
福建莆田海域指沟卡尔藻赤潮海洋监测浮标数据变化及成因分析
2022-10-28 01:23李青俞
渔业研究 2022年5期
李青俞
(莆田市海洋与渔业环境监测站,福建 莆田 351100)
赤潮是由一些海洋微藻、原生动物或者细菌在水体中暴发性繁殖或者聚集而造成的一类典型的生态灾害。赤潮的发生是一个综合了水文气象、物理、化学、生物等一系列因素的复杂过程[1-2]。当赤潮发生时,赤潮生物迅速消耗氮、磷等营养盐,合成有机物,夜间因其呼吸作用而大量消耗溶解氧,并且在赤潮衰败时,赤潮生物分解也会消耗大量的溶解氧,从而造成水体缺氧。如果是由有毒生物引发的赤潮,其毒素将杀死养殖生物或者在养殖生物体内富集,随着水产品被人摄食而进入人体,影响人的健康甚至危及生命。赤潮的种类包括所有能改变海水颜色的有毒藻或无毒藻引发的赤潮,以及那些虽然生物量低、不能改变海水颜色,但因含有藻毒素而具有危害性的藻华[2]。
南日岛位于福建省莆田市东南部兴化湾和平海湾交汇处,东涉台湾海峡,盛产鲍、大黄鱼、蟹、红毛藻等多种水产品。最近20年,南日岛周边海域是赤潮高发区,也是损失的重灾区[3];未来5年,南日岛所处的福清市南部—莆田东北部近岸海域的赤潮灾害危险度为高危险区[4],因此莆田南日岛海域赤潮防灾减灾需要受到极大关注。
指沟卡尔藻(Karlodiniumdigitatum)是一种有毒的海洋甲藻,曾被鉴定为指沟凯伦藻(Kareniadigitata),隶属裸甲藻目(Gymnodiniales)凯伦藻科(Kareniaceae)卡尔藻属(Karlodinium)。指沟卡尔藻曾于1995年、1996年夏季及1997年11月在日本沿海形成过赤潮,造成鱼类死亡[5];1989年首次在中国香港海域出现,1995年再次出现,但直到1998年才形成赤潮。在1998年我国香港海域暴发的特大型凯伦藻赤潮中,指沟卡尔藻作为新种被首次报道[6-7];2019年5月福建平潭发生大规模米氏凯伦藻赤潮,后被证实肇事种是指沟卡尔藻[8],这是我国目前仅有的两起被报道的指沟卡尔藻赤潮,均导致大量鱼类死亡和重大经济损失。
海洋监测浮标(以下简称浮标)具备实现各类天气条件下连续、定时、定点24 h不间断监测的特性,可实时地自动监测海洋环境要素的变化,实现对赤潮全过程的跟踪监测,弥补了人工监测的不足,对于研究赤潮机制和快速预警预报具有重要意义[9-16]。利用浮标监测赤潮最大的不足是其不能监测赤潮生物的种类和密度。莆田海域浮标建设依托国家2015年海湾(海岸带)整治修复项目,于2017年正式投入使用,目前共有4套。莆田海域浮标运行了近6年,积累了海量的数据,并被多次应用到莆田海域的赤潮防灾减灾工作中,对赤潮的监测发挥了重要的作用。
本文主要根据2022年莆田南日岛海域发生的指沟卡尔藻赤潮过程中浮标气象、水质及营养盐数据,并结合现场人工监测结果进行分析,初步探讨本次指沟卡尔藻赤潮暴发的原因,以期为赤潮研究提供资料参考。
1 材料和方法
1.1 浮标监测
浮标布设在南日岛坑口海域的鸟屿浮标(25.256 8°N、119.489 2°E)、港南海域的港南浮标(25.156 8°N、119.472 6°E)、筶杯岛海域的筶杯岛浮标(25.339 6°N、119.272 7°E),浮标布设位置见图1。由于本次赤潮发生在南日岛附近海域(图1红圈所示海域),因此本文使用布设在南日岛坑口浮标和港南浮标的数据进行分析,而筶杯岛浮标数据则用于参照。
浮标主要搭载美国YSI水质监测仪、芬兰VAISALA分体式气象监测仪及斯坦道OSTD-NutriS原位营养盐分析仪(坑口海域的鸟屿浮标未搭载营养盐分析仪)等设备,主要监测的要素有风速、风向、气压、相对湿度、气温、溶解氧(DO)、电导率、盐度、水温、pH值、叶绿素a(Chl-a)、浊度、氨氮、亚硝酸盐氮、硝酸盐氮、磷酸盐。浮标由厦门斯坦道科学仪器有限公司运维团队定期运维(4—6月每半月1次,其他月份每月1次)、校对保证浮标系统的正常运行。所使用的参数均定期与计量认证的通用方法比对,确保浮标监测数据的真实可靠性。浮标水质探头布设在水下1.0 m处,气象探头布设在水面1.5 m高度。水质和气象每隔30 min采集一组数据,营养盐数据每隔4 h采集一次。浮标采集的数据通过双北斗卫星通信系统实时发送至数据接收服务器。
自然界中浮游植物叶绿素以叶绿素a占绝大多数,在发生赤潮时,水体中叶绿素a含量通常超过10.0 μg/L[1]。在浮标用于赤潮监测和研究中,一般以叶绿素a含量达到或超过10.00 μg/L即认为出现赤潮[14,16-17]。因此本文以海域中叶绿素a大于10.00 μg/L且赤潮生物超过或接近赤潮基准浓度认为该海域中存在赤潮。
1.2 人工监测
赤潮现场人工监测依据《海洋监测规范》[18-19]相关规定开展。浮游植物样品按每L水加6~8 mL鲁哥氏碘液固定后带回实验室,在莱卡倒置显微镜下鉴定和显微计数,确定赤潮优势种及细胞密度。每份水样计数3个分样,取平均值。
2 结果与分析
2.1 赤潮过程描述
6月1日南日岛坑口海域检出指沟卡尔藻,密度为1.02×106cell/L。在光镜下,该藻为细小单一细胞,总体呈现球状或卵状,细胞长度为10~26 μm,宽度为10~22 μm;上壳半球形或宽的圆锥形,下壳半球形,底部无凹陷。按照《赤潮监测技术规程》[20]相关规定,该藻赤潮生物细胞密度基准浓度为1.0×106cell/L,故判定南日岛海域已经发生赤潮。在本次赤潮过程中,大部分时间赤潮海域水色正常,仅在部分时间(6月3日和6月7日)部分海域(坑口海域)可见到不连续黑褐色、油膜状片状水团,有分层现象。赤潮造成了网箱养殖的鲍鱼、大黄鱼、鲈鱼、红鱼、真鲷、黑鲷等出现不同程度的死亡。本次赤潮期间共采集赤潮海域10份贝类样品,并检测其麻痹性贝毒和腹泻性贝毒含量,结果均未检出。人工监测结果见表1。
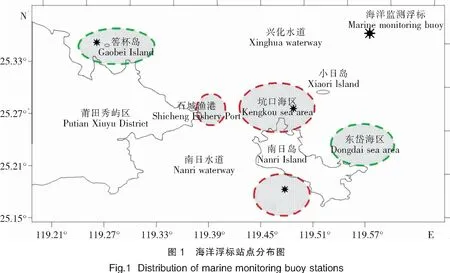
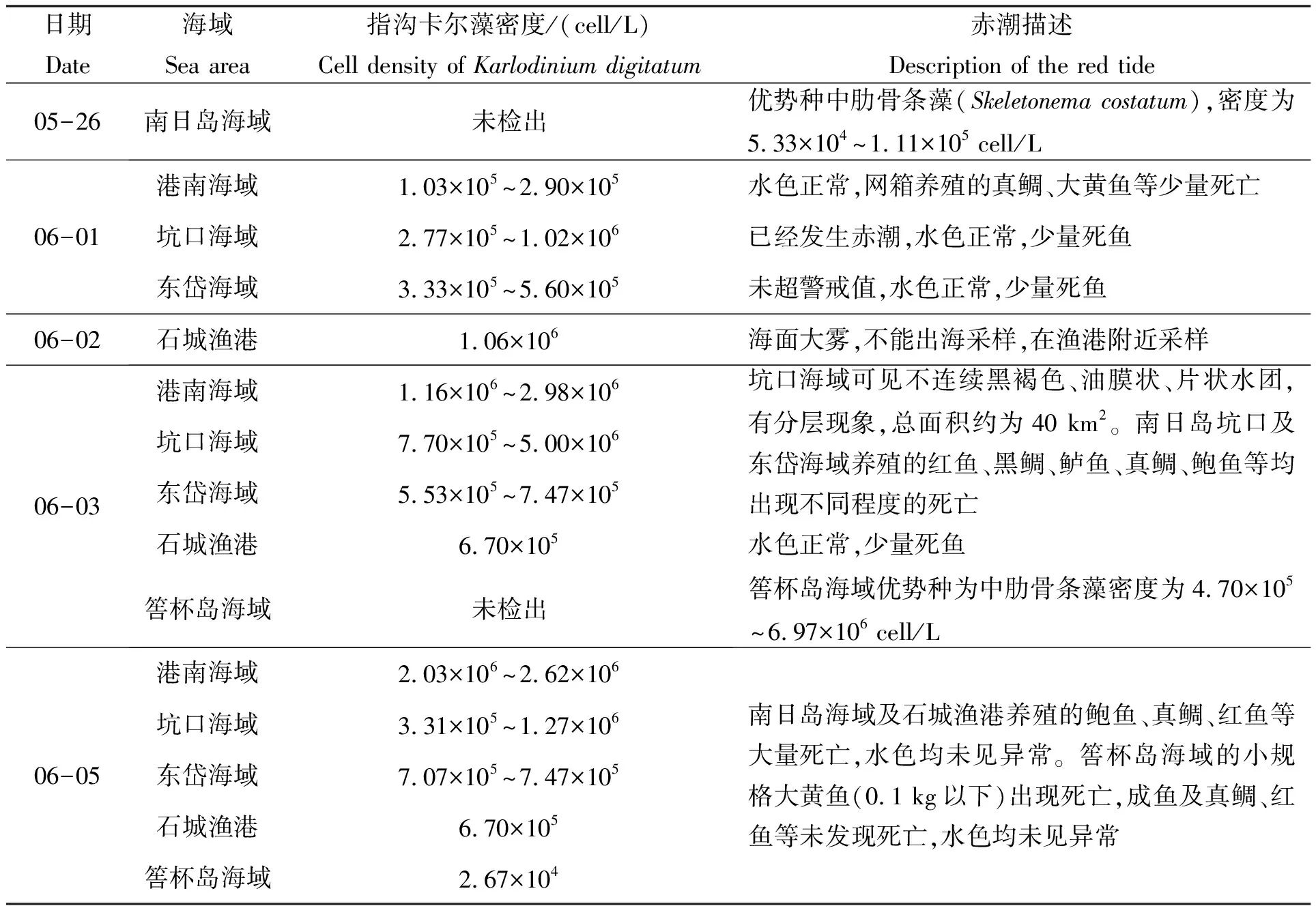
表1 本次赤潮过程人工监测结果
2.2 赤潮过程浮标数据变化及分析
为便于分析浮标数据,结合现场监测数据及浮标监测数据,将本次赤潮过程分为以下3个过程:赤潮暴发前(5月26日至31日)未检出指沟卡尔藻,部分监测指标出现异常;赤潮发展及维持阶段(6月1日至8日),此过程叶绿素a超过警戒值10.00 μg/L,指沟卡尔藻密度超过或接近赤潮基准浓度(1.0×106cell/L),养殖生物死亡;赤潮消亡阶段(6月9日至13日),叶绿素a低于10.00 μg/L,指沟卡尔藻密度远低于赤潮基准浓度,未出现养殖生物死亡。
2.2.1 气象数据
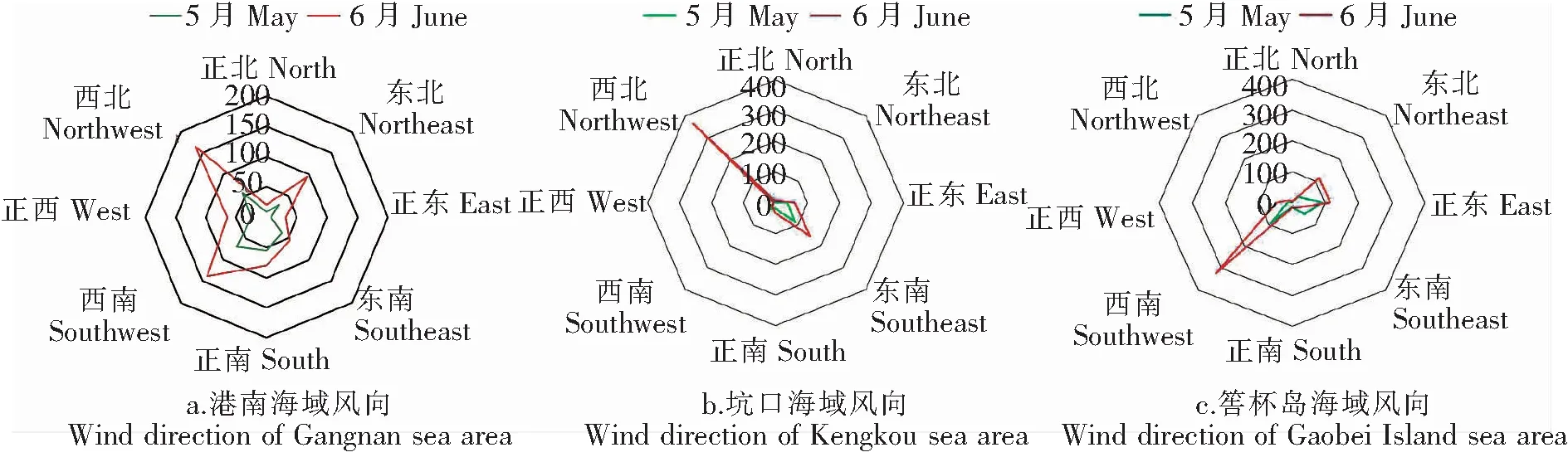
如表2所示,赤潮暴发前坑口及港南海域天气以阴天为主,平均气温逐渐升高,6月6日至9日莆田普降大到暴雨,各海域气温均降低,随后各海域气温均不同程度回升。如图2(a、b、c)风向玫瑰图显示,在整个赤潮过程中,港南海域以西北风为主,坑口海域以西北风为主,均有利于赤潮生物向岸边聚集;筶杯岛海域以西南风为主,不利于赤潮生物向岸边聚集。如图2(d)所示,整体上筶杯岛海域风速高于南日岛海域。赤潮暴发前坑口及港南海域日最大平均风速不超过5.0 m/s,存在2个低风速时间段,分别是5月28日及5月31日。在赤潮发展及维持阶段,天气仍以阴天为主,平均风速有逐渐增大趋势,日最大平均风速达8.0 m/s。
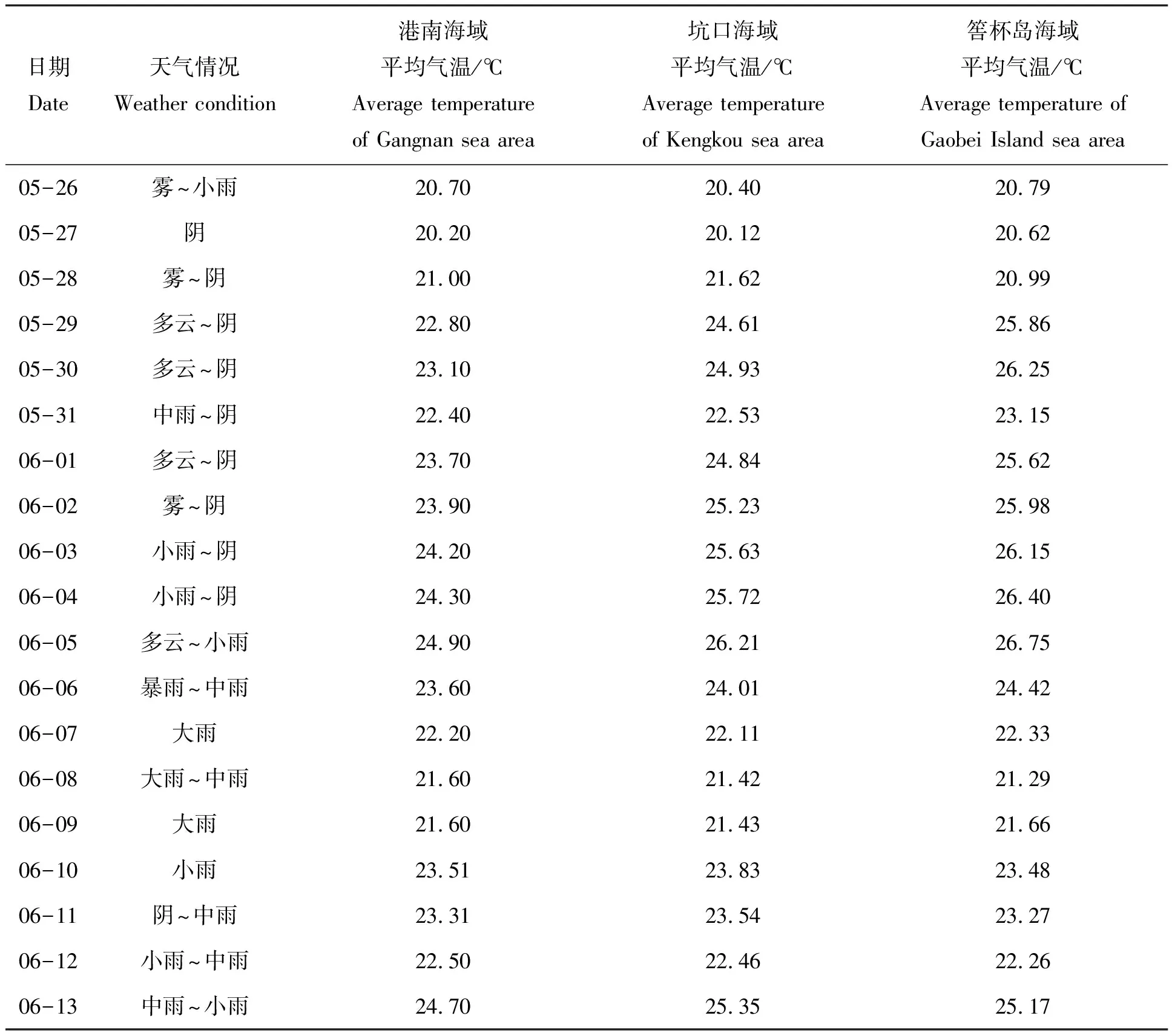
表2 气象信息
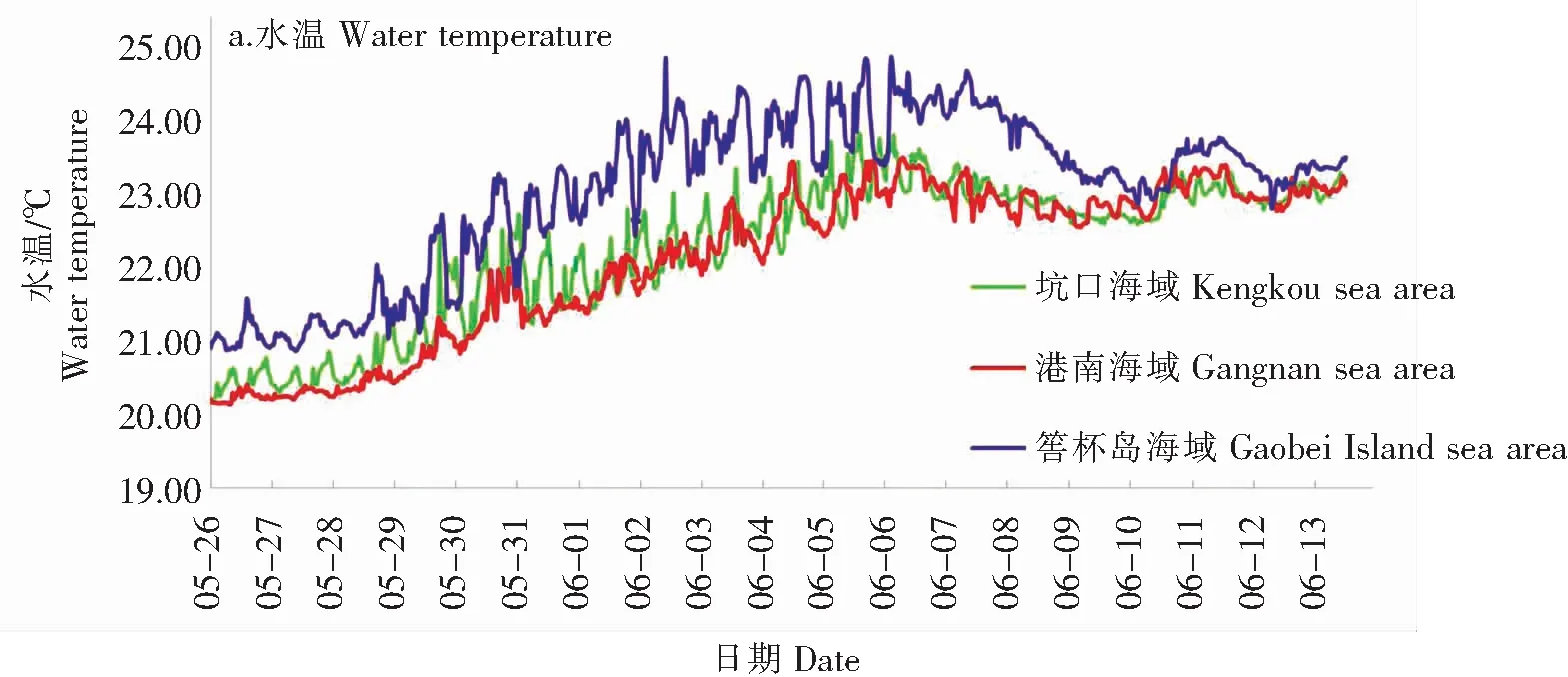
2.2.2 水温、盐度
水温如图3(a)所示,5月26日至6月6日,港南、坑口和筶杯岛浮标所在海域水温均呈现快速升高趋势,港南海域水温由20.15℃快速上升到23.52℃;坑口海域水温由20.20℃快速上升到23.85℃;筶杯岛海域水温由20.88℃升高至24.89℃。南日岛海域最高叶绿素a含量出现在水温22.29~23.18℃之间。6月6日至10日,水温有一个降低过程,10日至13日水温略有回升。
盐度如图3(b)所示,赤潮暴发前,港南海域盐度在30.00~30.50之间小幅度波动;坑口海域盐度有逐渐升高的趋势,由29.80升高至31.23;筶杯岛海域盐度因受到木兰溪径流影响而盐度偏低,在24.44~29.63之间波动,平均值为27.81;各海域盐度日内均呈现窄幅波动。在赤潮发展及维持阶段,港南及坑口海域海水盐度总体呈现日内宽幅波动的趋势,最高叶绿素a含量出现在盐度30.28~30.80。6月6日至13日,港南海域盐度在30.20~32.89之间宽幅波动;坑口海域盐度在26.87~31.69之间宽幅波动;筶杯岛海域盐度出现一个明显下降的过程,盐度由29.63降至最低10.40,平均值为23.23。
6月6日至13日,南日岛海域及筶杯岛海域水温降低和盐度宽幅波动与此段时间内莆田市出现的明显降水过程有关。盐度变化除受到潮汐影响外,还极易受到地表径流的影响。大量降水导致地表径流加大,随着淡水大量注入海域,导致各海域盐度宽幅波动,筶杯岛海域尤为显著。
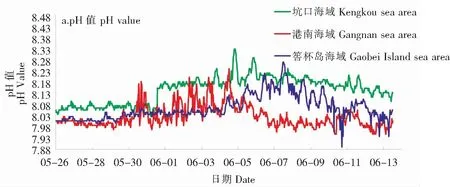
2.2.3 pH、溶解氧、叶绿素a
港南海域、坑口海域和筶杯岛海域pH、溶解氧、叶绿素a数据分析如图4所示。
1)港南海域
赤潮暴发前,港南海域pH值(7.97~8.06)、溶解氧(7.15~7.93 mg/L)、叶绿素a(1.10~5.33 μg/L)窄幅波动。pH值、溶解氧和叶绿素a在5月29日下午17:30(pH值8.05、溶解氧10.65 mg/L、叶绿素a17.64 μg/L)及5月30日13:30(pH值8.19、溶解氧8.43 mg/L、叶绿素a28.48 μg/L)均同时出现阶段性峰值。
2)坑口海域
赤潮暴发前,坑口海域叶绿素a含量在1.00~5.46 μg/L之间波动;虽然6月1日在坑口码头附近海域已检出密度为1.02×106cell/L的指沟卡尔藻,其已超过该藻的赤潮基准值,但浮标数据显示,整个海域叶绿素a含量相对赤潮暴发前峰值只是略有升高,直至6月3日才出现高于10.00 μg/L的峰值;6月3日前,叶绿素a的峰值均短暂出现在高潮后0.5 h内或低潮前0.5 h内;6月3日至8日,叶绿素a值表现为每日3~5个峰值,其他时间则较低。赤潮暴发前pH值在8.03~8.10之间窄幅波动;5月31日中午出现pH值异常增高,并一直维持在高位,在8.15~8.34之间波动,日内pH值波动变大直至赤潮消亡。赤潮暴发前,溶解氧浓度在6.69~7.69 mg/L之间窄幅波动;赤潮暴发期溶解氧浓度在6.63~9.77 mg/L之间宽幅波动,每日1个峰值,且峰值逐渐变大,至4日达到最高峰值9.77 mg/L,此后峰值逐渐降低直至赤潮消亡;赤潮消亡后溶解氧浓度呈现窄幅波动且整体含量低于赤潮暴发前。
现场人工监测数据显示,赤潮初期坑口海域仅靠近岸边的站位的指沟卡尔藻的密度超过赤潮基准值,而坑口海域其他区域指沟卡尔藻密度并未超过赤潮基准值,密度在2.77×105~7.70×105cell/L之间。浮标(浮标距离海岸2 km左右)每日在高潮后0.5 h和低潮前0.5 h可短暂监测到叶绿素a峰值,其他时段则较低。随着赤潮的扩散及面积增加,在指沟卡尔藻细胞群体暴发性增殖叠加高度聚集作用下,整个海域的叶绿素a含量才开始整体升高,此时浮标1 d内可在不同时段监测到多个峰值,叶绿素a含量整体较前一阶段高。据此推测,在坑口海域指沟卡尔藻赤潮初期,近岸水域高密度指沟卡尔藻是因藻类细胞高度聚集而导致的。
3)筶杯岛海域
5月26日至6月4日,筶杯岛海域pH值(8.00~8.08)、溶解氧(6.78~7.76 mg/L)和叶绿素a(1.21~10.65 μg/L)在各自“箱体”内窄幅波动,每日出现1~2个峰值,其中叶绿素a在5月31日至6月1日凌晨,形成阶段峰值,最高值达10.65 μg/L,这或与水体中较高密度的中肋骨条藻有关(表1)。筶杯岛海域6月5日检出指沟卡尔藻。6月5日至9日,筶杯岛海域pH值、溶解氧、叶绿素a均突破前期波动“箱体”,日内宽幅波动,在6月7日达到最高值,分别达到8.28、9.24 mg/L和23.5 μg/L,9日后逐日降低。
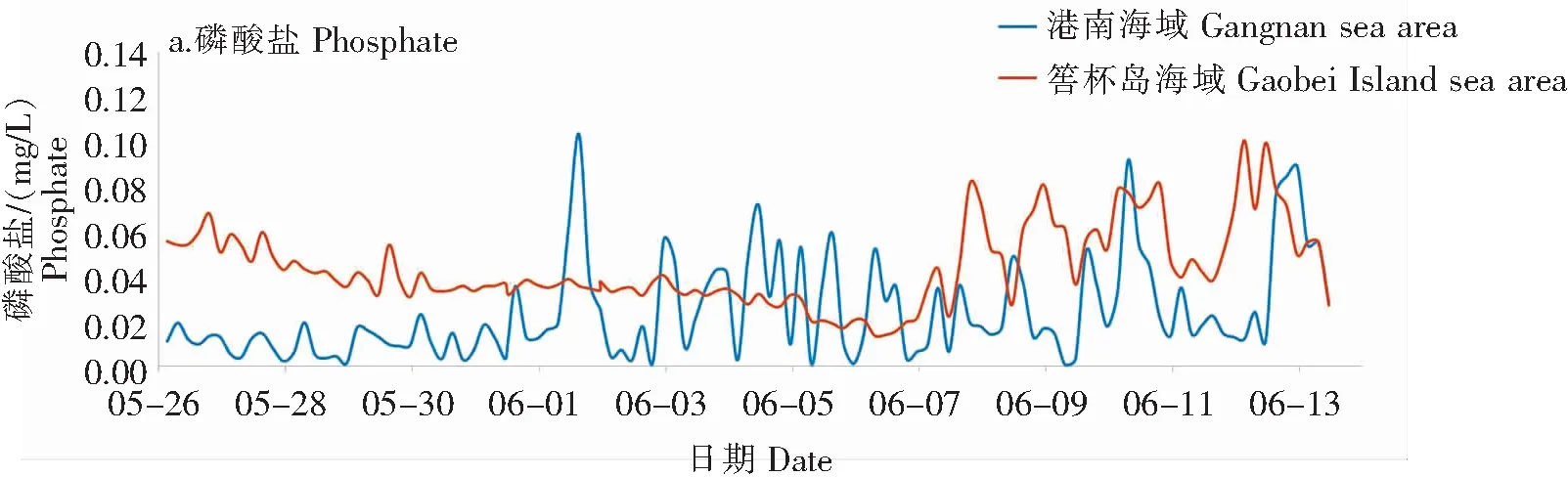
2.2.4 主要营养盐
港南海域和筶杯岛海域营养盐数据分析如图5所示。
1)港南海域
赤潮暴发前,港南海域磷酸盐含量在0.001 2~0.023 mg/L之间,平均值为0.011 mg/L,磷酸盐含量较低,参考《海水水质标准》(GB 3097—1997)[21]属于一、二类海水;氨氮含量为0.001 3~0.043 mg/L,平均值为0.001 8 mg/L,溶解无机氮(DIN)含量为0.029~0.35 mg/L,属于一、二类海水;氮、磷含量均呈现日内窄幅波动的趋势。氮磷比(N/P)值在1.47~110.90之间,平均值为17.14,高于正常海水(N/P值为16)[22-23],尤其是在5月31日,由于大量氨氮的汇入,该海域N/P值最高达110.90。因此,赤潮暴发前,港南海域属于低磷和高N/P的海域。
5月31日,南日岛海域中雨,港南海域磷酸盐及氨氮含量显著上升,磷酸盐含量于6月1日达到阶段峰值,最高值达0.10 mg/L,是赤潮暴发前平均值的9倍。6月1日至赤潮消亡,磷酸盐含量在0.000 6~0.010 mg/L,平均值为0.030 mg/L,呈现日内1~2个峰值宽幅波动的趋势,大部分时间高于海水三类水标准。氨氮浓度也显著上升并在5月31日达到峰值,最高值达0.24 mg/L,6月1日出现次高峰,最高值为0.12 mg/L。6月2日至赤潮消亡,氨氮含量与赤潮暴发前相当。溶解无机氮含量在0.019~0.17 mg/L之间,N/P值在0.34~171.60之间,平均值为9.74。此阶段,港南海域属于富营养水质和低N/P的海域。
5月26日至6月13日,亚硝酸盐氮含量为0.000 6~0.020 mg/L,平均值为0.010 mg/L;硝酸盐氮含量为0.001 2~0.13 mg/L,平均值为0.057 mg/L。
2)筶杯岛海域
5月26日至6月6日,筶杯岛海域营养盐含量总体呈现下降趋势,日内窄幅波动。磷酸盐含量在0.014~0.006 8 mg/L之间,平均值为0.036 mg/L,属于三、四类海水;氨氮含量为0.002 3~0.096 mg/L,平均值为0.005 4 mg/L;硝酸盐氮含量在0.20~0.72 mg/L之间,平均值为0.428 mg/L;亚硝酸盐氮含量在0.017~0.046 mg/L之间,平均值为0.039 mg/L,溶解无机氮含量在0.25~0.86 mg/L之间,平均值为0.52 mg/L;N/P值在9.49~18.86之间,平均值为14.41。
6月7至13日,筶杯岛海域营养盐含量总体急剧升高,日内宽幅波动。溶解无机氮含量在0.35~1.43 mg/L之间,N/P值在7.77~21.67之间,平均值为14.33。
5月26日至6月13日,筶杯岛海域都属于高营养盐和低N/P的海域。总体而言,该海域营养盐含量高于港南海域。
3 讨论
3.1 气象因素对赤潮的影响
赤潮暴发前,莆田市海域无明显天气过程影响,风速和风浪较小。各海域以阴天为主,属于弱光照条件。当水体中硅藻和甲藻并存时,在弱光照下,甲藻的竞争力较强,而在高光照下,硅藻的竞争力较强[24-25]。赤潮发生前后坑口海域平均风速为1.18 m/s,港南海域平均风速为2.26 m/s。弱光照和低风速为赤潮暴发前指沟卡尔藻的强竞争力提供了保证。港南海域以西北风为主,港南海岸线为西北东南走向,坑口海域以西北风为主,坑口海岸线为东北西南走向,均有利于赤潮生物向沿岸海域聚集。
3.2 水温、盐度对赤潮的影响
水温被认为是暴发赤潮较为重要的环境因子之一,水温的突然升高和降雨后盐度的降低容易刺激赤潮生物的生长而引发赤潮[26-27]。但本文研究发现,本次指沟卡尔藻赤潮暴发前、赤潮发生及维持阶段水温和盐度均是持续升高的,这和陈国斌研究厦门同安湾海域血红哈卡藻赤潮的结果[9]一致。南日岛海域赤潮发展及维持阶段水温在20.15~23.86℃之间,南日岛海域最高叶绿素a含量出现在水温22.29~23.18℃,推测指沟卡尔藻最适水温约为22~23℃。
盐度通过影响浮游植物的渗透压而能够制约浮游植物的时空分布,是影响浮游植物生长的重要环境因子之一[28-30]。本次指沟卡尔藻赤潮暴发前、赤潮发生及维持阶段盐度在30.02~32.89之间,而2022年6月平潭海域的指沟卡尔藻赤潮暴发期间盐度在29.20~32.1之间[31-33],推测指沟卡尔藻的适宜盐度范围大致在29~33之间。虽然许多赤潮生物对于盐度没有严格的要求,但是指沟卡尔藻是一种不具壳片甲藻,其细胞壁只有平滑、有弹性的薄板[6,34],因此指沟卡尔藻细胞对于盐度变化极为敏感,剧烈的盐度变化可使其细胞渗透压改变,导致细胞破裂而死亡。因此,6月6日后,盐度的宽幅波动是指沟卡尔藻赤潮难以继续维持而迅速消亡的原因之一。
3.3 水文条件对赤潮的影响
莆田海域春、夏两季主要受浙闽沿岸流(低温、低盐)和台湾暖流(高温、高盐)的影响[35]。浙闽沿岸流3月至4月逐渐减弱,5月北撤至平潭岛附近海域,6月基本消失[36]。如表3所示,通过统计2017至2022年历年5月26日至6月6日期间南日岛海域(浮标数据)平均水温及盐度发现:相比前5年,2022年港南海域平均水温偏低1.58~4.23℃、平均盐度偏低1.44~3.74,坑口海域平均水温偏低0.95~3.98℃、平均盐度偏低1.87~3.59,均为近5年来最低(筶杯岛海域属于河口海域,因而不在此讨论范围内)。盐度32的等值线可以大体代表浙闽沿岸流的边界[35],直至6月6日港南海域盐度才升至32左右。由此推测,相较往年,2022年5月底至6月初莆田海域浙闽沿岸流偏强并占据主导地位。
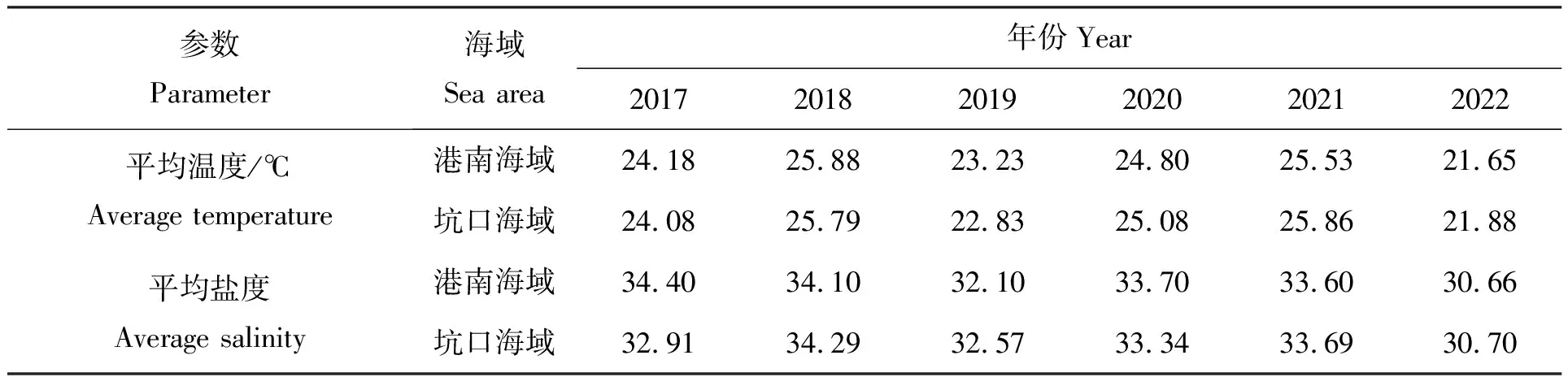
表3 南日岛海域2017—2022年5月26日至6月6日平均水温和盐度统计
较强的浙闽沿岸流带来的大范围低温海水对早期福建近岸甲藻赤潮优势种东海原甲藻的成长起到抑制作用[37],5月中旬南日岛海域东海原甲藻赤潮仅维持了5 d[38-39],这也间接为指沟卡尔藻赤潮暴发创造了条件。当浙闽沿岸流已经完全退出莆田海域时,其对东海原甲藻的抑制作用消失,这可能是6月9日坑口海域出现指沟卡尔藻赤潮向东海原甲藻赤潮演替的原因之一。
5月下旬至6月初,北向的台湾暖流的突然增强而引发的高温、高盐水体入侵,从而导致莆田海域浙闽沿岸流影响力减弱和近岸水体温度、盐度的迅速升高,冷暖水对峙的锋面向北移动。南日水道在台湾暖流和浙闽沿岸流两股水团交界处出现明显的温盐跃层[40]。在本次赤潮过程中,人工现场监测也发现不连续黑褐色、油膜状片状、有分层现象的水团。温盐跃层的出现使水体上下层之间交互不畅而导致分层,水体理化性质相对稳定,有利于指沟卡尔藻赤潮的形成和维持。
6月6日以后随着冷暖水对峙锋面的不断北移,叠加大风及大量降水,海域水体分层现象被打破,在浮标监测数据上则表现为水温、盐度等数值呈现宽幅波动。此后赤潮迅速消亡。因此,稳定水文条件是指沟卡尔藻赤潮得以发生和维持的重要条件。
3.4 营养盐对赤潮的影响
众多研究均表明富营养水质是赤潮暴发的重要条件。一般而言,近岸海域营养盐普遍充足,布设在港南海域和筶杯岛海域的浮标长期监测的数据也证明了这一点。硅藻和甲藻对温度和营养盐(氮、磷及其比值)变化的响应模式不同:硅藻偏好低温和高营养盐;而甲藻对温度和营养盐相对不敏感,但倾向于低磷和高氮磷比的环境。硅藻和甲藻的这些生态位特性差异决定了升温导致的低营养盐及富营养化引起的高N/P值都会促进甲藻的快速生长[41]。赤潮暴发前,港南海域的低磷、低营养盐环境有利于指沟卡尔藻在与其他硅藻竞争过程中抢占生态位。一般认为,浮游植物更偏好吸收氨氮,会优先利用氨氮,随后才是硝酸盐氮和亚硝酸盐氮[42]。这是由于氨氮属于还原态的氮,浮游植物细胞可直接利用其进行氨基酸等生命必需物质的合成,而硝酸盐氮和亚硝酸盐氮需要在细胞内被还原后才能被同化利用[42]。在深圳海域红色赤潮藻赤潮过程中,大鹏湾湾顶海域营养盐(尤其是氨氮)的突然升高被认为是导致赤潮的直接诱因[12]。本次指沟卡尔藻赤潮过程中也同样出现了营养盐特别是氨氮的突然升高现象。5月31日降水给港南海域带来大量的营养盐尤其是氨氮,一方面提供大量易被指沟卡尔藻吸收利用的氮源;另一方面,降水不仅提高了港南海域的营养盐含量,同时还提高了水体N/P值(如图5e所示)。这是因为富营养化引起的高N/P同样也促进了指沟卡尔藻的快速生长,加速了赤潮的暴发。由此推测,港南海域营养盐(尤其是氨氮)突然升高是本次指沟卡尔藻赤潮暴发的直接诱因。
3.5 赤潮扩散路径分析
结合现场人工采样及浮标数据分析,各海域受到赤潮影响从早到晚排序为港南海域最早、坑口海域和东岱海域次之、筶杯岛海域最晚。港南海域最早在5月29日下午pH值、溶解氧和叶绿素均同时出现阶段性峰值,叶绿素a含量更是高达17.64 μg/L;坑口海域在6月1日虽然已经检出高密度的指沟卡尔藻,但是整个海域的叶绿素a含量并不高,直至6月3日才出现高于10 μg/L的峰值;筶杯岛海域直至6月5日才检出指沟卡尔藻。由此推测,指沟卡尔藻赤潮最早应该在港南海域暴发。
兴化湾是受南日水道和兴化水道两股潮流作用,其中湾内南侧海域主要受南日水道潮流控制,湾内北侧和深槽海域主要受兴化水道潮流作用。涨潮时,南日水道潮流沿深槽南侧向湾顶运动[43]。兴化湾海域潮汐属于规则半日潮[43],5月30日至6月1日(农历五月初一至初三)是莆田海域的大潮期。大潮期间不仅带来了大量外海海水,同时也带来港南海域包含指沟卡尔藻细胞的水团。退潮时,坑口海域沿岸内湾性水域和石城渔港滞留了部分含有指沟卡尔藻细胞的水团。在表层风海流作用下,指沟卡尔藻在该海域沿岸大量聚集,并在适宜的条件下,其细胞群体暴发性增殖,而增殖后的种群又因弱风和相对缓慢的潮流无法扩散,最终导致在坑口海域和石城渔港内形成了赤潮。因此,潮流的聚集作用是坑口海域和石城渔港暴发赤潮的重要推手。
南日群岛内的主要潮流通道流向大致呈西北东南走向[44]。南日岛东部海域为敞开海域,与外海具有明显的交换作用。夏季南日岛外海存在一个低温高盐海水涌升中心[45]。据此推测,南日岛坑口海域指沟卡尔藻细胞群体随着潮流扩散至东岱海域,该海域低温和高盐的海水并非指沟卡尔藻最适的温盐条件,未能暴发性增殖,加之此处海水与外海交流通畅、潮流的聚集作用弱,因而未发生指沟卡尔藻赤潮。
随着南日岛海域与外海海水的交换,指沟卡尔藻细胞群体随着冷暖水对峙锋面的不断北移而扩散至更北部的海域。处于南日岛东北方向的平潭海域6月9日也报告发生了指沟卡尔藻赤潮,影响面积约8 km2,直至6月13日才消亡[31-33]。
综上所述,推测本次指沟卡尔藻赤潮扩散路径为:存在于港南海域的指沟卡尔藻“种子”细胞在适宜的条件下形成高密度的营养细胞群体,率先在港南海域形成赤潮,在潮水和冷暖水对峙锋面不断北移的作用下,扩散至石城渔港、坑口海域,继而进一步扩散至东岱海域、筶杯岛海域和平潭海域。
3.6 指沟卡尔藻赤潮的特点及毒性问题
本次赤潮过程海域水色正常,仅在部分时间部分海域(坑口海域)可见到异常情况,在赤潮发展及维持阶段,很难通过肉眼发现,也很难通过直接观察而估算赤潮面积,并且赤潮期间多次出现大雾和暴雨等不利情况,这给赤潮人工走航跟踪监测带来困难,反映出人工监测手段的不足之处,但浮标监测很好地弥补了这一缺点。
在1997年日本下关渔港指沟卡尔藻赤潮中,指沟卡尔藻密度最高在3.00×106~5.00×106cell/L之间,大部分海域超过5.00×105cell/L,造成了鱼类大量死亡,而104cell/L海域则未出现鱼类死亡[5-6]。在1998年我国香港海域“红潮”过程中,当监测点海域指沟卡尔藻最高密度为2.1×105cell/L时,该海域附近的渔场就已经受到影响[46]。2019年5月福建平潭发生大规模指沟卡尔藻赤潮,5月23至25日该海域指沟卡尔藻最高密度在3.60×105~9.80×105cell/L之间,造成了鱼类大量死亡[47-49]。本次指沟卡尔藻赤潮现场监测发现,当海域中指沟卡尔藻密度达到104cell/L时,其就能造成网箱养殖的小规格大黄鱼(0.1 kg以下)大量死亡,105cell/L密度就可以造成大黄鱼(含成鱼)、鲈鱼、红鱼、真鲷、黑鲷等网箱养殖的鱼类大量死亡。显然,按照赤潮监测技术规程(HY/T 069—2005)[20]中相关标准(指沟卡尔藻赤潮基准浓度定为1.0×106cell/L),此时已经非常不利于赤潮的防灾减灾了。指沟卡尔藻赤潮如何引起鱼类的死亡,以及其是否产生毒素及毒素的理化特性等问题还需进一步研究。
致谢:莆田市水产科学研究所王春忠研究员、莆田市海洋与渔业环境监测站张丽高级工程师及全体同仁在样品采集、数据整理上给予诸多支持,在此一并致谢。
猜你喜欢
矿山安全信息(2020年8期)2020-12-20
矿山安全信息(2020年39期)2020-12-20
矿山安全信息(2020年29期)2020-01-05
矿山安全信息(2020年44期)2020-01-04
广东海洋大学学报(2015年3期)2015-12-22
应用海洋学学报(2015年3期)2015-11-22
西安建筑科技大学学报(自然科学版)(2014年2期)2014-11-12
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
棋艺(2001年9期)2001-07-17
棋艺(2001年11期)2001-05-21
