
Application of neurotrophic and proangiogenic factors as therapy after peripheral nervous system injury
2022-11-11 13:46KamillaFaritovnaIdrisovaAlinaKazymovnaZeinalovaGalinaAndreevnaMasgutovaAlexeyAndreevichBogovJrCinziaAllegrucciValeriiaYurievnaSyromiatnikovaIlnurIldusovichSalafutdinovEkaternaEvgenievnaGaraninaDinaIvanovnaAndreevaAdiletAbdullaa
中国神经再生研究(英文版) 2022年6期
Kamilla Faritovna Idrisova, Alina Kazymovna Zeinalova Galina Andreevna Masgutova Alexey Andreevich Bogov Jr., Cinzia Allegrucci, Valeriia Yurievna Syromiatnikova Ilnur Ildusovich Salafutdinov Ekaterna Evgenievna Garanina Dina Ivanovna Andreeva Adilet Abdullaatovich Kadyrov, Albert Anatolevich Rizvanov, Ruslan Faridovich Masgutov
Abstract The intrinsic ability of peripheral nerves to regenerate after injury is extremely limited, especially in case of severe injury. This often leads to poor motor function and permanent disability. Existing approaches for the treatment of injured nerves do not provide appropriate conditions to support survival and growth of nerve cells. This drawback can be compensated by the use of gene therapy and cell therapy-based drugs that locally provide an increase in the key regulators of nerve growth, including neurotrophic factors and extracellular matrix proteins. Each growth factor plays its own specific angiotrophic or neurotrophic role. Currently, growth factors are widely studied as accelerators of nerve regeneration. Particularly noteworthy is synergy between various growth factors, that is essential for both angiogenesis and neurogenesis. Fibroblast growth factor 2 and vascular endothelial growth factor are widely known for their proangiogenic effects. At the same time, fibroblast growth factor 2 and vascular endothelial growth factor stimulate neural cell growth and play an important role in neurodegenerative diseases of the peripheral nervous system. Taken together, their neurotrophic and angiogenic properties have positive effect on the regeneration process. In this review we provide an in-depth overview of the role of fibroblast growth factor 2 and vascular endothelial growth factor in the regeneration of peripheral nerves, thus demonstrating their neurotherapeutic efficacy in improving neuron survival in the peripheral nervous system.
Key Words: fibroblast growth factor 2; growth factors; nerve growth factor; peripheral nerve injury; peripheral nervous system; vascular endothelial growth factor 1Institute of Fundamental Medicine and Biology, Kazan Federal University, Kazan, Russia; 2Republican Clinical Hospital, Kazan, Russia; 3Biodiscovery Institute, School of Veterinary Medicine and Science, University of Nottingham, Nottingham, UK; 4Department of Traumatology and Orthopedics, Kazan State Medical Academy, Kazan, Russia
Introduction
Peripheral neuropathies represent an important clinical problem. Approximately 50% of adult patients with diabetes develop peripheral neuropathies of varying severity during their lifetime (Hicks and Selvin, 2019). Despite the prevalence of the disease, there is no universally effective treatment for the most severe forms that develop due to axonotmesis and neurotmesis. Currently, one of the most effective methods of treatment of neurotmesis is surgical reconstruction of the damaged nerve. The surgical intervention always carries an additional risk for the patient, including possible development of infection and exacerbation of diseases of other organs. However, even with surgical treatment, successful reinnervation of the target organs is achieved only in less than 50% of cases (Grinsell and Keating, 2014). As a result, the number of studies focusing on the use of gene therapy and tissue engineering for treatment of peripheral neuropathies has increased significantly in recent years. Combined with gene therapy, tissue engineering provides promising applications for the treatment of traumatic nerve injuries (Moimas et al., 2013). The promising results stem from the modulation of favorable conditions for directed axon growth and angiogenesis. In addition, one of the advantages of gene therapy can be systemic delivery of the drug when the damage of the peripheral nervous system (PNS) is multiple or its localization is difficult to specify.
Cells and microenvironment, as well as “scaffolding”, are the key factors forming the basis of therapeutic application of tissue engineering and regenerative medicine (Tabata, 2009). The microenvironment plays an important role, while it provides a complex of bioactive molecules and factors regulating self-renewal and differentiation of stem cells. Among microenvironmental factors, biochemical signals (growth factors, cytokines, and extracellular components, such as vesicles) and biophysical signals (mechanical deformation, damage, loss of bonds) can be identified. Growth factors (GFs) are of particular importance because of their influence on signaling pathways and regeneration potential (Tayalia and Mooney, 2009). Physiological signals modulated by GFs are essential for controlling cellular processes such as chemotaxis, proliferation and differentiation. The number of studies reporting on the isolation, identification, and practical application of GFs is growing exponentially, as they are actively used to promote regeneration in the peripheral nervous system (Allen et al., 2015; Zhao et al., 2015; Krishnan and David, 2017) (Table 1). When placed inside the conduit’s microenvironment, GFs can improve axon growth and make the whole channel application more efficient (Dalamagkas et al., 2016). These results therefore suggest that superior outcomes in nerve regeneration may be obtained using GFs alone or in combination with conduit application instead of autografts (Wood et al., 2009; Roam et al., 2014).

Table 1 |Growth factors involved in peripheral nerve regeneration
To this end, new techniques have been studied based on delivery of active molecule (affinity-based delivery) (Wood et al., 2009). To deliver therapeutic compounds or to create the microenvironment for more efficient regeneration, a huge variety of hydrogels, microspheres, acellular matrices, threedimensional “scaffolding”, and fibrin glues of biological and synthetic origin are used, as well as various types of vectors for delivery (Figure 1). The design of neural conduits requires information on neural architecture, type of injury, nerve cells, scaffold materials, neural growth factors to add and optimize the mechanical properties of the conduit. 3D Biorinated Peripheral Nerve Tubes serve as nerve grafts to fill in the gaps in destroyed nerve bodies, making the peripheral nerve canals alive (Soman and Vijayavenkataraman, 2020). Over the past decades, strategies have been developed involving the use of biopolymers, cells, growth factors and physical stimuli, which have led to the development of various nerve conductors (NGCs), from simple hollow tubes to complex pipelines involving one or more landmarks (Sarker et al., 2018).
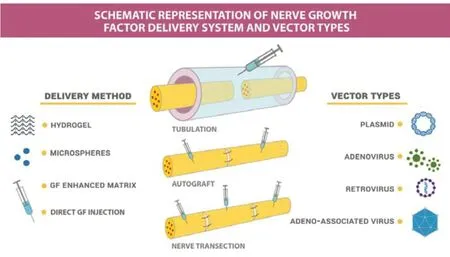
Figure 1|Schematic representation of nerve growth factor delivery systems and vector types.
Implantation of artificial NGC used to bridge the damaged area can provide the appropriate biochemical and biophysical landmarks needed to stimulate regeneration and restore PNS function. Improved canal design and fabrication techniques have made it possible to fabricate autograft-like structures at NGC (Li, 2019).
Many delivery systems have been designed to provide custom delivery kinetics for one or more growth factors. However, selection of the optimal method for delivering GFs is problematic since they are sensitive to environmental conditions and can easily lose therapeutic efficacy (Atienza-Roca et al., 2018; Subbiah and Guldberg, 2019). New effective methods of treatment of peripheral nervous system injuries are likely to include artificially created tissues and biomaterials that can cause tissue microenvironment, which includes both biochemical and mechanical properties that contribute to regeneration (Wilcox et al., 2020). Therefore, it is essential to evaluate the physicochemical nature of GF and biomaterials used to develop biomimetic delivery systems when GF-based therapy of traumatic injuries is considered (Subbiah and Guldberg, 2019).
Search Strategy and Selection Criteria
Studies cited in this review published from 1973 to 2021 were searched on the PubMed, Scopus, and Google Scholar database using the following keywords: fibroblast growth factor 2, growth factors, nerve growth factor, peripheral nerve injury, peripheral nervous system, vascular endothelial growth factor.
Key Growth Factors in the Nervous System
There are several major families of growth factors which act within the nervous system (Table 1). Some of them, e.g. neurotrophins, are abundant only in the nervous system, while others, like fibroblast growth factor (FGF), vascular endothelial growth factor (VEGF), insulin-like growth factor receptor-1, act on a number of cell types over the body, in addition to the nervous system (Landreth et al., 1999). Neurotrophins are highly conserved small proteins that dimerize to form biologically active isoforms (Lewin and Barde, 1996). Neurotrophin family includes five different members: Nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), neurotrophin 3 (NT3), neurotrophin 4/5 (NT4/5), and neurotrophin 6 (NT6) (Landreth et al., 1999).
NGF is the prototype of the neurotrophin family and is absolutely essential for the survival of sympathetic neurons and a subset of sensory neurons during development. NGF is one of the earliest and most studied neurotrophic factors, which has many biological function including protection and promotion of the nerve growth (Lewin and Barde, 1996; Duraikannu et al., 2019). NGF is synthesized in tissues that are innervated by innervated target tissues , in immune cells and components of PNS and central nervous system (CNS) (Aloe et al., 2012). NGF stimulates cell metabolism, morphological differentiation of target cells, contributing to the extensive axonal and dendritic growth (Sofroniew et al., 2001). One of the recent trends in peripheral nerve reconstruction research is the development of tissue engineering scaffolds that facilitate release and bioactivity rates of nerve growth factors. Thus, novel thiolated gellan gum modified with laminin and loaded NGF developed by Li et al. (2020b) demonstrated excellent biocompatibility and the ability to stimulate the growth. Their study of thiolated gellan gum gel loaded with NGF has promising applications in peripheral nerve reconstruction and shows high potential of application of tissue engineering-based scaffolds (Li et al., 2020b). The development of an effective nerve allograft treated with a dual preparation containing acrolimus and nerve growth factor has been shown to be highly effective in stimulating the growth of nerve cells under conditions of hypoxia caused by nerve damage (Yin et al., 2019).
Ligands of NGF and GDNF families of neurotrophic factors also play an important role in the regeneration of PNS (Piirsoo et al., 2010). It was found that exogenous administration of BDNF enhances myelination in the PNS, compared to the removal of endogenous BDNF that inhibits myelination in mice (Chan et al., 2001; Tolwani et al., 2004). Furthermore, BDNF is indispensable for re-myelination of the injured peripheral nerves in rodents (Zhang et al., 2000). Expression of members of NGF and GDNF families in the damaged nerve is regulated specifically for each gene in the course of time (Meyer et al., 1992; Funakoshi et al., 1993). Administration of exogenous FGF21 promotes functional and morphological recovery in a rat model of sciatic nerve injury, manifested as improved motor and sensory function, increased axonal remyelination, and re-growth and accelerated proliferation of Schwann cells (Lu et al., 2019b).
Functional recovery of nerves is highly dependent on the synergistic interaction between angiogenesis and neurogenesis processes after injury. Cross-talk between vascularization and nerve regeneration in the PNS plays an important role in the recovery of the PNS. Regenerative strategies through the synergistic delivery of multiple biochemical signals are gaining increasing attention, especially combination of pro-angiogenic factors and neurotrophic factors (Lu et al., 2019a).
VEGF and FGF affect angiogenesis and neurogenesis and may act synergistically with other neurotrophic factors. Thus, synergistic effect of NGF and VEGF was shown in the study of Li et al. (2020a), where GFs included into porcine decellularized neural matrix (pDNM) hydrogel improved functional nerve recovery afterin situadministration (Li et al., 2020a). Moreover, dual delivery of VEGF and NGF plasmids as gene therapy can enhance sciatic nerve regeneration (Fang et al., 2020). In the early stages of sciatic nerve injury, these GFs not only enhance nerve regeneration, but also facilitate vascular penetration, and after 12 weeks promote nerve regeneration and functional recovery in rats (Rao et al., 2020). In addition, a three-dimensional neurovascular microenvironment for endothelial and nerve cell growth can be mimicked by self-assembling nanofiber peptide hydrogel with dual-presenting VEGF-and BDNF-mimetic peptide epitopes (Lu et al., 2019a).
Despite the fact that there are growth factors that have been associated with regeneration in the PNS for a long time and their synergism has been well studied, growth factors associated with angiogenesis and wound healing, such as VEGF and FGF, also have neurotrophic effects, and their synergism will be discussed further in the article.
Growth Factors in Nerve Regeneration
Among wide gamut of growth factors, neurotrophic factors demonstrate cytoprotective and regenerative effects on the nervous system. These are widely used as therapeutics to prevent the progression of damage in peripheral nerves (Hernandez-Morato et al., 2016). Neurotrophic GFs affect inflammatory and proliferative processes and the course of wound healing (Shi et al., 2013). Among them, FGF-2 and VEGF are the most investigated (Zhang et al., 2002).
The process of regeneration of nerve fibers is complex and multifactorial. Usually, endogenous factors released after injury are not enough for effective repair, especially in case of severe combined injuries. Due to this, functional impairment of the central nerve system occurs after spinal cord injury, stroke, ischemia, traumatic injury and associated axonal disconnection. The outcome is different in the PNS, as long-distance axon regeneration and substantial functional recovery can happen in the adult (Huebner and Strittmatter, 2009). However, the prognosis for such injuries is hard to be improved without additional therapeutic or surgical interventions. For full functional recovery, regenerating axons must restore their connections with skeletal muscle fibers, smooth muscle and epithelial cells (Coletti et al., 2013). The process of regeneration of the nerve fiber is inextricably linked with wallerian degeneration through complex interaction of a large number of biologically active substances and cells involved in their synthesis (Deumens et al., 2010). Therefore, the delivery of neurotrophic factors to the damaged area is considered to be one of the most promising approaches to stimulate neuroregeneration (Zhang et al., 2002; Mason et al., 2011). However, studies on the stimulation of posttraumatic regeneration in peripheral nerves using direct gene therapy are limited (Fu et al., 2007; Esaki et al., 2011). Indeed, effective delivery of therapeutic doses of GFs in the nervous system can be difficult due to the special features of neuroanatomy (Langert and Brey, 2018).
The delivery of GFs can be performed by using recombinant proteins as well as gene and gene-cell therapies. Together with nerve guidance conduits, they act through releasing molecules that enhance axonal growth and accelerate regeneration and recovery of functions (Carballo-Molina and Velasco, 2015). Therefore, their delivery to the PNS has the potential to be used for treatment of various diseases, such as neuropathic pain (Aloe et al., 2012), traumatic nerve injury (Boldyreva et al., 2018), and acute or chronic pain (Llorian-Salvador and Gonzalez-Rodriguez, 2018).
Biology of Fibroblast Growth Factor 2
The basic fibroblast growth factor, bFGF or FGF2, was first discovered in pituitary extracts (Armelin, 1973) and then identified in bovine brain extracts as a component causing fibroblast proliferation (Gospodarowicz, 1974). FGF2 belongs to a large family of growth factors that includes 23 members (Imamura, 2014), which share common functional and structural features (Ornitz and Itoh, 2001). FGFs use paracrine or endocrine signaling in developmental and pathophysiological processes, including metabolism regulation, angiogenesis, wound healing, embryonic development and regulation of endocrine secretion (Aimi et al., 2015; Owen et al., 2015). Through FGF receptors (FGFRs) binding, FGF2 regulates a broad spectrum of biological functions, including cellular proliferation, survival, migration, and differentiation. FGF-2 stimulates proliferation of fibroblasts, vascular endothelial cells and keratinocytesin vitro, as well as new vessels formation, tissue granulation, epidermal regenerationin vivo. In addition, it regulates homeostatic processes in the nervous system (Nedeau et al., 2008). FGF2 is one of the central factors that are essential for the development of CNS and PNS. During embryonic development, high levels of FGF-2 are detected from neurulation onwards and either FGF1 or FGF2 can induce proliferation of neural crest cellsin vitro(Murphy et al., 1994). Moreover, expression of FGF-2 and its receptors is temporally and spatially regulated during development and neurogenesis in specific brain regions (Powell et al., 1991).In vivo, FGF-2 expression is activated both at the site of injury and in the bodies of neurons after nerve damage. It also interacts with its receptors on Schwann cells, stimulating their proliferation (Allodi et al., 2014).
In vitroresearch initially indicated that the bFGF was biologically active, as it could induce lengthening of neurites after its release from synthetic guidance channels. A study of rat sciatic nerve repair demonstrated increased expression of FGF-2 and its receptors in the dorsal root ganglia at the proximal and distal stumps of the peripheral nerve after injury, four weeks after implantation of nerve guidance channels containing bFGF and connecting the two stumps of the nerve by keeping nerve bundles with both myelinated and nonmyelinated axons (Aebischer et al., 1989). Therefore, it was demonstrated that locally applied bFGF mediated regenerative effects on damaged sensory neurons and supported axonal growth (Grothe and Nikkhah, 2001). It was also shown that neurotrophic factors stimulate the recovery of peripheral nerves and act during the early regeneration of the sciatic nerve, regulating the proliferation of Schwann cells. Indeed, a study using FGF-2 deficient mice indicated that FGF-2 is important for axonal maturation and remyelination one week after sciatic nerve crush (Jungnickel et al., 2004). Later it was also shown that the level of FGFR1/2 in sensory neurons rises after peripheral nerve injury and differentiation of Schwann cells and axons is induced. FGF-2 induces proliferation of Schwann cells and inhibits myelination through FGFR1/2 (Haastert et al., 2009).
Considering its importance in neural development, the role of FGF-2 has also been investigated in neurodegenerative diseases, including Alzheimer’s disease, multiple sclerosis, Parkinson’s disease and traumatic brain injury (Woodbury and Ikezu, 2014).
Biology of Vascular Endothelial Growth Factor
VEGF is a potent angiogenic factor that was first described as essential factor for growth of vascular endothelial cells. VEGF functions via interaction with VEGF receptor 1 (VEGFR-1), VEGF receptor 2 (VEGFR-2), VEGF receptor 3 (VEGFR-3), and co-receptors neuropilin-1 and neuropilin-2 (Muratori et al., 2018). Diverse biochemical properties of VEGF and its cooperation with other GFs defines whether VEGF functions to stimulate proliferation or angiogenesis (Maharaj and D’Amore, 2007). Its predominant physiological role is to accelerate accumulation of endothelial cells in the blood vessel network during vasculo- and angiogenesis (Pereira Lopes et al., 2011). New research on both CNS and PNS emphasize the importance of VEGF in neuronal survival and proliferation (Dumpich et al., 2015).
There are many examples of neurons and glial cells of the PNS and CNS which are stimulated by VEGF, such as Schwann cells, dorsal root ganglia cells, hippocampal neurons, oligodendrocytes or Purkinje cells (Dumpich et al., 2015). For example, it has been shown that VEGF is necessary for the development of visual chiasm. In addition, a recent study demonstrated the positive effect of VEGF-B in neuroregeneration (Guaiquil et al., 2014). Evidence of the VEGF importance in the nervous system comes mostly from studies on the CNS. However, its role in the PNS may be different from that in the CNS, as CNS has a limited capacity for regeneration in contrast to the PNS, where damage is often accompanied by partial or full recovery of nerves (Zochodne, 2012).
Studies conducted on PNS have shown that VEGF-induced growth is enhanced after damage and it may play a role in neuroprotection. Indeed, VEGF has considerable effect on the growth of peripheral trigeminal neurons innervating the cornea and dorsal root ganglion neurons (Yu et al., 2008). In addition, VEGF is responsible for the directed growth of axons of the dorsal root ganglia, invasion of Schwann cells and neovascularization after sciatic nerve damage (Sondell et al., 1999). However, it is not yet known if VEGF has also a neuroprotective effect on sensory neuronsin vivo. Although VEGF therapy has already shown therapeutic potential in peripheral neuropathies, these effects are mainly associated with its stimulating effect on blood vessels, and not with direct neuroprotective properties (Kirchmair et al., 2007). Indeed, a study examining the effect of VEGF on sciatic nerve regeneration demonstrated a positive relationship between increased vascularization and enhanced nerve regeneration (Hillenbrand et al., 2015).
VEGF may also support and increase the growth of regenerating nerve fibers, probably through a combination of angiogenic, neurotrophic and neuroprotective effects mediated by multiple factors secreted by activated macrophages (Caillaud et al., 2019). Using crush injury and end-to-end injury damage models, it has been shown for the first time that VEGF induces peripheral neuritogenesis (Muratori et al., 2018). Differential expression of VEGF under conditions of regeneration and degeneration after damage to the peripheral nerve has been demonstrated using median nerve crush injury and median nerve transection followed by end-to-end microsurgical suturing. Using this model, it was shown that increased expression of VEGF165 accelerates Schwann cells migration, which is the main stimulus for the promotion of neurite outgrowth (Muratori et al., 2018). An increase in the level of VEGF can also promote axon regeneration and functional reinnervation by stimulating axon growth and proliferation of Schwann cells (Haninec et al., 2012). Moreover, the positive effect of VEGF on the angiogenesis and intraneural revascularization during peripheral nerve regeneration has been demonstrated by the enhancement of oxygen and nutrients supply that is needed to form Bühngner bands with Schwann cells. However, the molecular interactions between Schwann cells, macrophages, and neo-vascular endothelial cells needs further investigation (Caillaud et al., 2019). Saffari et al. (2020a, b) described application of the superficial inferior epigastric artery fascial flap to achieve vascularization to the nerve graft site in an experimental model and they found that it enhances revascularization of transplanted nerve allografts.
Synergy between Fibroblast Growth Factor 2 and Vascular Endothelial Growth Factor
The expression levels of FGF-2 and VEGF are interrelated. Synergy between these two growths factors has been demonstrated by the evidence that FGF-2 increases endogenous synthesis of VEGF and VEGF itself is necessary for FGF-2-induced expression of placental growth factor in vascular smooth muscle cells (Couper et al., 1997). In addition, VEGF-A and FGF-2 are well-studied pro-angiogenic molecules that are used for therapeutic angiogenesis (Kano et al., 2005). The combination of VEGF-A and FGF-2 has a powerful synergistic effect on the formation of new vessels in experimental conditions bothin vivoandin vitro. In addition to the combination of VEGF-A and FGF-2, VEGF-A and platelet-derived growth factor BB (PDGF BB) (Richardson et al., 2001) and FGF-2 and PDGF-BB (Cao et al., 2003) have a strong synergistic effect on induction of neovascularization in experimental animal modelsin vivo. However, the mechanisms that underlie these synergisms are not fully understood.In vitroexperiments have shown that, in addition to direct mitogenic effects, VEGF-A and FGF-2 enhance the intercellular transmission of PDGF-B signals in a specific way dependent on the cell types. Indeed, VEGF-A enhances the expression of endothelial PDGF-B, while FGF-2 enhances the PDGF receptor mural (PDGFR) expression. Co-stimulation using VEGF-A and FGF-2 causes enhanced formation of neovascular cells compared to stimulation with a single agent. Thus, enhanced signaling of endogenous PDGF-B-PDGFR is necessary for the synergistic effect of VEGF-A and FGF-2 on angiogenesis in adults (Kano et al., 2005). Several studies have shown that FGF2 can induce neovasculogenesis in an indirect way through activation of the VEGF/VEGFR receptor system. For instance, a study by Yanagita et al. (2014) showed a cooperative effect of FGF-2 and VEGF on periodontal ligament cells, where intercellular interactions between PDL cells and endothelial cells can stimulate angiogenesis.
Biomaterials for Delivery and Release of Growth Factors
Advanced technologies that ensure delivery and release of GFs at the site of injury are critical for achieving effective regeneration. The biomaterials used to deliver and release growth factors are of great variety. A wide range of biomaterials has been studied to find the most effective substrate for FGF production and release bothin vitroand/orin vivo, including synthetic and natural polymers and even tissue matrices. Initiatory synthetic nerve guidance conduits were initially made of silicone. Currently, biodegradable materials such as aliphatic polyesters, collagen, or polyurethanes are used (Pfister et al., 2007). These substrates can be combined with cells that actively secrete GFs to promote repair and regeneration. For instance, FGF-2 derived from marrow mononuclear cells promotes the regeneration of the peripheral nerve, stimulating the survival and proliferation of glial cells in a model of sciatic nerve damage with complete transection of the sciatic nerve, followed by connecting the proximal and distal sections inside a silicone tube (Ribeiro-Resende et al., 2012). This work supports the hypothesis that various molecules derived from the bone marrow work together during the regeneration of peripheral nerve, leading to the decrease of neuron death and the increase of axonal growth. Pfister et al. (2007) proposed application of nerve guidance conduits with integrated delivery systems for growth factors or growth factor producing cells in order to make nerve regeneration more effective. The delivery can be mediated by viral bacterial plasmids. Solovyeva et al. (2014) studied the effect of plasmid DNA encoding the pro-angiogenic GFs VEGF and FGF2 on cytokine production by human adipose tissuederived stem cells. Genetic modification of adipose tissuederived stem cells with the recombinant plasmid pBud-VEGFFGF2 results in increased secretion of IL-8 and MCP-1, both involved in wound healing.
A complementary technology is based on the use of scaffolds as prototype of extracellular matrix. Different materials are used for their manufacturing, including natural and synthetic polymers, as well as ceramics. Methods of delivery of factors that are most important for PNS recovery vary greatly and are constantly being improved (Table 2).

Table 2 |Delivery methods for GFs that are the most important for regeneration of PNS
Implantation of scaffolds allows quickly covering of the defect and recovery of the function of damaged nerves (Whitaker et al., 2001). It is important to emphasize that endogenous expression of GFs together with exogenous supplementation in the nerve regenerative environment can further improve outcome. Most studied growth factors used in this context include NGFs, FGFs, BDNF, GDNF, ciliary neurotrophic factor, VEGF, and NT-3. All of them are known to have different affinities to specific ECM proteins (Pabari et al., 2011). Wagner et al. (2018) synthesized biodegradable scaffolds from polycaprolactone fumarate for tissue ingrowth through large interconnected pores. As a result, scaffolds seeded with microspheres containing VEGF or VEGF with BMP-2 or FGF-2 have a significantly higher level of vascular ingrowth. Thus, this method of delivery of growth factors shows potential for tissue regeneration in various types of tissues (Wagner et al., 2018).Application of functionalized materials for neural scaffolds together with GFs can considerably improve outcome. However, the optimal method of incorporation needs further investigation (Du et al., 2018). Giannaccini et al. (2017) demonstrated that synthetic NGC provide physical guidance to the regenerating stump and limit scar tissue infiltration at the site of injury. VEGF and NGF on magnetic nanoparticles can improve the rates of regeneration and this can be considered as a realistic alternative to autografts (Giannaccini et al., 2017).
Growth Factors and Therapies for Nerve Regeneration
Several models have been developed for the study of GFs and their effects on reparative processes. Delivery of GFs is used for increasing the expression of these genes in the damaged area. One of the possible options is GFs injections into a damaged nerve or conduit using microcapsules or minipumps designed to provide constant or prolonged concentration of the solution in the tissue. Transplantation of cells secreting the necessary factors or the use of plasmids promoting production of neurotrophic and angiogenic substances are also widely used (Pfister et al., 2007; Masgutov et al., 2018). Notably, regenerating nerves can get into neuroma, a phenomenon indicating the effect of “candy store” linked to local elevation of GF concentration causing trapping of regenerating axons (Eggers et al., 2013).
Peripheral nerve injury is a major neurological disorder that can cause multiple motor and sensory impairment. Guaiqui et al. (2014) showed that VEGF-B stimulates nerve regeneration and improves the restoration of tissue sensitivity as well as the ability of nerves to improve healing and regeneration of innervated tissue. Indeed, VEGF-B induces strong lengthening and branching of neurons and requires specific transmembrane receptors, as well as the activation of complex intracellular signaling (Guaiquil et al., 2014).
At the same time, it was shown that injections of FGF-2 using retroviral delivery lead to a significant improvement in the regeneration of nerves and re-innervation (Allodi et al., 2014). Heparin-based release of bFGF in the rat sciatic nerve model showed how FGF affects functional recovery and regeneration of the peripheral nerve.In vivo, the coacervate improves the bioavailability of bFGF, thus improving both motor and sensory nerve function. It can also speed up regeneration and stimulate the proliferation of Schwann cells (Li et al., 2017).
Recent studies revealed that VEGF also stimulates neurogenesis, neuronal patterning, neuroprotection and glial growth. VEGF is an important participant in the neuroregeneration due to the close relationship between nerve fibers and blood vessels (Hobson et al., 2000). Application of VEGF significantly increases the permeability of blood vessels with an increase in axon regeneration and migration of Schwann cells after axotomy. Reinnervation of the target-organ is facilitated by VEGF, thus demonstrating a mutual relationship between increased vascularization and enhanced nerve regeneration (Hobson et al., 2000).
Furthermore, delivery of a plasmid vector with the VEGF gene into the rat sciatic nerve transection area improves regeneration and contributes to the recovery of motor function (Fu et al., 2007). At the same time, allotransplantation of FGF2-transfected Schwann cells as part of a silicone conduit implanted in a defect of the rat sciatic nerve supports the regeneration of myelin fibers (Haastert et al., 2009). Using the rat sciatic nerve insertion model, the effectiveness of local injection of the plasmid pBud-VEGFFGF2, expressing both factors at the same time, was also evaluated. Direct injection of this plasmid into the central and peripheral stumps of the nerve, as well as into the autograft, stimulates the regeneration and recovery of motor function (Masgutov et al., 2011). VEGF treatment of cultured neurons autonomously of blood vessels increases neurite survival and its growth. VEGF also supports the migration of neurons in the embryonic brain and supports co-growth of axons and arteries in developing skin (Rosenstein et al., 2010). In addition, intramuscular overexpression of VEGF using AAV-VEGF vectors considerably reduces the progression of muscle atrophy in a rat nerve injury model (Moimas et al., 2013).
Shen et al. (2020) also evaluated the regenerative capacity of peripheral nerves after nerve injury. They observed the dynamic changes of genes in L4-6 dorsal root ganglion after rat sciatic nerve crush using transcriptome sequencing. Their data showed that many growth factors, including nerve growth factor, brain neurotrophic factor, fibroblast growth factor 2, and amphiregulin, are involved in tissue remodeling and biological processes associated with axonal growth. Their experiment outlines the pattern of expression of growth factors in the dorsal root ganglia after peripheral nerve injury (Shen et al., 2020).
As mentioned above, damage of the PNS can include polyneuropathy, followed by deterioration of innervation and blood circulation of the target organ. In that case, the infiltration of a neurotrophic ulcer in the heel region at equidistant points with equal doses of the pBud-VEGF165-FGF2 plasmid, combined with vacuum therapy and classical surgical techniques, demonstrated a successful reparation of the ulcer after 5 months (Mullin et al., 2013).
Conclusions and Future Directions
Insufficient levels of GFs increase the probability of neurons undergoing apoptosis and compromise regeneration of PNS after injury. Therefore, the application of exogenous GFs is considered to be an effective therapy for the treatment of acute peripheral nerve injuries, as GFs support the regeneration of axons and formation of new myelin sheaths.
It is important to acknowledge that the combination of classical surgical methods for restoring peripheral nerves and methods of direct therapy with growth factors both accelerate axonal growth, stimulate nerve revascularization and the migration of Schwann cells. Due to the variable changes in nerves during different types of PNI, the aim of peripheral treatment is to maintain the pro-regenerative capacity of the de-axonised distal nerve and to promote recipient axonal regeneration. This is confirmed by numerous experimental studies demonstrating improvement of post-traumatic restoration of function and reinnervation of damaged tissue. Efficient use of GFs at appropriate sites in PNS can improve the regenerative capacity of tissue after trauma, despite their delivery still remaining a clinical challenge. Therefore, it is of utmost importance to define the best methods for GF delivery and to find optimal conditions for their most effective therapeutic use.
Author contributions:Manuscript concept, preparation, review, and literature search: KI, MG, AB, VS, IS, GE, DA, AK, RM; Editing: CA, AZ, AR. All authors approved the final manuscript.
Conflicts of interest:The authors declare no conflicts of interest.
Financial support:AAR was supported by state assignment 0671-2020-0058 of the Ministry of Education and Science of Russian Federation. IIS and GAM were supported by the Russian Foundation for Basic Research grant 18-54-45023 Ind_a. This work is part of Kazan Federal University Strategic Academic Leadership Program (to IIS).
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix,tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewer:Abu Rafee Malik, Indian Veterinary Research Institute, India.

- 中国神经再生研究(英文版)的其它文章
- The importance of fasciculation and elongation protein zeta-1 in neural circuit establishment and neurological disorders
- Promoting axon regeneration in the central nervous system by increasing PI3-kinase signaling
- Microglial voltage-gated proton channel Hv1 in spinal cord injury
- Liposome based drug delivery as a potential treatment option for Alzheimer’s disease
- Retinal regeneration requires dynamic Notch signaling
- All roads lead to Rome — a review of the potential mechanisms by which exerkines exhibit neuroprotective effects in Alzheimer’s disease