
山区天然林生态修复研究进展
2022-11-08 03:17:02胡天祺王振锡郝康迪
广西林业科学 2022年5期
胡天祺,王振锡,郝康迪,陈 哲,董 巍
(1.新疆农业大学 林学与风景园林学院,新疆乌鲁木齐 830052;2.新疆教育厅干旱区林业生态与产业技术重点实验室,新疆乌鲁木齐 830052)
天然林是森林资源的主体和精华,是自然界中群落最稳定、生态功能最完备和生物多样性最丰富的陆地生态系统,是维护国土安全最重要的生态屏障[1]。我国的生态文明建设日益受到重视,天然林的经营与管理逐渐成熟,相继出台了多个关于天然林的保护政策与方案。2019年,中共中央办公厅国务院办公厅印发《天然林保护修复制度方案》,包括完善天然林管护制度、建立天然林用途管制制度、健全天然林修复制度、落实天然林保护修复监管制度、完善支持政策和强化支持保障等方面。2020年,国家发展改革委自然资源部印发《全国重要生态系统保护和修复重大工程总体规划(2021 —2035年)》,总结我国生态保护和修复工作的成效,提出生态保护与修复中存在的主要问题,并对全国重要生态系统保护和修复重大工程进行布局。我国有23.04%的大陆领土被森林覆盖,其中天然林约占中国森林总面积的64%[2]。山地森林是指位于山地系统和各个山脉中的森林(相对高度波动超过100 m,从山脚到山脉顶部或到无树空间边界的平均表面坡度超过5°)及高山高原和高原上的森林,不包括丘陵高地上的森林。对山区天然林进行修复可促进人与自然和谐共生、满足人民群众对优质生态环境的需求[3-4]。
1 生态修复的概念
生态修复概念的提出可追溯至200年前;19世纪后期,Cairns 主编的《Restoration of Damaged Eco‑systems》(《受损生态系统的恢复过程》)一书出版后,开始将生态修复作为生态学的1 个分支进行研究。各国在进行生态修复研究时,关于生态恢复、生态修复、生态重建和生态改良等概念虽表述不同,但其核心理念可归纳为恢复与发展两大方面,即让最初受到自然或人为干扰损害的生态系统得到恢复,使其能可持续发展,被人们长期利用[5-8]。总的来说,生态修复是对受到干扰和破坏的生态环境进行修复,使其尽可能恢复到原来的状态,使原来定居的物种能重新定居,或使与原来物种相似的物种能定居,将受干扰和破坏的土地恢复到具有生产力的状态,确保该土地保持稳定的生产状态,不再继续恶化[9-12]。
2 国外山区天然林生态修复案例与研究进展
2.1 水平-垂直结构三维可视化
Dubravac 等[13]在研究克罗地亚普利特维采湖国家公园严格保护区山毛榉(Fagus longipetiolata)森林的自然再生时,通过野外测量和水平-垂直结构三维可视化方法,对幼树的再生、生长、发育和存活过程进行研究。该研究重点分析了1998—2009年山毛榉群落树冠结构发育和自然更新的过程,通过记录树木的空间排列与高程信息绘制水平树冠投影,并在数字高程模型(Digital elevation model,DEM)上进行记录,在3 个2 m × 60 m(面积360 m2)条状样地内记录幼苗和灌木层的高度结构(图1),并将它们分为6个高度等级(<30、31~60、61~130、131~150、151~200和201~250 cm)。结果表明,1株优势山毛榉幼枝再生核的发生是由其枯背引起的,这为其进行自然再生提供了可能;研究期间,山毛榉幼苗数量变化最大,从1998年的3 556 株/hm2增至2009年的12 694株/hm2,增长了近4 倍,山毛榉幼苗所占比例从8.7%上升至22.6%,占据重要地位,但山毛榉幼苗大多质量较差,在被动保护条件下能否进一步发展,需进一步观察。该研究结果表明,通过水平-垂直结构三维可视化方法研究冠状结构动力学和自然再生的过程,是一种新途径,可为观测林分发展趋势提供更生动的表达,并可为取得的成果提供高质量展示。在气候变化的条件下,通过1 个可靠的网络试验模型持续改进监测方法,对于森林保护是必要的[14-15]。
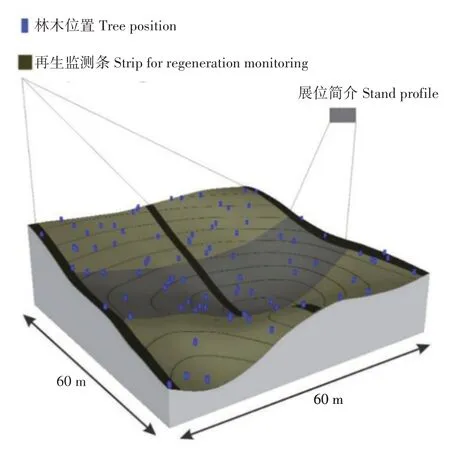
图1 带有基本测量元素的试验区数字高程模型[13]Fig.1 Digital elevation model of test plot with basic survey elements
2.2 Schrenk云杉(Picea asperata)丰度模型
哈萨克斯坦山区森林约占全国森林总面积的40%。Mambetov 等[16]在对北天山山林中的Schrenk云杉进行自然再生评价时,发现北天山山区森林中Schrenk 云杉生态修复的方法,并对其进行评估。研究区Ketpen-Temerlik 山脉(79°30′ ~ 85°00′ E,43°20′~44°00′N)位于北天山系,是其最东端;整个山脊长约400 km,宽约40 ~ 50 km。在Ketpen-Te‑merlik 山区森林中,Schrenk 云杉是主要的森林树种,自然生长在海拔1 500~2 900 m 的地区,主要分布在天山北、西北和东北坡上。Ketpen-Temerlik 山脉最高峰海拔3 680 m,位于与中国接壤的东部;向西逐渐降低至3 400 m。在哈萨克斯坦,Ketpen山脊北部与Dzungar Alatau 接壤,被伊犁洼地分隔;在东部,Ketpen 山脉逐渐进入Temerlik 山脉,并与Boro‑horo 山脉合并。Mambetov 等根据海拔高度、坡度和方位角编制太阳辐射的年到达量,在此基础上,构建Schrenk 云杉丰度模型。结果显示,Schrenk 云杉的自然再生不能在所有森林条件下均以相同方式进行;由于环境条件不同,每个试验监测站点均存在生态修复限制,有必要考虑场地条件(区域绝对高度、暴露量和坡度)。为在林区中心建立模型,从森林的下部边界到上部边界选择几条峡谷或1个大的坡度,在绝对高度上每100 m 铺设1 条水平通道,将云杉幼苗和灌木丛分为3个高海拔组。
Schrenk云杉生长缓慢,田间条件下很难确定其灌木丛的林龄,该指标只能通过根颈处的切口来判断。Чешев 等[17]根据云杉幼龄生长特征和死亡率的具体情况,开发了1个评估Schrenk云杉自然更新的量表。根据量表尺度,云杉的所有自播和灌木均按高度分为3 组(20 cm 以下、20 ~ 50 cm 和50 cm 以上),最终估值以超过50 cm 的树丛数目为依据。结果表明,幼苗生长和灌木丛的研究应在高海拔群体中进行;林分冠层下的更新说明林分的高密性不是自生云杉出现的限制因素,但它阻碍较老灌木丛的生存;在高密度林分中,云杉的灌木丛仅出现在直径为1.4~1.8 m 的树冠内腔,在这种情况下,光成为其生存的主要限制因素。在天山北部的云杉林中,随林分密度降低、树龄增加,成年植物和地被植物间的竞争对灌木丛的生存产生负面影响[17-20]。
2.3 自然扰动模型分析
俄罗斯的Omelko 等[21]采用树木生态学方法重建俄罗斯Sikhote-Alin 山脉南部古生代红松(Pinus koraiensis)-阔叶林的扰动历史。过程基于1 种假设,即林分中树木径向生长的增加是由于周围树木死亡后生存条件的改善,研究者使用以下公式计算每个样本在一段时间内径向生长百分比(GC,%)的相对变化:

式中,M1 为过去10年的平均增长;M2 为未来10年的平均增长(可通过年度气候波动进行预测)。根据该方法,对数据进行拟合,得到特定的边界线曲线。GC值在特定的M1值上大于边界线函数值的20%或50%,分别表明次要增长释放和主要增长释放。每个GC值与标记树的某个核心关联,可以确定给定年份中具体某个个体经历的增长释放。过去200年间,频繁的小规模扰动和罕见的中度扰动对增长释放产生影响(图2)。
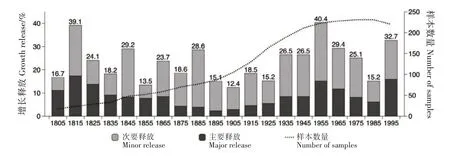
图2 俄罗斯远东南部红松-阔叶林(俄罗斯远东南部Sikhote-Alin)中5种主要乔木的生长释放[21]Fig.2 Growth releases of five dominant trees in a mixed Korean pine-broadleaved forest in South of the Russian Far East(Southern Sikhote-Alin,Russian Far East)
研究人员对5种优势树种的种群结构进行点格局分析。结果表明,1800 — 2000年间,5 种优势树种均未发生强烈的林分更替干扰,低强度和中等强度的扰动占主导地位,产生这些扰动最可能的原因为大风;高级更新集主要由耐荫针叶树和耐光落叶树的未成熟植物组成,说明落叶树种的未成熟植物有较强的耐荫性,且程度低于针叶树。原始植物中,针叶和落叶植物对光的需求较大。由于优势种的种群较稳定,当前的干扰机制促进林分中耐荫、耐光树种的共存,但影响林分中各植物的丰度。研究结果表明,应限制对红松阔叶林的木材采伐,因为树木再生需很长时间;还需制定具体的森林管理制度,模仿自然干扰制度(以中、小规模干扰为主),这是所有树种成功更新的必要条件。频繁的砍伐促进先锋树种更新,但长寿树种没有足够的时间发展,林分将迅速退化[22-25]。
2.4 人工促进再生
在泰国东北部萨卡拉特森林生态系统恢复影响的研究中,KAMO 等[26]对单个外来种或本地种的不同人工林林下植物种类进行对比,发现在泰国东北部Sakaerat 天然森林中,附近林分内生长的植物种类比在草原内生长的植物种类多。森林加速草地的次生演替,在重新造林前,草地的演替是通过反复发生的火灾实现的。在人工林林下树种的自然更新中,天然林中的大型树种似乎是所研究林分中主要的种子来源。这些人工林的小叶能提供足够的遮荫条件,并在森林发育的早期为幼苗生长提供适宜的光环境。
研究倾向于认为,植树是一种生产木材和纤维素等可再生资源的方法,利用人工林促进树种的自然再生是1 个与传统方法非常不同的概念;这两种做法不应是替代的,而应是互补的,以建立具有高度生物多样性的森林生态系统[27-31]。相比种植单一树种的森林,物种丰富的生态系统可在景观水平上实现高度的生物多样性,可给森林带来各种好处,包括木材生产、水和环境保护、碳储存及教育和风景娱乐;为实现这一目标,需在重新造林的地区保留一些天然森林,避免大规模伐光[32-33]。
3 国内山区天然林生态修复案例与研究进展
3.1 生态阈值的负反馈
上个世纪,由于自然灾害和经济发展需求,中国的森林生态系统遭受严重破坏。森林生态修复的首要任务是恢复退化的森林生态系统,自然恢复是最好的选择,但需很长时间才能满足需求[34-35]。根据自然规律进行人工干预是加速退化森林生态系统恢复的首选辅助方式。20世纪80年代以来,中国的森林恢复倡议在短期内取得显著成果,但未能根据响应阈值进行人工干预也在一定程度上引起森林的生态退化。因此,科学家和环境管理人员高度重视森林生态系统的恢复举措。在中国福建省长汀县森林退化生态系统的研究中,研究人员从寻找最佳树冠覆盖度的角度为天然林的生态修复提供新参考[36]。结果显示,将林区的冠层覆盖率限制在最佳水平,可维持生态系统的稳定性并防止冠层覆盖率过高时可能出现的潜在不利后果(林下植被消失等);当植被覆盖率降低到特定退化阈值以下时,将出现植被群落持续退化、表层土壤侵蚀和土壤肥力下降等现象。说明在缺乏全面人工恢复措施的情况下,在一定的树冠覆盖度下进行自然生态修复是困难的。研究人员也使用树木覆盖率为预测变量,使用其他生态指标为响应变量(植物种类、灌木和草皮、土壤养分及土壤侵蚀模量等),分析自然恢复和人工恢复是否存在响应阈值。结果表明,自然恢复地块中,生态系统的树冠覆盖度与一些关键生态指标呈非线性关系;一种情况是生态系统在任何树冠覆盖度水平上均不稳定(图3);另一种情况是在自然恢复开始后很长时间开始稳定(图4)。根据研究的回归结果预测,树冠覆盖率超过82.2%时,灌木和草本植物的覆盖率开始稳定(R2=0.976 9),侵蚀模量(R2=0.804 6)和径流(R2=0.895 5)将在树冠覆盖率分别超过56.3%和89.8%时开始稳定。说明自然恢复产生的生态系统比植树造林产生的生态系统更稳定。生态修复是一个渐进的过程,在人为干预后可能存在显著的结果时间滞后。研究发现,滞后时间比大多数修复时间要长得多,应增加生态恢复的持续时间,增加发现变化的可能性,而不仅仅确定短期影响[36-38]。
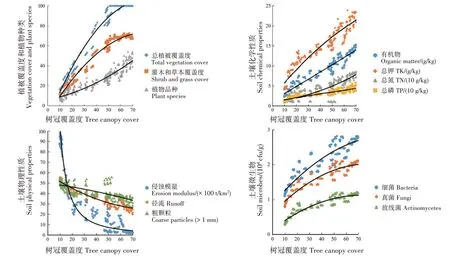
图3 树冠覆盖度与关键生态参数间的关系[36]Fig.3 Relationships between tree canopy cover and key ecological parameters
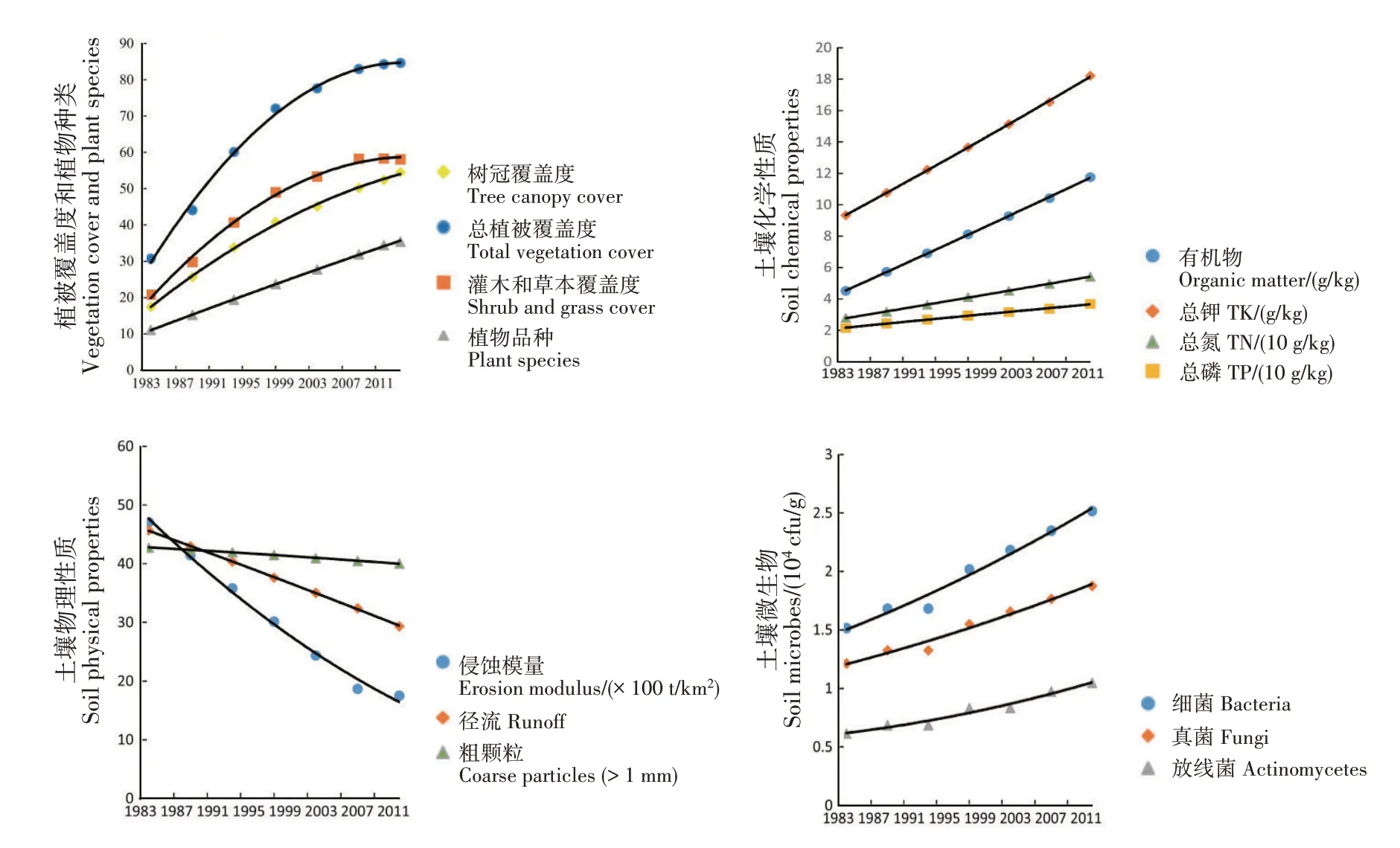
图4 样地中关键生态参数随时间的变化[36]Fig.4 Variations of key ecological parameters in sample plots with time
3.2 土壤侵蚀的控制
土壤侵蚀主要由水引起,是土壤退化的表现形式之一,对全球陆地生态系统构成严重威胁。在森林恢复对三峡库区水土流失影响的研究中,发现植被恢复是减轻不同景观和人为干扰水平土壤流失风险的1 种有前景的策略[39]。由于复杂的地形条件和土地利用,森林恢复对土壤侵蚀的影响存在不确定性,特别是在山地库区进行密集人类活动的前提下。研究者评估中国三峡库区森林恢复对土壤侵蚀的控制效果,利用修正的通用土壤流失方程和时间序列数据估算三峡库区2001—2015年的土壤侵蚀及其变化,在像元水平上估算土壤侵蚀坡度,确定土壤侵蚀对森林恢复的响应。结果表明,2001— 2015年,土地利用/覆盖变化以退耕还林为主,三峡库区年均土壤侵蚀速率呈下降趋势,年下降率为1.28%;土壤侵蚀速率的变化存在显著的空间差异,25°~35°间地形坡带的土壤侵蚀速率显著降低,林地恢复较密集(图5)。在不同土地转化过程中,林地平均土壤流失率的斜率最大。说明森林恢复可有效减少山地库区土壤侵蚀,但在不同植被恢复过程中差异显著,且存在时滞效应和海拔梯度差异。虽然大部分森林恢复发生在陡坡地区,但坡度仍是空间上的主导因子。随区域生态修复工程实施,大面积耕地被改造成林地或灌丛。自然植被覆盖度提高,降水和地表径流侵蚀减少。新森林主要分布在陡峭的斜坡地区(N 15°),积极的造林政策能更好地满足这些地区水土流失控制的需要,对土壤侵蚀的控制效果比三峡库区中实施的其他土壤保持政策更有效。
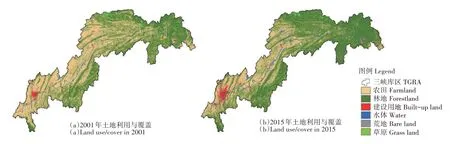
图5 2001与2015年三峡库区土地利用/覆盖空间分布[39]Fig.5 Spatial distribution of the land use/cover in 2001 and 2015 in the Three Gorges Reservoir area
4 山区天然林的生态修复重点方向
4.1 改进和扩大检测方法
在气候变化的情况下,有必要通过所有保护区的永久性试验区网络不断改进和扩大监测方法。计算机化模型可获得高质量且可靠的信息,结合遥感方法和数据(机载图像和卫星图像等),可表征大规模区域的森林动态。高分辨率卫星图像在森林冠层建模(用于表征森林的间隙动态)方面有很大潜力,可通过遥感和GIS 技术协助森林生态系统的生态修复过程。激光雷达在林业特别是在高海拔森林和垂直结构中已取得一系列成功;因传感器高度有限,普通的光学传感器很难提供森林的垂直资料,但激光雷达可通过采样的方式被感知,配置高频脉冲激光,可准确提供森林的水平和垂直信息[40-41]。除激光雷达外,也可通过其他技术获取高精度数字高程数据,例如合成孔径雷达(Synthetic aperture radar,SAR),目前有机载与星载两种搭载方式,为高精度数字高程数据的获取提供了新途径,也是当下林业遥感的热点。
4.2 制定完善、具体的森林管理制度
有学者认为,在中国的干旱和半干旱地区,种植不适合当地环境的树木,以目前的绿化政策(包括大规模苗木和树木种植项目),可能会破坏生态条件,新种植的树木无法改善环境,可能导致环境恶化;在脆弱的干旱和半干旱地区进行大规模造林,会增加缺水的严重性,减少造林地的植被覆盖,并对物种多样性产生负面影响[42]。法规和政策应在国家计划的区域内更灵活地制定,地方政府应努力在国家计划指南的框架内创建地方(县、乡和村等)一级的自我可持续政策[43]。
4.3 考虑不同区域的限制
森林规划至关重要,各种活动的影响会随地区不同而不同。中国的自然和社会经济状况因地区而异,因此在国家林业计划的指导下,区域项目应着重于区域特定条件和需求。在确定不同山区天然林的合适恢复措施时,对生态恢复区进行分类研究至关重要。为了解特定区域潜在的森林恢复需求,需详细调查该森林区域的具体状况。详细的分区有助于确定关键区域,并优先进行恢复计划和评估工作。我国已确定了进行森林恢复的某些关键区域,如北京和天津附近的生态修复区、西北黄土高原水蚀生态修复区和长江中游生态修复区[44]。山区天然林的修复对环境的影响是深刻而全面的,但在一定程度上,这些影响尚不清楚,需进一步评估。因此,有必要对恢复工作的影响进行更多研究,提高恢复工作的有效性。这些影响可能包括不同恢复情景对水流模拟、养分循环、森林演替和物种多样性变化等自然资源方面的综合影响,这些相关问题需进一步研究[45-47]。
4.4 共同进行自然再生与人为干预
在进行森林经营的过程中,错误的经营方法可能导致大量山地森林遭到破坏。我国小兴安岭的红松林在不进行人工干预、完全通过自然再生恢复的情况下,恢复至其原始状态大约需要1 000年的时间;天山山脉通过自然更新天山云杉林,大约还需要近两个世纪才能恢复初始状态;因此,在进行自然再生的同时需进行科学的人工干预,不应坐等自然恢复[48-50]。进行人工生态修复可按照要求,选择目的树种,通过合理密度或混交等方式加速更新过程。在生态修复过程中,需注意以下几点:造林树种及其比重和混交模式的选择;清理伐区;造林密度与造林方法;幼林抚育管理制度;幼所需种苗量及工作量。多方面因素共同作用,才能适应我国生态恢复及林业发展的需要。
5 小结与展望
国内外学者运用多种方法与技术对不同地区山区天然林的生态修复进行研究,从单一树种到局部森林生态系统,再到具体地区的生态整体层面,进行了多方面的研究与分析,肯定了山区天然林对当地生态环境的调节作用,同时结合遥感和地理信息系统为山区天然林的生态修复提供新的技术手段。纵观世界,各个国家在山区天然林修复方面的研究有所差异,但整体研究方向一致,均认为山区森林对于生态环境的作用至关重要。人们已认识到环境对人类发展的重要性,森林在整个生物圈中的作用也越来越受到重视,国内外学者们关于山区天然林修复研究所得出的结论对荒漠化防治、森林经营和树木病虫害防治等方面的发展均起到了重要的参考作用。
猜你喜欢
山西林业(2021年2期)2021-07-21 07:29:28
农村百事通(2020年21期)2020-11-23 02:04:24
绿色中国(2019年19期)2019-11-26 07:13:16
绿色中国(2019年19期)2019-11-26 07:13:16
中国水土保持科学(2019年6期)2019-04-26 05:14:42
作文大王·笑话大王(2017年1期)2017-02-21 16:08:53
湖南城市学院学报(自然科学版)(2016年2期)2016-12-01 04:06:46
作文大王·笑话大王(2016年10期)2016-10-18 14:58:58
作文大王·笑话大王(2016年7期)2016-08-08 11:28:43
湖南城市学院学报(自然科学版)(2016年4期)2016-02-27 14:02:42
