
从江香猪附睾3β-HSD的年龄依赖性表达模式
2022-11-01 04:20:20刘文娇蒙利洁
福建农林大学学报(自然科学版) 2022年4期
刘文娇, 蒙利洁, 龚 婷
(1.贵州大学高原山地动物遗传育种与繁殖教育部重点试验室;2.贵州省动物遗传育种与繁殖重点试验室;3.贵州大学动物科学学院,贵州 贵阳 550025)
类固醇激素作为哺乳动物体内最主要的性腺激素,包括雌、雄激素等,对动物性器官的发育、成熟具有重要的调控作用,与动物的繁殖性能息息相关[1].3β-羟基甾脱氢酶(3β-hydroxysteroid dehydrogenase, 3β-HSD)是合成类固醇激素所必需的酶,可以将肾上腺前体类固醇转化为孕酮、睾酮等[2].人类有两个3β-HSD家族基因,分别为3β-HSD1基因和3β-HSD2基因,编码3β-HSD1和3β-HSD2两种同工酶,而猪的3β-HSD基因只有一种[3].研究表明:3β-HSD在大鼠的肾上腺、卵巢和睾丸均有表达[4-5].在小鼠饮食中添加高剂量糖精钠,发现睾丸中包括3β-HSD在内的睾酮合成相关酶表达量与对照组及中、低剂量处理组相比极显著降低,且高剂量处理组小鼠的精子活力、精子密度有所下降,畸形精子数量增多[6];在大鼠退化的附睾中注射睾酮,发现附睾小管直径增大、附睾上皮高度增加、上皮细胞中的主细胞数量增加[7].因此,推测3β-HSD作为睾酮合成过程的关键酶不仅在睾丸中发挥作用,也可能影响附睾的形态结构、生理作用,从而影响雄性生殖生理活动.
本课题组前期对附睾发育进行了研究,将从江香猪的附睾发育分为4个时期,即初情期前(15 d)、初情期(30 d)、初情期后(60 d)、性成熟期(180 d)[8-9].哺乳动物精子的发生过程在睾丸中完成,随后在附睾运行的过程中受到附睾上皮细胞及平滑肌细胞合成和分泌各种因子的相互作用,经过附睾腔特殊的动态微环境后促使精子成熟[10].睾酮作为睾丸和附睾中最重要的一种雄激素,其合成受到多种酶的调节,如类固醇激素合成急性调节蛋白(StAR)、细胞色素P450侧链裂解酶(CYP11A1)、3β-HSD、17β-羟基类固醇脱氢酶(17β-HSD)等的调节[11].本课题组前期研究表明,雄激素受体在不同日龄从江香猪的附睾中均有表达,这提示与雄激素相关的酶可影响附睾对精子成熟、运输、储存等的调控功能[12].另有研究表明,3β-HSD在胎盘的巨大滋养层细胞中存在表达,表明3β-HSD参与孕酮的合成,且孕酮是促进雌性第二性征发育成熟及维持妊娠所必需的[13].胆汁酸在脂肪代谢中起着不可替代的作用,肝脏中3β-HSD的缺乏会影响胆汁酸代谢,引起血清中维生素水平的下降,最终导致胆道淤积、肝脏肿大[14].3β-HSD在睾丸中的表达[15]已有文献证实,但对该酶是否存在于附睾中,并直接影响附睾生理功能的研究较为缺乏.
从江香猪是我国一种重要的微型地方猪种,具有体型矮小、性成熟早、基因纯合、肉质鲜嫩等特点,目前已被列为国家二级珍稀保护畜种.然而,该猪种与大白猪等猪种相比繁殖力较低,严重影响了其扩繁推广.对从江香猪繁殖性能开展相关机制的研究,对提高该猪种公猪的种用价值有重大意义.因此,本试验选取不同日龄从江香猪公猪作为研究对象,在蛋白水平上探索3β-HSD在附睾中的表达模式,旨在为探究3β-HSD对从江香猪精子生成调控的潜在分子机制及提高从江香猪的生产性能提供参考.
1 材料与方法
1.1 材料
本试验严格遵循贵州大学动物试验伦理(批准号:1801227).所用动物为出生15(初情期前,n=3)、30(初情期,n=3)、60(初情期后,n=3)、180 d(性成熟期,n=3)的健康雄性从江香猪共12头,购于贵阳绿生源畜牧科技发展有限公司.
1.2 样品采集
通过手术法获取附睾、睾丸组织,其中一侧附睾、睾丸组织在4%多聚甲醛固定液中固定后做连续切片两张,每日龄得到12张切片用于免疫组织化学检测;剩余一侧附睾、睾丸组织在采集后立即剪碎置于液氮中保存,用于后续Western blot分析.
1.3 附睾组织石蜡切片制作

1.4 免疫组织化学检测
1.5 SDS-PAGE与Western blot分析
SDS-PAGE与Western blot分析参照蒙利洁等[12]的方法进行.在剪碎的附睾中加入含1%苯甲基黄酰氟(PMSF)的RIPA裂解液(北京索莱宝生物科技有限公司)提取总蛋白.蛋白浓度测定及蛋白定量分析参照BCA试剂盒(北京索莱宝生物科技有限公司)说明书进行,随后定量、变性蛋白质.SDS-PAGE电泳后切胶,将蛋白转移至用甲醇浸泡2 min的聚偏二氟乙烯膜(PVDF)上,使用恒定电流100 mA转膜2 h.将膜浸入5%脱脂奶粉中于37 ℃摇床封闭3 h,加入一抗[3β-HSD(购自赛默飞世尔科技有限公司,货号:sc374244,稀释比例:1∶1 000)、β-actin(购自江苏亲科生物研究中心有限公司,货号:AF7018,稀释比例:1∶5 000)于4 ℃摇床反应过夜.洗膜3次,每次10 min,随后加入HRP标记的二抗(1∶5 000)于37 ℃摇床孵育2 h.洗膜3次,每次10 min,黑暗条件下使用ECL超敏发光进行显色,使用ChemiDocXRS系统成像、拍照,条带灰度用Image J软件分析.
1.6 数据处理
数据以“平均数±标准差”表示.3β-HSD灰度值采用Image J软件测定,免疫组织化学检测结果用正置显微镜进行观察、拍照.使用GraphPad Prism 6.0软件进行单因素方差分析,使用Dunnett法进行多重比较.其中,P<0.01表示差异极显著,用**表示.
2 结果与分析
2.1 3β-HSD在不同日龄从江香猪附睾中的表达定位
不同日龄从江香猪附睾3β-HSD免疫组织化学检测结果(图1)显示,从江香猪附睾在初情期前(15 d)、初情期(30 d)、初情期后(60 d)、性成熟期(180 d)均有3β-HSD的表达.3β-HSD在各日龄段从江香猪附睾平滑肌中均有表达,其中,15 d的3β-HSD特异性表达于附睾液及脱落的生精细胞中,30 d的3β-HSD少量表达于基细胞中,60 d的基细胞和微绒毛被3β-HSD深染,180 d可在精子中观察到3β-HSD的表达.
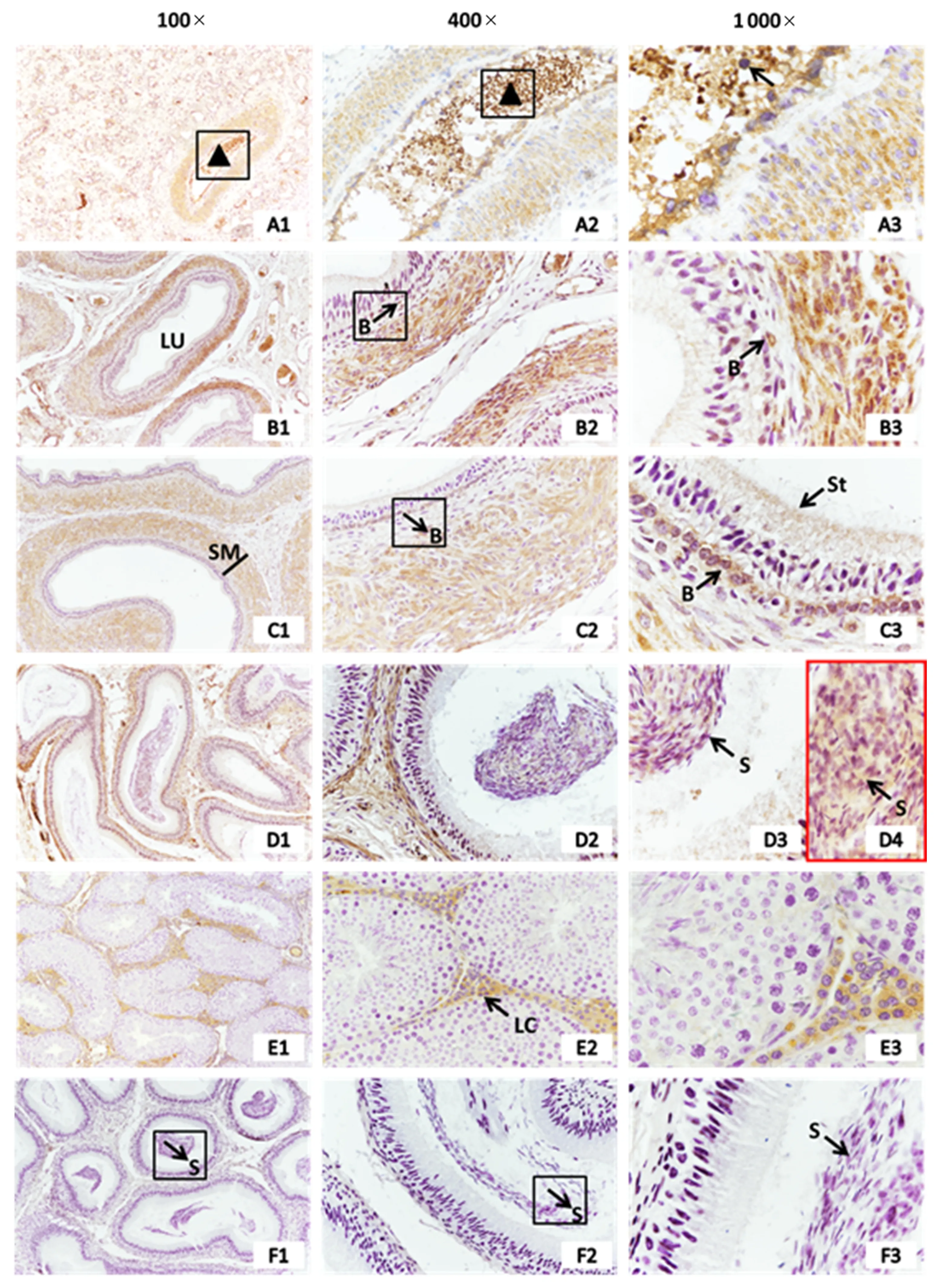
A1~A3、B1~B3、C1~C3、D1~D4分别为初情期前、初情期、初情期后、性成熟期的附睾;E1~E3:性成熟期的睾丸(阳性对照);F1~F3:性成熟期的附睾(阴性对照,用PBS代替3β-HSD一抗)(LU:管腔;SM:平滑肌;S:精子;B:基细胞;St:微绒毛;LC:间质细胞;A1、A2中的三角形表示附睾液;A3中的箭头表示脱落的生殖细胞).图1 从江香猪附睾3β-HSD免疫组织化学检测结果Fig.1 Immunohistochemistry detection of 3β-HSD in epididymis of Congjiang Xiang pig
2.2 从江香猪附睾3β-HSD的表达
以β-actin为内参蛋白,从江香猪附睾3β-HSD表达的Western blot检测结果(图2)显示,3β-HSD表达于从江香猪初情期前、初情期、初情期后、性成熟期4个发育时期的附睾中,且附睾3β-HSD蛋白的分子质量约为43 ku.3β-HSD表达灰度值在30 d显著增加,并达到最大值,表达丰度表现为:初情期>性成熟期、初情期后、初情期前,此结果与免疫组织化学染色结果一致.

A:3β-HSD抗体的特异性检验;B:3β-HSD在15、30、60和180 d 附睾组织中的表达量[**表示差异极显著(P<0.01)];C:Western blot检测结果.图2 从江香猪附睾3β-HSD表达的Western blot检测结果Fig.2 The expression of 3β-HSD protein in epididymis detected by Western blotting
3 讨论与结论
从江香猪的附睾形态与其他哺乳动物相似,由假复层柱状上皮和周围的平滑肌组成[16].研究证实,雄激素和一些管腔因子(如类β-半乳糖苷酶、脂质转运蛋白、人类附睾蛋白6/G蛋白耦联受体64)在附睾的发育过程中起着必要的作用.如:β-半乳糖苷酶与附睾中的精子成熟机制有关[17];LCN5、LCN6、LCN8是脂质转运蛋白家族的重要成员,其发挥作用时可与精子成熟过程中精子膜脂类的转运相联系[18];人类附睾蛋白6/G蛋白耦联受体64具有调节附睾水平的作用[18-19].但目前对上述因子介导的精子运输、成熟及附睾形态变化的分子机制仍知之甚少.本试验结果显示,从江香猪附睾中3β-HSD的表达具有年龄依赖性,且在初情期的表达量最高.初情期前、初情期后、性成熟期3β-HSD的表达量相似,均极显著低于初情期,这可能与附睾自身独特的微环境有关.精子成熟的过程依赖于附睾高度专业化的管腔内微环境,免疫组织化学检测结果显示,从江香猪初情期的附睾上皮细胞类型已分化完成,细胞形态多为立方柱状.初情期后附睾上皮细胞向基底层移动,此时细胞多为窄高柱状[20].暗示从江香猪初情期附睾3β-HSD的高表达可能参与了附睾精子运输、成熟及附睾发育的调控.因此,附睾中的3β-HSD可能会影响精子的运输、成熟及雄性性器官的发育,从而形成有助于精子成熟和储存的环境.
附睾与睾丸的连接通过睾丸输出小管介导,成熟哺乳动物中每秒有1 000~1 500个精子进入附睾,在长达1、2、6 m(分别为小鼠、大鼠、人)的附睾小管中运行,最终到达附睾尾部储存[21].Mewe et al[22]研究发现,在去除附睾平滑肌后,附睾的收缩完全停止,表明精子在附睾中的运行离不开附睾平滑肌、管腔液体、微绒毛的相互作用.本试验结果显示,3β-HSD主要表达在从江香猪附睾的平滑肌中,并且在各发育时期均有表达.3β-HSD在初情期前表达于附睾上皮的主细胞中.主细胞占附睾上皮细胞的80%,负责分泌大部分附睾管腔内的蛋白因子.因此推测附睾主细胞可能通过分泌3β-HSD对附睾微环境或精子成熟产生影响.研究表明,雄激素对附睾的结构和功能具有重要的调控作用[23-24].雌、雄激素受体在附睾平滑肌中均有表达,表明雌、雄激素对精子的运输和成熟具有调控作用.本课题组前期证实了3β-HSD在从江香猪附睾平滑肌中存在表达,表明附睾平滑肌可能是雄激素合成关键酶的靶器官[25].初情期的附睾上皮细胞已经发育完全,此时的3β-HSD在附睾上皮周围的平滑肌和少数基细胞中呈阳性表达.初情期时,从江香猪初次产生具备受精能力的精子,此时的3β-HSD在附睾中高表达,可能与精子的成熟、运输机制有关;初情期后的3β-HSD定位于附睾平滑肌、微绒毛和基细胞中.初情期后开始产生大量精子,微绒毛可以吸收管腔物质及分泌更多的蛋白质,这种结构有利于精子储存,此时附睾平滑肌和微绒毛中3β-HSD的阳性表达对于精子的运输、成熟和储存是必要的[26-27].基细胞具有改变形状以调整管腔容积和压力的能力,其在保护管腔结构完整性的方面起到了重要作用,从而对精子的储存和运输产生影响[28].此外,性成熟期的3β-HSD主要在附睾平滑肌中呈阳性表达,在精子中也有表达.附睾管中的因子和精子会影响平滑肌的收缩.Mewe et al[22]在去除牛附睾管中的精子后,平滑肌收缩加快,这或许可以解释本试验初情期后从江香猪附睾中3β-HSD表达量的下降.总体上,附睾各种特征都与其特定的功能有关[29-30].3β-HSD在附睾平滑肌中的表达可能对精子在附睾内的运输进行调节,在微绒毛中的表达对精子运输、成熟也起调节作用,在基细胞中的表达可能通过改变附睾容积从而影响精子储存.
3β-HSD参与精子的发生及成熟、获能、储存和受精等一系列过程[31-32].本试验结果显示,附睾中3β-HSD的表达在初情期达到峰值.附睾中的3β-HSD可能与从江香猪初情期初次释放精子、附睾迅速发育、附睾功能基本完善等方面有关.进入附睾后,精子的结构和功能进一步完善,需要多种因素相互作用,初情期时附睾上皮细胞中3β-HSD的表达量最高,提示附睾中的3β-HSD对精子运输、成熟及储存发挥重要作用.
猜你喜欢
房地产导刊(2020年9期)2020-10-28 08:37:24
中国果树(2020年2期)2020-07-25 02:14:18
———从江加车村
原生态民族文化学刊(2018年4期)2019-01-16 01:26:22
现代营销(创富信息版)(2018年6期)2018-09-05 03:14:38
浙江柑橘(2016年4期)2016-03-11 20:13:01
西南军医(2016年6期)2016-01-23 02:21:14
乡村地理(2015年3期)2015-11-10 06:32:34
现代检验医学杂志(2015年6期)2015-02-06 01:44:11
西南军医(2015年4期)2015-01-23 01:19:02
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
