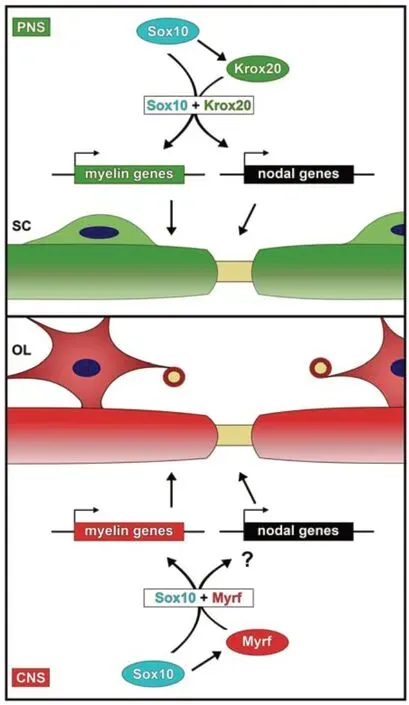
Figure 1|Regulatory mechanism for the coordinated expression of myelin and nodal genes in myelinating glia.
Open questions & future directions:While the study sheds light on the molecular coordination of myelination and node formation in Schwann cells, it does not address how transcription of nodal genes is regulated in neurons. In fact, little is known about the neuronal regulatory mechanisms. As Sox10 is not expressed in neurons, it is clear that other transcription factors must be in charge of nodal gene expression in neurons. This is especially intriguing for genes like neurofascin that are expressed (albeit in different isoforms) on both the axonal and the Schwann cell side but are apparently under the control of different transcription factors in both cell types. Considering that expression of nodal genes has to be coordinated across two cell membranes, it is very likely that reciprocal signals help to fine-tune gene expression in Schwann cells and neurons. Again, the identity of such signals is largely unknown. But axon-derived neuregulins and neuronal electrical activity are excellent candidates for an axon-to-Schwann cell signal. Both also impact Schwann cell myelination and would therefore further help to coordinate formation of node and myelin sheath.
Over the last decades, it has become clear that not only myelination, but also proper nodal assembly depends on the presence of myelinating glial cells (Faivre-Sarrailh and Devaux, 2013; Rasband and Peles, 2021). It is furthermore obvious that node formation and myelination need to be harmonized. This is true not only for the PNS, but also for the central nervous system (CNS) where oligodendrocytes take over the function played by Schwann cells in the PNS (Figure1). However, there are significant differences between both compartments of the nervous system. In the PNS, myelin formation starts at birth and in mice is almost complete within 14 days. In contrast, myelin formation in the mouse CNS takes approximately until one month of age although the process starts at about the same time (Rasband and Peles, 2021). In both PNS and CNS, a contact must be established between the terminal cytoplasmic loop of the myelinating glial cell and the axon at the paranodal junction to properly form nodes of Ranvier (Rosenbluth, 2009). However, the axo-glial contact at the node itself differs substantially. In the PNS, Schwann cell microvilli contact the adjacent axolemma. Oligodendrocytes do not form such microvillous processes. Instead nodal regions contain an extracellular matrix complex in the CNS (Nelson and Jenkins, 2017). The relative impact of paranodal versus nodal mechanisms for overall node formation also differs between PNS and CNS (Rasband and Peles, 2021).
In the same vein, Schwann cells and oligodendrocytes apply similar overall principles of transcriptional regulation for their differentiation and the myelination process, but rarely employ the exact same transcription factors and control mechanisms. The role of Krox20 as a central general regulator of myelination is restricted to Schwann cells. In oligodendrocytes, it occurs only in a subpopulation where its exact role has yet to be studied (Floriddia et al., 2020). In the context of myelination, many of the roles of Krox20 in Schwann cells are taken over in oligodendrocytes by Myrf, a structurally unrelated, ER-bound transcription factor with autoproteolytic activity (Bujalka et al., 2013). Like Krox20 in Schwann cells, so is Myrf a direct target gene of Sox10 in oligodendrocytes and once induced cooperates with Sox10 to jointly activate myelin and lipid biosynthetic genes (Hornig et al., 2013) (Figure 1). If myelination and node formation are similarly coordinated on the transcriptional level in oligodendrocytes as they are in Schwann cells, Myrf may have a comparable role in CNS node formation as Krox20 in the PNS and could cooperate with Sox10 during the process. However, considering the aforementioned differences in node structure, the higher variability of node formation in the CNS and the contribution of other glial cells such as astrocytes and NG2 glia to CNS nodes, other scenarios including a more flexible coupling between myelin sheath and node formation appear plausible as well. In any case, it is very likely that even if Myrf is the relevant cooperating partner of Sox10 during node formation in oligodendrocytes, different regulatory regions will be used for nodal gene activation in both glial cell types. Fortunately, candidate regulatory regions in the CNS can be identified from available ChIP-Seq data so that this question can be readily tackled in future studies.
The present work was supported by grants from the Deutsche Forschungsgemeinschaft (DFG) to MW.
Franziska Fröb, Michael Wegner*
Institut für Biochemie, Emil-Fischer-Zentrum, Friedrich-Alexander-Universität Erlangen-Nürnberg, Erlangen, Germany
*Correspondence to:Michael Wegner, Dr.rer.nat., michael.wegner@fau.de.
https://orcid.org/0000-0002-6404-8631 (Franziska Fröb)
https://orcid.org/0000-0002-4586-3294 (Michael Wegner)
Date of submission:March 31, 2021
Date of decision:May 19, 2021
Date of acceptance:July 9, 2021
Date of web publication:October 29, 2021
https://doi.org/10.4103/1673-5374.327343
How to cite this article:Fröb F, Wegner M (2022) Coordination of Schwann cell myelination and node formation at the transcriptional level. Neural Regen Res 17(6):1269-1270.
Copyright license agreement:The Copyright License Agreement has been signed by both authors before publication.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.

- 中国神经再生研究(英文版)的其它文章
- The importance of fasciculation and elongation protein zeta-1 in neural circuit establishment and neurological disorders
- Promoting axon regeneration in the central nervous system by increasing PI3-kinase signaling
- Microglial voltage-gated proton channel Hv1 in spinal cord injury
- Liposome based drug delivery as a potential treatment option for Alzheimer’s disease
- Retinal regeneration requires dynamic Notch signaling
- All roads lead to Rome — a review of the potential mechanisms by which exerkines exhibit neuroprotective effects in Alzheimer’s disease