
The power of “touch” and early enriched stimulation: neuroplasticity effects in rodents and preterm infants
2022-11-11 13:46AlbertoFernndezTeruel
中国神经再生研究(英文版) 2022年6期
Alberto Fernández-Teruel
Early postnatal stimulation, e.g., neonatal handling (NH) in its most frequent form, and environmental enrichment (EE, the exposure of juvenile animals, usually during several weeks, to environments involving rich and variable sensory stimulation) produce profound and longlasting behavioral and neurobiological effects. Both treatments reduce anxiety and stress sensitivity, and improve neurodevelopment and learning/memory in unconditioned and conditioned tasks in laboratory rodents. In addition, both manipulations lead to long lasting ‘‘protective’’ effects against age-related hippocampal neurodegeneration, cognitive deficits and associated stress-related neuroendocrine processes (e.g., Meaney et al., 1988; Fernández-Teruel et al., 1997, 2002). The present commentary is focused on summarizing relevant evidence on the enduring positive effects of early enriched sensory (NH- or EE-like) stimulation on neurobehavioral development and neuroplasticity (including the promotion of neural regeneration or the prevention of neurodegeneration) in rodents, and to discuss the possible clinical relevance and translatability of similar treatment approaches to humans.
The power of “touch”:from NH treatment in rodents to massage therapy of preterm human infants: At least part of the long lasting neurodevelopmental effects of exposure to NH or to early (prenatal and infantile) EE in rats have been attributed to the presence of enhanced “positive” maternal behavior (e.g. increases of licking/grooming of pups; Weaver et al., 2007; see Guzzetta et al., 2009). NH, EE and high levels of positive maternal care lead to enduring beneficial effects on behavior, cognition, and hypothalamuspituitary-adrenal axis function. Moreover, these environmentally-enriched conditions influence hippocampal glucocorticoid receptors, hippocampal synaptogenesis and function, and neuroplasticity through (for example) their effects on nerve growth factor-inducible protein A-, brain-derived neurotrophic factor- and glutamate receptor-mediated processes (Fernández-Teruel et al., 2002; Weaver et al., 2007). Following the idea that tactile stimulation (i.e., “touch”), such as that provided by the mother rats, was a crucial factor in the long-lasting beneficial effects of these early sensory-enriched treatments, Schanberg and Field (1987) tried to model the licking-grooming behavior of the mother by stroking the rat pups with a soft wet paintbrush. This kind of stimulation, administered during the first neonatal weeks reversed the negative effects of maternal separation stress on growth and growth hormone secretion. Likewise, the neonatal stimulation reversed maternal separation stress effects on ornithine decarboxylase activity (the first enzyme in the synthesis of polyamines, which are involved in the regulation of protein synthesis), on hypothalamus-pituitaryadrenal axis function (Schanberg and Field, 1987), as well as on brain-derived neurotrophic factor and synaptophysin expression (for further references see Guzzetta et al., 2009). Along these lines, Field and co-workers applied a similar stimulation treatment (called “massage”) to preterm human infants (Schanberg and Field 1987; Field et al., 2008), and found that such a massage therapy led to faster weight gain, increased vagal tone, lower cortisol levels and decreased stress-related behavior. Massaged infants showed better neurodevelopmental scores and earlier discharge from the hospital. Importantly, in addition to a greater weight gain, the preterm neonates receiving massage therapy exhibited increased levels of serum insulin and insulin-like growth factor-1 (IGF-1; Field et al., 2008).
The specific mechanisms underlying the effects of massage therapy on brain development and plasticity in human infants are still an open issue. The study by Guzzetta et al. (2009) was outstanding in this respect. They found that, in rat pups and preterm infants, massage therapy improved the maturation of visual evoked potentials, visual acuity and electroencephalographic activity across all brain regions (i.e., across all the electrodes; Guzzetta et al., 2009). EE treatment of rat pups from postnatal days (PND) 1 to PND12, also improved visual acuity development at PND25. Most importantly, these authors reported that blood levels of IGF-1 and insulin growth factor binding protein 3 were increased in massaged infants, and an increased number of IGF-1-positive neurons was found in the visual and auditory cortex of massaged rat pups. Moreover, circulating cortisol levels were reduced in massaged infants and corticosterone levels were reduced in the stimulated rat pups. Remarkably, the antagonism of IGF-1 activity reversed the effects of massage in rat pups, as well as the positive developmental effects of EE (administered between PND18 and PND25) on visual acuity in rats (see Ciucci et al., 2007; cited by Guzzetta et al., 2009). These findings suggest a causal role of IGF-1 function in mediating the brain maturational effects of tactile (massage) and EE stimulation (Guzzetta et al., 2009). These studies by Guzzetta and co-workers constituted the first clear evidence of positive brain maturational effects of neonatal massage therapy in human infants. These are important findings, since it is known that chronic excessive cortisol inhibits growth hormone secretion and weight gain, and the latter hormone normally stimulates IGF-1 synthesis. Thus, this suggests that by decreasing cortisol and increasing IGF-1, massage therapy in preterm human neonates (and NH treatment in rats) seems to facilitate weight gain and, according to Guzzetta et al. findings, accelerates brain maturation (Schanberg and Field 1987; Field et al., 2008; Guzzetta et al., 2009).
Postnatal tactile and enriched sensory stimulation and recovery from brain damage:Altogether, massage therapy in preterm human infants, and enriched sensory stimulation (NH, EE) in rodents, appear to display potent and long-term positive effects on development, brain development and neuroplasticity. Do such neuroplasticity effects extend to positive neuroregeneration effects in brain-injured subjects? As said above, both NH and EE treatments are capable of preventing the age-related neuronal degeneration in the hippocampus (Figure 1), and this goes along with NH and EE complete prevention of age-related spatial learning/memory deficits (Meaney et al., 1988; Fernández-Teruel et al., 1997;Figure 1). It has been proposed that a likely mechanism mediating NH and EE prevention of age-related hippocampus-dependent impairments might involve the attenuation of the ‘‘glucocorticoid-glutamate-calcium’’ neurotoxic cascade (Fernández-Teruel et al., 2002). But, are these early sensoryenriched treatments able to reverse or attenuate neuronal damage produced by brain insults? Brain injury induced by perinatal hypoxia-ischemia (HI) is an early brain insult that is directly related to perinatal morbidity and various neurodevelopmental deficits, and preterm infants are at increased risk of HI injury. Male human infants are more vulnerable than females to brain insult. In one study, male and female mice were submitted to HI (PND7) and NH treatment (PND1-21). Sensorimotor, behavioral, cognitive and brain neuropathological measures were taken at PND70-90 (Muntsant et al., 2019). Male mice appeared to be more vulnerable to HI-induced long-term memory impairment, and NH attenuated such a deficit more clearly in males than in females (Muntsant et al., 2019). NH treatment significantly attenuated HIinduced neuropathology in several brain regions (hippocampus, thalamus, caudate/putamen, neocortex, corpus callosum) in males, whereas the effect of NH was also significant but of lower magnitude in females (corpus callosum and neocortex). However, the neuropathological HI effects were statistically similar between both sexes, which could be because the authors did not use unbiased stereological quantification procedures (Muntsant et al., 2019).
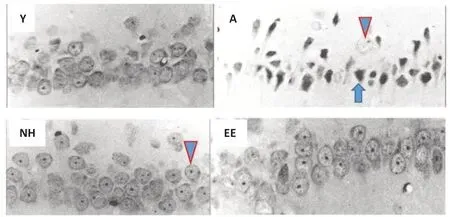
Figure 1|NH and EE treatments display lifelong protective effects against neurodegeneration in the rat hippocampus.
The findings by Muntsant et al. (2019) are generally in line with the study by Rodrigues et al. (2004), who showed that postnatal tactile (NH-like) stimulation of male rats enduringly prevented the HI deleterious effects on hippocampal and dentate gyrus volume in adulthood (Rodrigues et al., 2004). HI-induced neurodegeneration is attributed to excessive glutamate function and calcium influx (leading to excitotoxicity). In this regard, we have found suggestive evidence that NH treatment enduringly protects against N-methyl-D-aspartate-induced seizures in rats (Fernández-Teruel et al., 2002).
Remarkably, prenatal and early postnatal EE in male rats has also been found to enduringly attenuate the HI-induced deleterious neurodevelopmental and cognitive effects. The treatment also attenuates the hippocampal molecular and cellular damage produced by HI, and prevents the increase of amyloid precursor protein and the decrease of growth associated protein 43 in hippocampus and cerebral cortex (growth associated protein 43 is a protein related to neuronal regenerative processes, axon terminal growth and experiencedependent neuroplasticity; Durán-Carabali et al., 2020). In a related vein, it is also noteworthy that EE of adolescent male rats (administered from PND35 to PND57-66) rescues the impairments of adult neuroplasticity (LTP) and hippocampal neurogenesis induced by pre-pubertal stress (administered between PND25-27; Rule et al., 2020). Such an EE treatment also reverses amygdala functional alterations and hippocampus-dependent cognitive impairments produced by early life stress (Rule et al., 2020). Of note, none of the aforementioned EE studies used isolated animals as controls (i.e., in all cases they used animals housed in groups of > 2 per cage), which improves the possible translatability of their findings to humans.
Given the increased vulnerability of male subjects to perinatal brain insults (Muntsant et al., 2019), and that most studies have used only one sex for analysis in this field (an exception is Muntsant et al., 2019), it is necessary to design protocols including sexual dimorphism as an objective of study. Moreover, more precise and unbiased (stereological) methods of neuropathological analysis are desirable for future studies.
Conclusions and perspectives:Enriched sensory stimulation, applied during the neonatal (and even prenatal), prepubertal or adolescent periods, most commonly in the form of massage therapy, NH or EE (involving soft tactile and multisensory stimulation), has a dramatic and long lasting impact on development, neurodevelopment and neuroplasticity. Depending upon the particular treatments and whether they are applied to human infants or rodents, such effects are (among others): (i) increased growth hormone, IGF-1 and weight gain; (ii) improved cortical maturation/neuroplasticity; (iii) reduced stress-related behaviors and hypothalamus-pituitary-adrenalaxis activity; (iv) increased levels of neurotrophic factors; (v) increased brain neuroregeneration following brain insults (e.g., HI); (vi) increased hippocampal neurogenesis and cognitive function; and (vii) decreased age-related hippocampal neurodegeneration and cognitive impairments. A recent neurobiological model proposes that early tactile and multisensory stimulation (NH or EE) enhance glutamate N-methyl-Daspartate receptor-mediated synaptic activity/plasticity (e.g., long term synaptic potentiation), which modulates dopaminergic, opioid and serotoninergic pathways in limbic areas and other regions (e.g., prefrontal cortex) within the neural reward systems. The resulting enduring structural changes are supported/maintained by neurotrophic factors (e.g. brain-derived neurotrophic factor), whose function may in turn be facilitated by the above activated serotoninergic circuits (Fu and Depue 2019).
Whatever the specific neuro-molecular mechanisms, given the enormous and long-term neuroplasticity potential (particularly according to animal studies) of massage/tactile stimulation (NH) and EE, this allows proposing that these interventions may have a future important value in clinical conditions. Massage therapy is increasingly being applied in infant neonatal care units with positive results (Guzzetta et al., 2009; Niemi, 2017). Most interestingly, some forms of EE interventions are also being evaluated in clinical controlled trials. It is worth mentioning, in that regard, the pilot randomized controlled trials carried out by Finlayson, Dusing and colleagues (Finlayson et al., 2020). In both randomized controlled trials (Australia, and Virginia, USA) the authors have evaluated the effects of an EE-like program, named SPEEDI (“Supporting play, exploration, and early development intervention”), which involves multisensory stimulation of preterm infants (5 days/week) starting in the neonatal care unit and continuing at home, for a total of approximately 15 weeks (Finlayson et al., 2020, and references therein). The findings of these randomized controlled trials are very promising, showing the superiority of the SPEEDIvs. “usual care group” in some measures of problem-solving behavior, and motor, receptive and expressive communication scores at approximately 1-month follow-up (Finlayson et al., 2020, and references therein). This shows how EE-like protocols, “conceptually” similar to those used in rodent research, may be used in human infants, and indirectly suggests that the SPEEDI stimulation program might optimize neurodevelopmental processes, thus providing the basis for conducting larger clinical trials (Finlayson et al., 2020, and references therein).
There is still a long way to go to establish whether the long lasting effects of NH and EE on neurobehavioral development and neuroplasticity observed in rodents are generalizable to humans. This will require more comprehensive and longitudinal clinical studies, which also include more direct measures of brain function as well as neuroendocrine parameters, along the lines (for example) of the study by Guzzetta et al. (2009).
The author gratefully acknowledges support from grants PSI2017-82257-P, PID2020-114697GB-I00, and 2017SGR-1586 (to AFT).
Alberto Fernández-Teruel*
Department of Psychiatry and Forensic Medicine & Institute of Neurosciences, School of Medicine, Autonomous University of Barcelona, Bellaterra, Barcelona, Spain
*Correspondence to:Alberto Fernández-Teruel, PhD, Albert.fernandez.teruel@uab.cat.
https://orcid.org/0000-0001-5993-7058 (Alberto Fernández-Teruel)
Date of submission:March 15, 2021
Date of decision:April 21, 2021
Date of acceptance:June 19, 2021
Date of web publication:October 29, 2021
https://doi.org/10.4103/1673-5374.327336
How to cite this article:Fernández-Teruel A (2022) The power of “touch” and early enriched stimulation: neuroplasticity effects in rodents and preterm infants. Neural Regen Res 17(6):1248-1250.
Copyright license agreement:The Copyright License Agreement has been signed by the author before publication.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewer:Luz Elena Durán-Carabali, Universidade Federal do Rio Grande do Sul, Brazil.
Additional file:Open peer review report 1.

- 中国神经再生研究(英文版)的其它文章
- The importance of fasciculation and elongation protein zeta-1 in neural circuit establishment and neurological disorders
- Promoting axon regeneration in the central nervous system by increasing PI3-kinase signaling
- Microglial voltage-gated proton channel Hv1 in spinal cord injury
- Liposome based drug delivery as a potential treatment option for Alzheimer’s disease
- Retinal regeneration requires dynamic Notch signaling
- All roads lead to Rome — a review of the potential mechanisms by which exerkines exhibit neuroprotective effects in Alzheimer’s disease