
氮添加对青藏高原高寒草甸土壤磷组分的影响
2022-11-04 08:11耿依仪邓艳芳王宇飞周华坤邵新庆李芝雯
草地学报 2022年10期
韩 冰, 耿依仪, 邓艳芳, 王宇飞, 周华坤, 邵新庆*, 李芝雯
(1.中国农业大学草业科学与技术学院, 北京 100193; 2.祁连山国家公园青海服务保障中心, 青海 西宁 810000; 3. 中国科学院西北高原生物研究所, 青海省寒区恢复生态学重点实验室, 青海 西宁810001; 4.青海省祁连县农牧 水利科技和乡村振兴局, 青海 西宁 810400)
磷作为影响陆地植物生产力的重要营养元素,是维持生态系统功能稳定的关键因素[1]。大部分磷被难溶性化合物吸附和固定,只有一小部分磷可被植物和微生物吸收利用[2,3]。尤其在石灰性土壤中,磷的流动性和生物有效性不能满足植物的需求[4]。青藏高原是我国重要的生态安全屏障[5],高寒草地占其总面积的53%,近1.4亿公顷[6]。高寒草甸作为高寒草地的植被类型之一,多位于干旱和半干旱地区,且低温限制了该地区氮、磷的矿化过程[7],土壤中可被植物吸收利用的氮、磷含量缺乏[8],养分限制模式为磷限制或氮磷限制。
氮是限制陆地生态系统初级生产力的关键营养元素[9]。额外添加氮素可以增加土壤中的无机氮含量从而提高植物生产力。有研究表明,氮添加会通过增加地上植物的生产力从而增加植物对土壤有效磷的需求,从而加重了陆地生态系统(包括森林、草地、湿地和苔原)的磷限制[10],同时植物也会释放羧酸盐来调控土壤磷周转[11]。氮添加条件下土壤理化性质的变化也影响着土壤磷的转化。有研究得出土壤硝态氮是驱动不稳定有机磷变化的重要因素[12],氮添加通过积累硝酸盐致使土壤酸化[13],促进了石灰性土壤中磷酸盐的溶解[14],但在酸性土壤中,土壤pH值的降低会促进土壤中的金属阳离子(如铝和铁)与磷形成难溶化合物,从而抑制酸性土壤中有机磷矿化成为有效磷的过程[15]。氮添加还会通过改变微生物群落影响微生物对有效磷的吸收[16]及土壤磷酸酶的活性[17]。
为维持和提高高寒草地生态系统的稳定性和适应性,施用氮肥是常见的养分管理措施之一[18]。在高寒草甸开展的一些长期施肥试验研究结果表明,氮添加极大的促进了地上生物量的提高[19]。目前的研究主要通过测定高寒草甸土壤中全磷和有效磷含量去分析土壤磷在氮添加处理下的变化特征[20-21],但对于土壤磷的各组分之间如何变化及氮添加与磷组分之间影响关系的研究还不够深入。土壤磷的分级可以进一步提供土壤中磷的迁移和有效性的信息[22],由此我们提出两个科学假设:(1)在氮添加条件下,高寒草甸土壤的活性磷组分含量会显著增加;(2)氮添加会通过降低土壤pH值促进土壤无机磷的溶解,还会通过增加微生物活性和磷酸酶活性促进土壤有机磷的矿化,从而增加土壤中磷的生物有效性。
综上所述,本研究以青藏高原高寒草甸为研究对象,通过在野外定位试验平台上实施氮添加的试验处理,将土壤中的磷按照有效程度进行分级,探究土壤磷组分在氮添加下的变化特征,并进一步探究氮添加下土壤的理化性质和生物特性与土壤磷组分之间的关系,深入理解氮添加条件下高寒草甸生态系统土壤特性对土壤磷转化特征的影响,为探究青藏高原高寒草甸可持续管理利用提供理论依据和技术支撑。
1 材料与方法
1.1 研究区域概况
养分添加试验地位于青海省海晏县西海镇(36°92′ N,100°93′ E),依托青海省海北州高原现代生态畜牧业科技试验示范园建立。海拔高度范围在3 000~3 100 m,该地区离青海湖较近,为高原大陆性气候,全年日照总长大约有2 580 h,特点是雨热同期,夏季凉爽湿润,冬季寒冷干旱。年平均降水有400 mm,主要集中在6—8月。年均温为1.5℃,最高月均温22.9℃,最低月均温-24.8℃。试验地的草地类型属于高寒草甸草原,土壤为粘壤土,样地中的主要植物种类有针茅(Stipaaliena)、赖草(Leymussecalinus)、洽草(Koeleriamacrantha)、冰草(Agropyroncristatum)、扁蓿豆(Melissitusruthenica)、矮嵩草(Kobresiahumilis)、星毛委陵菜(Potentillaacaulis)、二裂委陵菜(Potentillabifurca)和冷蒿(Artemisiafrigida)。植物生长集中在6—9月,生长期较短,8月初为植物生长旺盛时期。
1.2 试验设计
养分添加试验样地建立于2019年6月。其中,有三个氮添加水平,分别为5 g·m-2(N5),10 g·m-2(N10)和15 g·m-2(N15)。一共3个氮添加处理,加上对照(CK)共4个处理,设置4个重复,总计16个小区。每个小区的大小为3 m×4 m,各小区之间设置1 m的间隔。养分添加的处理安排在每年草地的返青期(6月初),氮添加以尿素(N含量46%)的形式每年施入一次。在试验期间,样地周围有围栏围护,采用封闭管理。
1.3 取样方法
在连续施肥两年后的生长高峰期(2020年8月初),使用0.5 m×0.5 m的样方取各小区的植物样品,将样方内的植物贴地刈割后烘干称重,记作植物地上生物量(g ·m-2)。使用土钻(直径5 cm)在各小区内取0~10 cm的土样,各小区重复取样5次,充分混合后过粗筛(2 mm),筛去根系和大块土粒,分成2份,1份风干后常温保存,用于测定土壤理化性质;1份储存在-20℃冰箱,用于测定土壤无机氮、酶活性和微生物生物量。
1.4 试验方法

植物样品用硝酸-高氯酸消煮30 min后用比色法测定植物磷含量(g ·kg-1),地上植物整株含磷量(g ·m-2)为植物磷含量乘以植物生物量。
土壤磷组分的测定用Tiessen改进后的Hedley磷分级方法提取土壤中的磷素[23-24]。称取0.5 g的风干土壤至50 mL离心管中,分别用阴离子交换树脂膜、0.5 mol ·L-1NaHCO3、0.1 mol ·L-1NaOH、1 mol ·L-1HCl、浓HCl进行连续浸提,最后用浓H2SO4和H2O2于360℃消煮至液体澄清。有研究将Hedley磷分级方法测出的磷组分进行了土壤磷库的划分[25-26],将树脂提取态无机磷(Resin-Pi)和碳酸氢钠提取态无机磷(NaHCO3-Pi)划分为活性磷(Labile P),将氢氧化钠提取态无机磷(NaOH-Pi)、氢氧化钠提取态有机磷(NaOH-Po)和(1 mol ·L-1盐酸提取态无机磷)1 mol ·L-1HCl-Pi划分为中等有效磷(Moderately labile P),将浓盐酸提取态全磷(HCl conc.-Pt)和残余态磷(Residual-P)划分为稳定磷(Stable P),磷库的划分可以帮助我们更加系统的认识到磷的有效性。
1.5 数据分析
利用Shapiro-Wilk检验数据是否符合正态分布(P>0.05),单因素方差分析(One-Way ANOVA)用于分析土壤磷组分和土壤特性在氮添加下的变化,使用P<0.05的显著性水平。通过SPSS 19.0软件完成数据分析,绘图由Origin 2019和R 4.0.2共同完成。Pearson相关分析用于比较土壤特性和土壤磷组分之间的相关性,通过R 4.0.2中的ggplot2包绘制相关分析热图。
2 结果与分析
2.1 不同氮添加水平对土壤磷组分的影响
由图1可知,与CK相比,N5和N10处理下的活性磷显著增加了28.7%和43.8%;与CK相比,3个水平的氮添加都降低了土壤中等有效磷的含量,N5差异显著(P<0.05)。稳定磷在N10和N15显著降低18.0%和11.0%(P<0.05);总磷在N10和N15显著降低4.23%和5.86%(P<0.05)。

图1 氮添加对土壤磷库的影响Fig.1 Effects of N addition on soil P pool注:平均值±标准误;不同小写字母分别代表在0.05水平差异显著,n=4,下同Note:Mean±SE. Different lowercases letters indicate the significant difference at the 0.05 level,n=4,the same as below
由图2所示,相较于CK,土壤NaHCO3-Pi含量在N5中显著提高了44.1%(P<0.05);3个氮添加处理使土壤中HCl conc.-Pt的含量均有不同程度降低,N5和N10差异显著(P<0.05);N15处理下Residual-P的含量显著降低了15.1%(P<0.05);对于Resin-Pi,NaOH-Pi,NaOH-Po和1 mol ·L-1HCl-Pi,不同水平的氮添加对其含量都没有显著的影响。

图2 氮添加对土壤磷组分的影响Fig.2 Effects of N addition on content of soil P fractions
由图3所示,与CK相比,NaHCO3-Pi占总磷的比例在N5和N10处理下显著提高了49.1%和36.9%(P<0.05);Residual-P占总磷的比例在N10处理下显著降低了18.5%(P<0.05)。此外,不同水平的氮添加处理相较于CK对Resin-Pi,NaOH-Pi,NaOH-Po,1 mol ·L-1HCl-Pi和HCl conc.-Pt所占总磷的比例都没有显著改变。
2.2 不同氮添加水平对土壤生物因素和非生物因素的影响
不同水平的氮添加对土壤ACP活性无显著改变(图4a),ALP活性随氮添加水平的上升有增加的趋势但并不显著。相较于CK,土壤MBC在N10和N15处理下显著提高了12.1%和8.10%(P<0.05,图4c)。土壤MBN在N10和N15处理下均显著增加了17.9%和18.0%(P<0.05,图4d)。


图3 氮添加对土壤磷组分占总磷比例的影响Fig.3 Percentages of different P fractions in total P after N addition

表1 氮添加对土壤理化性质的影响Table 1 Responses of soil physicochemical properties to N addition
图4 氮添加对土壤酸性磷酸酶活性和碱性磷酸酶活性(a)、 微生物生物量碳和微生物生物量氮(b)的影响Fig.4 Effect of N addition on acid phosphatase and alkaline phosphatase activity (a),microbial biomass carbon and microbial biomass nitrogen (b)
2.3 氮添加下植物生物量及地上部整株含磷量的变化
植物地上生物量随着氮添加水平的增加而提高,N10和N15处理相较于CK显著提高97.4%和151%(P<0.05,图5a)。植物地上部磷含量在不同氮添加处理中都有所增加,N10和N15处理相较于CK显著增加了0.07 g·m-2和0.06 g·m-2(P<0.05,图5b)。
2.4 氮添加下影响土壤磷组分的关键因素
由图6所示,土壤MBC和NaOH-Pi含量存在显著正相关关系(P<0.05);MBC和MBN均与Residual-P呈显著负相关关系(P<0.05);土壤pH值和1 mol ·L-1HCl-Pi、HCl conc.-Pt、Residual-P存在显著正相关关系(P<0.05);其他土壤因素和各磷组分含量之间均无显著相关关系。

图5 氮添加对植物地上生物量(a)和植物地上部整株磷含量(b)的影响Fig.5 Effect of N addition on the aboveground biomass of plants (a) and the shoot P content of plants (b)
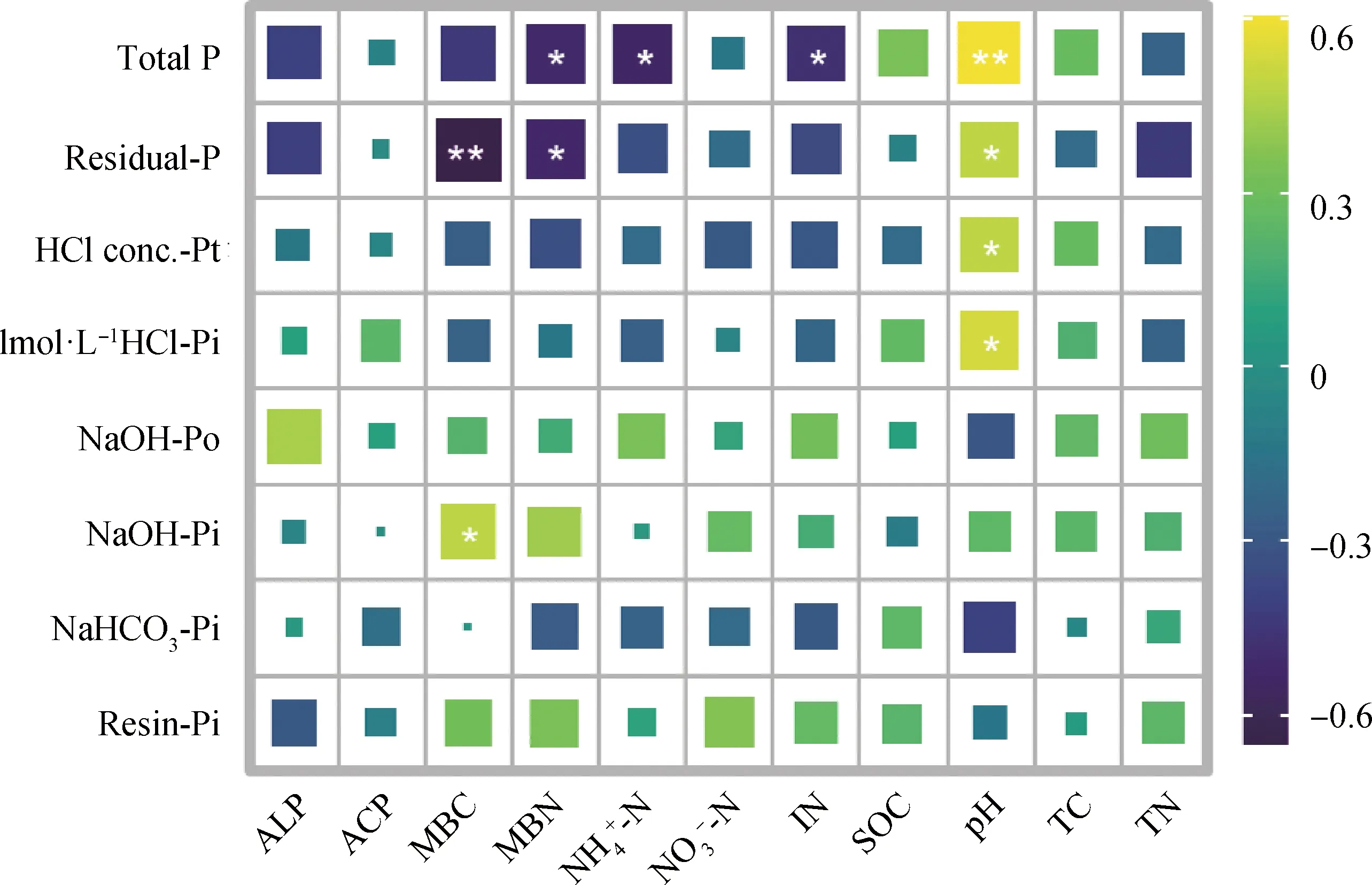
图6 氮添加处理下土壤因素和磷组分的相关性分析Fig.6 The correlations between the soil factors and P fractions after N addition.注:** 表示P<0.01,* 表示P<0.05。ALP,碱性磷酸酶;ACP,酸性磷酸酶;MBC,微生物生物量碳;MBN,微生物生物量氮;IN,无机氮;SOC,土壤有机碳Note:** indicate P<0.01,* indicates P<0.05. ALP,alkaline phosphatase activity,ACP,acid phosphatase activity,MBC,microbial biomass carbon,MBN,microbial biomass nitrogen,IN,inorganic nitrogen,SOC,soil organic carbon
3 讨论
3.1 不同氮添加水平下土壤磷组分状况
N5处理下土壤中有效磷组分的含量和比例增加,缓解土壤中的磷限制。在青藏高原亚高山生态系统的施氮试验结果表明[27],氮施入土壤会通过促进有机磷的周转和释放显著增加有效磷的含量。氮添加会导致土壤发生一系列的变化从而增加植物磷吸收效率,植物会通过产生磷酸酶和释放有机酸(如草酸、柠檬酸和苹果酸)这两种途径进一步促进土壤磷的有效性[28]。在森林生态系统中的试验也得到了相似的结果,氮添加通过降低铁铝氧化物含量减少了难分解和封闭的磷组分,同时增加了土壤的有效磷[29-30]。韩兴国课题组[31]在内蒙古草原长期氮添加的试验结果也表明,氮富集会通过土壤酸化活化草地中矿物结合态的磷,从而增加土壤中有效磷的含量。
本研究中,N15处理下,由于地上生物量和植物含磷量的增加,土壤活性磷的含量和比例在N15处理下均没有显著变化,同时稳定态磷和总磷含量均显著下降。对草地生态系统原位氮添加试验进行的meta分析结果表明,氮添加会显著降低土壤全磷的含量[32]。有草地生态系统的研究得出,土壤全磷含量在两年的氮添加处理下显著下降[33]。这是由于在氮添加条件下地上净初级生产力增加,对土壤有效磷的需求也提高,土壤的中等有效磷库和稳定磷库向有效磷转化并被地上植物吸收利用,这导致了土壤中磷的消耗。也有研究通过相关分析可以得出,土壤磷库含量降低与地上总生物量的积累存在比例关系[28]。在半干旱草原添加尿素的试验结果得出,氮的输入促进了难溶性磷转化为中等有效磷,从而提高了植物对磷的吸收,这归因于较高的植物生产力[29]。
3.2 氮添加下调控土壤磷组分的重要生物因素
本研究采用土壤磷酸酶活性、微生物量碳和氮代表土壤生物因素。其中,土壤磷酸酶根据对环境酸碱度的适应分为酸性磷酸酶(ACP)和碱性磷酸酶(ALP)[35],本研究选取的试验地为石灰性土壤,由ALP主导土壤磷的矿化过程。在不同水平的氮添加下,土壤ALP活性存在上升的趋势,土壤ACP活性没有显著改变。氮添加下土壤磷酸酶活性提高的原因可以归结为土壤微生物在分泌胞外酶时,由于外源氮输入满足了微生物对氮元素的需求,微生物为维护氮磷养分平衡,会将能量集中生产磷酸酶以获取土壤中的有效磷[36]。还有一个原因是,磷酸酶所含氮含量较高,在氮添加条件下,微生物获取足够的氮才能供应磷酸酶的分泌过程[37]。在草原生态系统长期的施氮试验中也得到了相似的结果[38],在氮输入增加的情况下,土壤磷酸酶活性增加。氮沉降可以显著增加温带森林土壤的MBN和MBC含量[39],与本研究结果一致,土壤MBC含量在氮添加处理下显著增加。这与高寒草甸速效养分缺乏有关,由于气温较低,土壤中氮的有效性还是不足以满足植物和微生物的需求,氮的添加为微生物提供了必要的营养元素,因此土壤中的MBC和MBN增加。
氮添加下土壤微生物生物量的增加,这可能有利于微生物固定土壤中的有效磷[40],但同时土壤微生物也可以参与有机磷矿化和无机磷溶解过程[41]。土壤中的微生物也参与调节土壤中磷组分的转化,氮添加下土壤MBN和MBC与Residual-P均存在显著负相关关系,说明微生物也促进了稳定磷向有效磷的转化。此外,氮添加下土壤MBC与NaOH-Pi存在显著正相关关系,说明微生物也在一定程度上促进了稳定磷向中等有效磷的转化。
3.3 氮添加下调控土壤磷组分的关键非生物因素
土壤pH值是衡量土壤状况的重要非生物因素,会受到氮添加水平的影响。本研究结果表明向土壤中施加氮会显著降低土壤的pH值。可能的原因是由于氮在土壤中的硝化作用,硝酸盐的淋溶产生的质子超过盐基阳离子消耗的质子,这会导致土壤酸化[42]。在森林生态系统的研究中,森林生态系统下表层(0~10 cm)土壤pH值在氮添加下也出现了显著降低[29]。还有研究解释是,氮添加提高了微生物生物量,土壤微生物可以产生有机酸,从而降低了土壤pH值[43]。
土壤pH值降低可以减少石灰性土壤中的交换性钙和镁[44],从而促使土壤中的无机磷溶解成为有效磷。相较于土壤有机磷,土壤无机磷的溶解过程受土壤pH值的影响较大[45]。在本研究中,土壤pH值在氮添加下显著降低,并与稳定磷组分呈显著正相关关系。以上结果说明,土壤酸化会增强土壤稳定磷组分向土壤活性磷组分的转化,氮添加可以通过促进土壤难溶性磷组分的溶解而增加土壤磷的有效性。除此以外,土壤pH值还会通过影响土壤微生物进一步影响土壤磷有效性。已知土壤pH值是影响微生物多样性的关键因素,通过改变土壤微生物细胞的pH值稳态来调节土壤养分的有效性[46]。有研究认为土壤中的ALP完全来源于土壤微生物[47],因此,土壤pH值的变化会通过影响微生物的群落结构从而影响土壤中磷酸酶,进而影响土壤磷组分。
4 结论
由于地上植物对磷的吸收,高寒草甸土壤磷组分对不同水平氮添加的响应不一致。土壤pH值是影响青藏高原高寒草甸碱性土壤磷组分周转的关键非生物因素,氮添加通过降低土壤pH值促进稳定磷的溶解,并通过增加土壤微生物生物量促进土壤稳定磷的活化。本研究结果表明,氮添加通过改变土壤pH值和微生物生物量增加了土壤活性磷组分,促进了植物对磷的吸收。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
纺织标准与质量(2022年2期)2022-07-12
作物学报(2022年3期)2022-01-22
煤气与热力(2021年12期)2022-01-19
海外文摘·文学版(2021年3期)2021-05-19
当代化工(2019年3期)2019-12-12
活力(2019年21期)2019-04-01
分析化学(2018年7期)2018-09-17
天津农业科学(2014年12期)2014-12-11
