
蓝莓不同砧穗组合嫁接亲和性研究
2022-10-28 08:09:54顾莉莉沈朱俐康海婷李永强徐丽珊郭卫东
浙江师范大学学报(自然科学版) 2022年4期
顾莉莉, 沈朱俐, 宗 宇,2, 康海婷,李永强,2, 徐丽珊,2, 郭卫东,2
(1.浙江师范大学 化学与生命科学学院,浙江 金华 321004;2.浙江省特色经济植物生物技术重点实验室,浙江 金华 321004)
嫁接是果树生产中被广泛使用的技术.通过嫁接可以保持果树品种的优良性状,快速进行品种的更新换代.嫁接在园艺作物种质资源保存、抗逆栽培、树体大小和花期调控、砧穗互作等方面有较多应用[1-5],也是研究植物体内物质运输[6]、成花机制[4-5]、信号转导及细胞识别机制[7-8]等基础理论的重要方法和手段.果树不同种或品种经过嫁接形成嫁接体需要经历复杂的生理生化过程,嫁接成功的基础之一是砧木和接穗完全愈合,不同物种的愈合虽略有差异,但几乎都需要经历4个阶段:从嫁接面产生愈伤组织;砧穗相互接触的细胞形成胞间连丝;砧穗之间分化出形成层并相互连接;砧穗的木质部及维管束连接等[9-10].其中,任何一个环节受到阻碍都会影响到砧穗之间嫁接的亲和性和成活率[7,11-13].
高丛蓝莓(Vacciniumcorymbosum)是我国蓝莓主要的栽培类型,因其具有无根毛、根系浅等特点,从而导致树体的生长容易受到水分和高温胁迫,对栽培的土壤环境要求非常严格.蓝莓是典型的嗜酸性果树,树体在土壤pH为4.5~5.5时生长旺盛,当土壤pH高于6时,因其根际土壤酸化能力较弱,导致高丛蓝莓生长受阻,主要表现为叶片失绿黄化,根系颜色变深,最终导致植株死亡[14-15].高丛蓝莓苗通过组织培养获得的自根无性系进行繁育,生成的不定根抗性差,组培苗细弱,生根和炼苗过程繁琐.除此之外,高丛蓝莓成熟果实的采收几乎完全依靠人工完成,耗时耗力且成本高,采收不及时会造成果实的大量浪费.嫁接有助于改良蓝莓树体结构,减少灌丛枝的发枝量,可以显著提高蓝莓树体的通透性,有利于成熟果的机械化采收.虽然已有利用臼莓和乌饭树[16]做砧木进行蓝莓嫁接栽培的研究,但本实验室前期研究发现,一些越橘属野生种与高丛蓝莓亲缘关系较远[17-18],嫁接后可能存在亲和性差、嫁接成活率低、接穗生长不良等情况.兔眼蓝莓(V.ashei)是高丛蓝莓的近缘种,相比之下其根系更发达,土壤环境适应能力更强,能够在pH6.5的低有机质土壤中旺盛生长,是通过嫁接方式改良高丛蓝莓植株生长势,增强高丛蓝莓根系适应性的良好备选砧木.
我国蓝莓栽培起步晚,发展迅速,已经成为世界上蓝莓产量最大的国家之一[19].上述多个因素限制了蓝莓栽培范围的扩大,严重影响了蓝莓产业的健康可持续发展.选择根系发达、适应广域土壤pH范围的砧木进行嫁接栽培可以使蓝莓植株获得更好的抗性,进一步拓宽了蓝莓栽培的地域范围.本研究以9种兔眼蓝莓为砧木,2种高丛蓝莓为接穗进行嫁接试验,统计了嫁接后不同时期各种砧穗组合的成活率,测定了嫁接口直径、接穗叶片中的叶绿素相对含量和嫁接成活相关酶活力,并进行了相关性分析,以期筛选出亲和性高的蓝莓砧穗组合,为蓝莓嫁接栽培和嫁接相关基础研究提供参考.
1 材料和方法
1.1 植物材料
试验在浙江师范大学蓝莓种质资源圃中进行.选取3年生盆栽的‘红宝石’‘乡铃’‘顶峰’‘波尼塔’‘蓝沙’‘乌达德’‘贝姬蓝’‘园蓝’和‘杰兔’等9个兔眼蓝莓品种为砧木,对砧木品种进行短截处理,保留距离基部10 cm的单个枝条进行嫁接.从6年生的‘奥尼尔’和‘实大’2个高丛蓝莓品种植株上剪取1年生枝,将其剪成5 cm的带芽茎段做接穗,于2018年2月使用劈接法进行嫁接.不同砧穗组合的蓝莓经嫁接处理后采用避雨栽培,常规水肥管理,嫁接后第40 d剪除嫁接口处的绑缚膜.
1.2 方 法
1.2.1 嫁接成活率统计
在嫁接后第180,360和540 d对不同砧穗组合的成活率进行3次统计,将嫁接植株成活的标准设定为:接穗上至少有1个萌发出≥5 cm的新梢,成活株数所占最初嫁接总株数的百分比为嫁接成活率.
1.2.2 接穗叶片中叶绿素相对含量的测定
分别于嫁接后的第180,360和540 d用手持式SPAD-502 Plus叶绿素仪(Konica Minolta Investment Ltd.,日本)测量接穗叶片中的相对叶绿素含量.选取生长于接穗中部的健康成熟叶片,用湿纸巾擦除叶片表面灰尘.每个砧穗组合随机选择3棵嫁接成活蓝莓,每棵测定5个叶片,每个叶片固定选取中部较宽部位测定3次,平均值作为该嫁接组合叶片的SPAD值,即叶绿素相对含量.
1.2.3 嫁接口直径测量
用游标卡尺对嫁接第180,360和540 d后的嫁接口直径进行测量.为防止嫁接口因膨大或残损影响测量的准确度,以嫁接口上方1 cm处的枝条直径作为嫁接口直径.每个嫁接口测量3次,每个嫁接组合随机选择3株进行测量,平均值作为该嫁接组合的嫁接口直径.
1.2.4 接穗叶片中酶活力的测定
分别于嫁接后第180,360和540 d,采集不同嫁接组合的接穗中部成熟叶片,测定PPO,POD,PAL和SOD酶活力.每个嫁接植株采集3个叶片,每个组合采集10个单株进行酶活力测定.PPO的提取与活力测定参考张银杰等[20]的方法.PAL和POD的提取与活力测定分别参考Rohde等[21]和Chen等[22]的方法.SOD的提取与活力测定采用氮蓝四唑光还原法[23].
1.3 数据分析
采用Microsoft Excel 2013进行实验数据处理并绘图,相关性分析圆圈图使用R语言包CORRPLOT[24]绘制,使用SPSS 23.0软件进行单因素方差分析(Duncan法,P<0.05),
2 结果与分析
2.1 蓝莓不同嫁接组合的成活率
研究分别嫁接了563个‘实大’接穗和476个‘奥尼尔’接穗.随着嫁接后天数的延长,嫁接在兔眼蓝莓砧木上的‘实大’和‘奥尼尔’成活率均表现出下降趋势.‘实大’在嫁接后第180 d的平均成活量为218株,平均成活率为34.40%,嫁接后第360 d时的平均成活率为29.29%,在第540 d时的成活率为23.29%.其中,‘实大-顶峰’在嫁接后第540 d的成活率最高,为37.21%;‘实大-蓝沙’次之,为30.51%;‘实大-波尼塔’成活率最低,仅为19.05%(见表1).

表1 不同砧木上的‘实大’和‘奥尼尔’蓝莓的嫁接成活率 单位:%
随着嫁接时间延长,‘奥尼尔’的嫁接成活率逐渐降低,这种趋势与‘实大’相似,但‘奥尼尔’成活率阶段性的变化大于‘实大’.‘奥尼尔’在嫁接后第180 d,成活244株,平均成活率为42.39%;在嫁接后第360 d,嫁接成活率降至36.23%,存活嫁接组合206株;嫁接后第540 d时,‘奥尼尔’成活率快速降至20.78%,仅存活111株.‘奥尼尔-杰兔’砧穗组合平均嫁接成活率最高,在嫁接后第540 d,成活率为32.76%;‘奥尼尔-红宝石’次之,为30.00%;‘奥尼尔-波尼塔’最低,仅为14.29%.嫁接在‘顶峰’和‘杰兔’上的‘实大’和‘奥尼尔’均有较高的成活率,而嫁接在‘波尼塔’‘贝姬蓝’和‘园蓝’的‘实大’和‘奥尼尔’成活率显著小于其他6种砧木类型.
2.2 蓝莓不同嫁接组合叶片中叶绿素的相对含量
对嫁接后不同时期的‘实大’和‘奥尼尔’叶片中叶绿素的相对含量进行了测定,结果表明,随着嫁接后时间的增加,‘实大’和‘奥尼尔’叶片中的叶绿素平均相对含量呈现下降趋势,‘实大’的降幅大于‘奥尼尔’.‘实大-顶峰’和‘实大-杰兔’嫁接组合中叶片叶绿素的相对含量与其他组合不同,表现为先下降后升高的趋势(见表2).
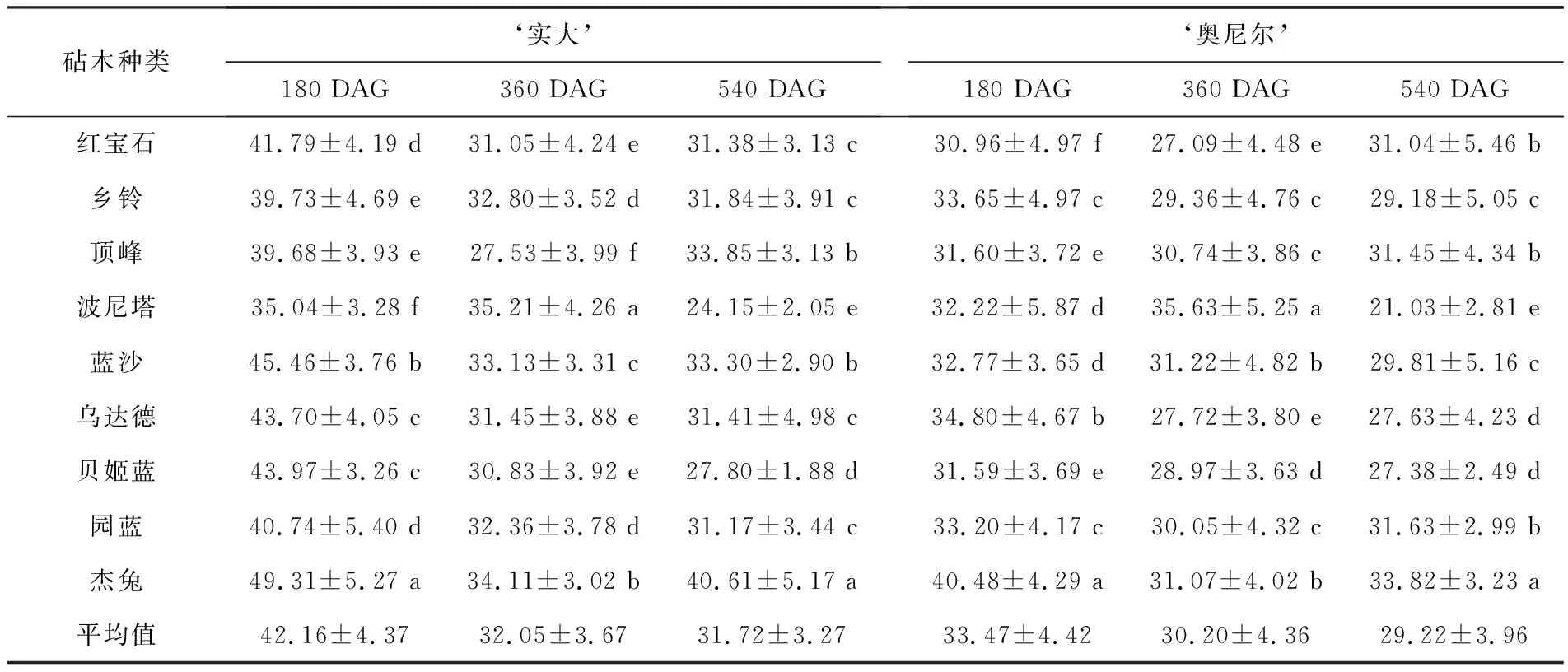
表2 不同砧木上的‘实大’和‘奥尼尔’蓝莓叶片的叶绿素相对含量
嫁接后第540 d,‘实大-杰兔’嫁接组合叶片中叶绿素相对含量显著高于其他嫁接组合,SPAD值为40.61±5.17,‘实大-顶峰’次之,SPAD为33.85±3.13.嫁接在‘波尼塔’和‘贝姬蓝’蓝莓上的‘实大’叶片SPAD显著小于其他组合,其SPAD值分别为24.15±2.05和27.80±1.88,‘实大-园蓝’的SPAD值为31.17±3.44,虽然显著高于‘实大-波尼塔’和‘实大-贝姬蓝’,但与嫁接在‘红宝石’和‘乡铃’上的‘实大’叶片叶绿素相对含量没有显著差异.
嫁接在不同砧木上的‘奥尼尔’蓝莓叶片中叶绿素相对含量变化趋势不同于‘实大’.在180 DAG时叶片SPAD的平均值为33.47±4.42,嫁接后第360和540 d的叶片SPAD分别下降为30.20±4.36和29.22±3.96(见表2).嫁接在不同砧木上的‘奥尼尔’叶片SPAD值存在较大差异.‘奥尼尔-红宝石’和‘奥尼尔-杰兔’呈现先下降后上升的趋势;‘奥尼尔-乡铃’‘奥尼尔-波尼塔’和‘奥尼尔-贝姬蓝’呈现出持续下降的趋势;其他组合中叶片SPAD值变化较小.在540 DAG时‘奥尼尔’不同砧穗组合中,‘奥尼尔-杰兔’叶片SPAD值最大,为33.82±3.23,‘奥尼尔-园蓝’次之,为31.63±2.99,‘奥尼尔-波尼塔’最小,为21.03±2.81.
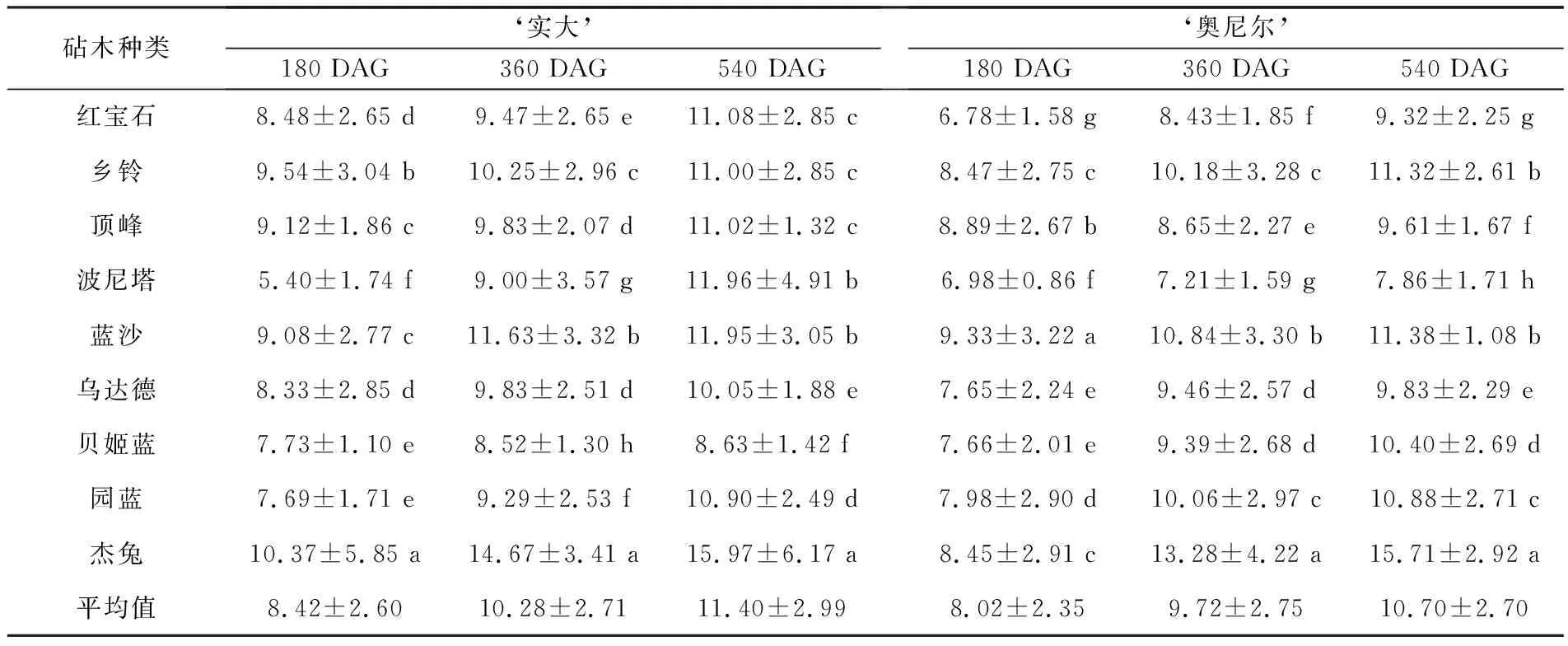
表3 不同砧木上的‘实大’和‘奥尼尔’蓝莓的嫁接口直径 单位:mm
2.3 蓝莓不同嫁接组合的生长量
随着嫁接后切口的愈合和接穗的生长,不同嫁接组合的嫁接口直径逐渐增大,而且后期增速加快.如图3所示,嫁接在不同砧木上的‘实大’嫁接口直径平均值为(11.40±2.99) mm,‘奥尼尔’嫁接口直径平均值为(10.70±2.70) mm.嫁接后第180 d,‘实大’和‘奥尼尔’嫁接组合之间生长量差异显著.‘实大-杰兔’嫁接口的直径显著大于以‘实大’为接穗的其他砧穗组合,其嫁接口直径为(10.37±5.85) mm;‘实大-乡铃’次之,为(9.54±3.04) mm;最小嫁接口直径(5.40±1.74) mm出现在‘实大-波尼塔’组合.从360 DAG开始,各砧穗组合之间的生长量差异逐渐减小,在540 DAG时,‘实大-杰兔’仍是嫁接口直径最大的组合,砧木‘红宝石’‘乡铃’和‘顶峰’之间没有显著性差异.
在180 DAG时,‘奥尼尔’的不同砧穗组合中‘奥尼尔-蓝沙’嫁接口最大直径为(9.33±3.22) mm.从360 DAG开始,‘奥尼尔-杰兔’的生长量快速增加,360 DAG时嫁接口直径为(13.28±4.22) mm,540 DAG时为(15.71±2.92) mm,显著大于嫁接在其他砧木上的‘奥尼尔’蓝莓.不同砧穗组合在540 DAG时仍表现出显著性差异,嫁接口直径最大为‘奥尼尔-杰兔’组合,其次是‘奥尼尔-蓝沙’和‘奥尼尔-乡铃’,其嫁接口直径分别为(11.38±1.08)和(11.32±2.61) mm.‘奥尼尔-波尼塔’的嫁接口直径最小,仅为(7.86±1.71) mm,显著小于其他以‘奥尼尔’为接穗的砧木组合.
2.4 蓝莓不同嫁接组合叶片中的酶活力
测定了不同的砧穗组合嫁接后第180,360和540 d的叶片PPO,POD,PAL和SOD酶活力,4种酶的活力变化在不同砧穗组合中呈现较大差别(见图1和图2).‘实大’PPO酶活力的变化趋势多为先升高后下降,最高酶活力出现在‘实大-红宝石’,为17.90 U·mg-1FW(见图1).嫁接在‘顶峰’‘蓝沙’和‘杰兔’上的‘实大’PPO酶活力表现为持续升高的趋势,3种砧穗组合的最大值均出现在嫁接后第540 d,其酶活力分别为15.10,16.99和13.60 U·mg-1FW.‘实大-贝姬蓝’组合中PPO酶活力虽然也呈现先升高后降低的趋势,但其酶活力显著小于其他砧穗组合,360 DAG时PPO出现最大值,仅为0.43 U·mg-1FW.
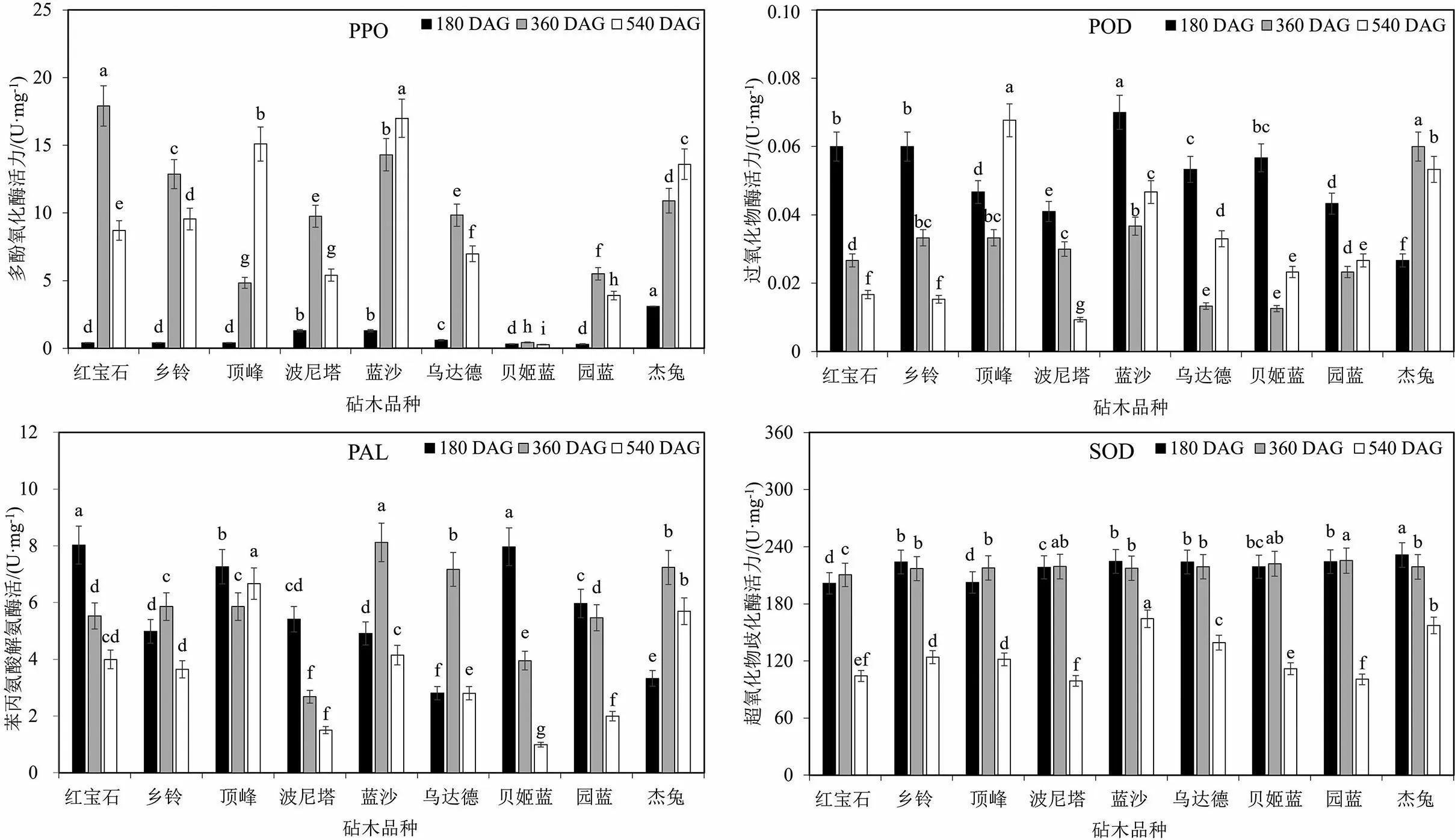
差异显著性分析采用邓肯新复极差法,处理之间不同小写字母表示存在显著性差异(P<0.05)
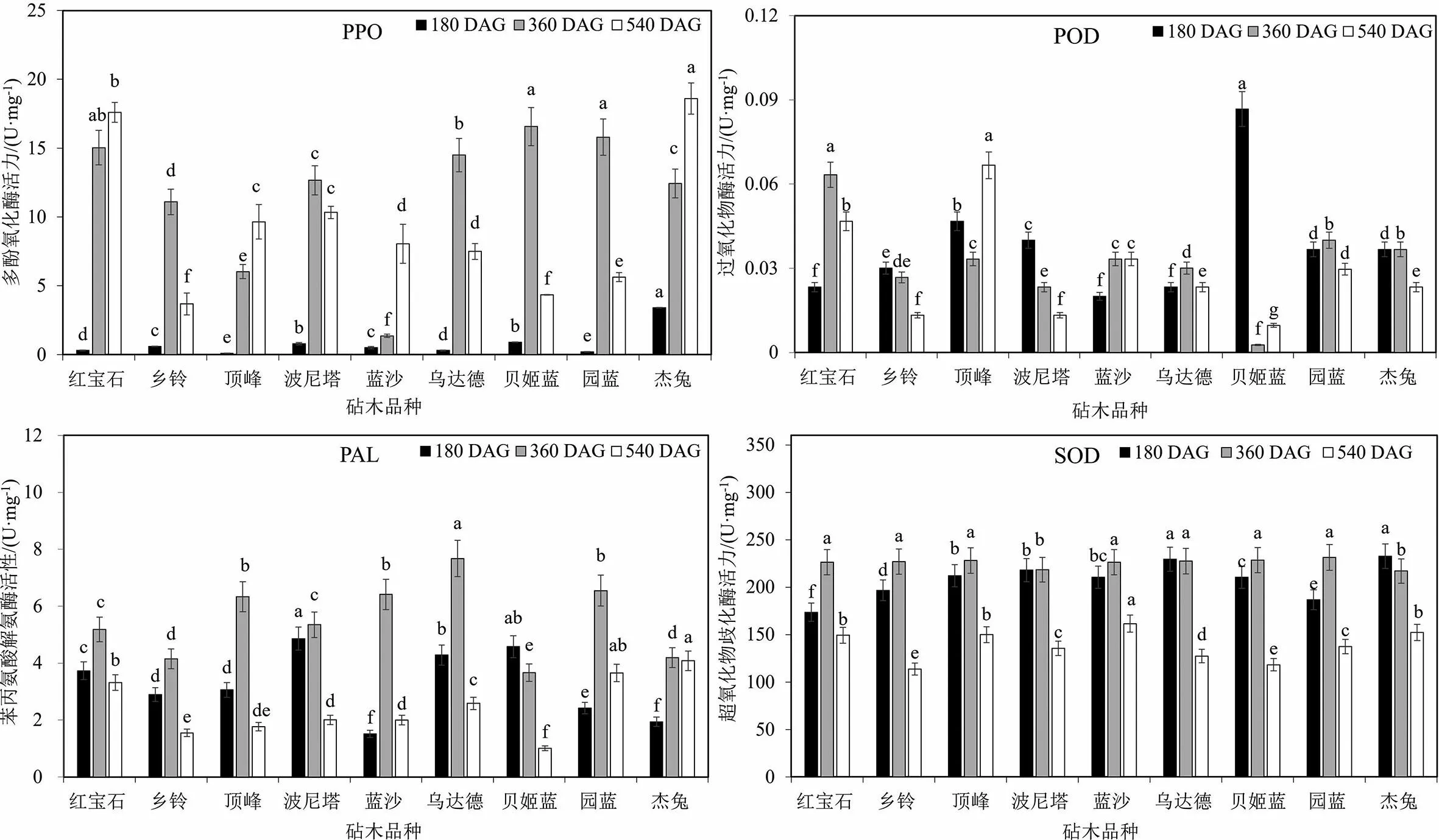
图2 嫁接在不同砧木上的‘奥尼尔’蓝莓叶片中PPO,POD,PAL和SOD酶活力
PAL酶活力的变化在以‘实大’为接穗的不同砧穗组合中出现多样化趋势.嫁接在‘红宝石’‘波尼塔’‘贝姬蓝’和‘园蓝’上的‘实大’叶片PAL酶活力随着嫁接后时间的延长出现降低的趋势,而以‘乡铃’‘蓝沙’‘乌达德’和‘杰兔’为砧木则表现为先升高后降低的趋势.以‘红宝石’‘顶峰’‘贝姬蓝’和‘园蓝’为砧木时,180 DAG的‘实大’蓝莓PAL酶活力均显著高于同时期的其他砧穗组合.以‘蓝沙’‘乌达德’和‘杰兔’为砧木时,360 DAG的‘实大’PAL酶活力显著高于同时期的其他砧穗组合,其酶活力分别为8.12,7.16和7.24 U·mg-1FW.
不同砧穗组合中POD酶活力的变化大部分为先降低再升高,而且POD酶活力在4种酶中最低.除‘顶峰’和‘杰兔’之外,嫁接在其他砧木上的‘实大’180 DAG均出现了最大值.‘实大-顶峰’和‘实大-杰兔’组合中酶活力最大值分别出现在540 DAG和360 DAG,它们分别为0.068和0.060 U·mg-1FW.4种酶中活力最高的是SOD酶,不同砧穗组合的SOD酶活力在180 DAG和360 DAG时期虽然出现了显著性差异,但同一砧木的2个时期间酶活力的差值较小,只有‘实大-顶峰’和‘实大-杰兔’2个组合较大,其差值分别为15.16和12.44 U·mg-1FW,‘实大-顶峰’SOD酶活力最大值出现在360 DAG,后者最大值出现在180 DAG.‘实大-杰兔’180 DAG的SOD酶活力显著高于同时期的其他组合,为231.33 U·mg-1FW.所有组合的SOD酶活力在540 DAG时表现为显著下降.
以‘奥尼尔’为接穗的嫁接组合中,PPO酶活力的变化趋势与嫁接在不同砧木上的‘实大’相似(见图1和图2).PPO酶活力大部分表现为先升高后降低的变化趋势,嫁接在‘红宝石’‘顶峰’‘蓝沙’和‘杰兔’上的‘奥尼尔’叶片PPO酶活力在3个时期均为上升趋势.其中‘奥尼尔-杰兔’组合在180 DAG和540 DAG时期的PPO酶活力显著高于其他砧穗组合,分别为3.40和18.60 U·mg-1FW.‘奥尼尔’叶片中PAL酶活力在不同砧木上的变化趋势较为一致,除‘奥尼尔-贝姬蓝’以外,嫁接后3个时期酶活力的变化均为先升高后降低.‘奥尼尔-贝姬蓝’组合中叶片PAL酶活力从180 DAG到540 DAG呈现连续下降的趋势,而且其360 DAG和540 DAG时期的PAL酶活力显著低于其他组合,分别为3.67和1.01 U·mg-1FW.
以‘奥尼尔’为接穗的砧穗组合中叶片POD酶活力呈现多种变化趋势,其中‘奥尼尔-红宝石’‘奥尼尔-乌达德’‘奥尼尔-园蓝’和‘奥尼尔-杰兔’4个组合先升高再降低,‘奥尼尔-乡铃’和‘奥尼尔-波尼塔’组合中POD酶活力呈现连续下降的趋势.POD酶活力的最大值出现在180 DAG的‘奥尼尔-贝姬蓝’,为0.086 U·mg-1FW,显著高于其他组合的酶活力,360 DAG和540 DAG时,‘奥尼尔-贝姬蓝’中的酶活力快速下降,分别为0.003和0.010 U·mg-1FW,显著小于其他组合中的酶活力.
嫁接在不同砧木上的‘奥尼尔’叶片SOD酶活力的变化趋势与‘实大’不同砧穗组合相似,大部分酶活力呈现先上升后下降的趋势,只有‘奥尼尔-乌达德’和‘奥尼尔-杰兔’呈现连续下降的趋势,且以‘乌达德’和‘杰兔’为砧木的‘奥尼尔’叶片中SOD酶活力显著高于其他组合,分别为229.44和232.58 U·mg-1FW.
2.5 嫁接体发育过程中生理生化指标的相关性
对不同砧穗组合嫁接体发育过程中的生理生化指标进行了相关性分析,图3表明,接穗‘实大’的不同组合在180 DAG时PPO酶活力与PAL酶活力、POD酶活力均呈现显著负相关,相关系数分别为-0.57和-0.59,PPO酶活力与SOD酶活力之间为显著正相关,相关系数为0.52,而PAL酶活力与SOD酶活力是显著负相关的关系,相关系数为-0.75.PPO酶活力与叶绿素相对含量存在相关性,相关系数为0.54.生理指标与酶活力在360 DAG时存在多个相关性.嫁接成活率与PAL酶活力显著正相关,相关系数为0.81,嫁接口直径与PAL酶活力、POD酶活力存在显著正相关关系,相关系数分别为0.66和0.88.PPO酶活力与SOD酶活力在该时期呈现显著负相关关系,相关系数为-0.77.在540 DAG时,嫁接成活率、叶绿素相对含量与PPO酶活力、PAL酶活力均呈现显著正相关关系,嫁接口直径与酶活力在此时期有相关性,但相关系数小于同时期其他指标间的相关系数.

A:PPO酶活力;B:PAL酶活力;C:POD酶活力;D:SOD酶活力;E:嫁接成活率;F:叶绿素相对含量;G:嫁接口直径
以‘奥尼尔’为接穗的嫁接体发育过程中生理生化指标的相关性与‘实大’有较大差别(见图4).在180 DAG时,PPO酶活力与接穗叶片中叶绿素相对含量呈显著正相关关系,相关系数为0.87.PAL与嫁接口直径之间为显著负相关,相关系数为-0.78.在360 DAG时,叶绿素相对含量与嫁接成活率之间为显著正相关,相关系数为0.72;与SOD酶活力之间为显著负相关,相关性系数为-0.60.在180 DAG和360 DAG 2个时期其他生理生化指标间不存在显著相关性.在540 DAG,嫁接成活率与PAL酶活力、叶绿素相对含量和嫁接口直径呈显著正相关,叶绿素相对含量与嫁接口直径之间也呈现显著正相关,相关性系数为0.69.
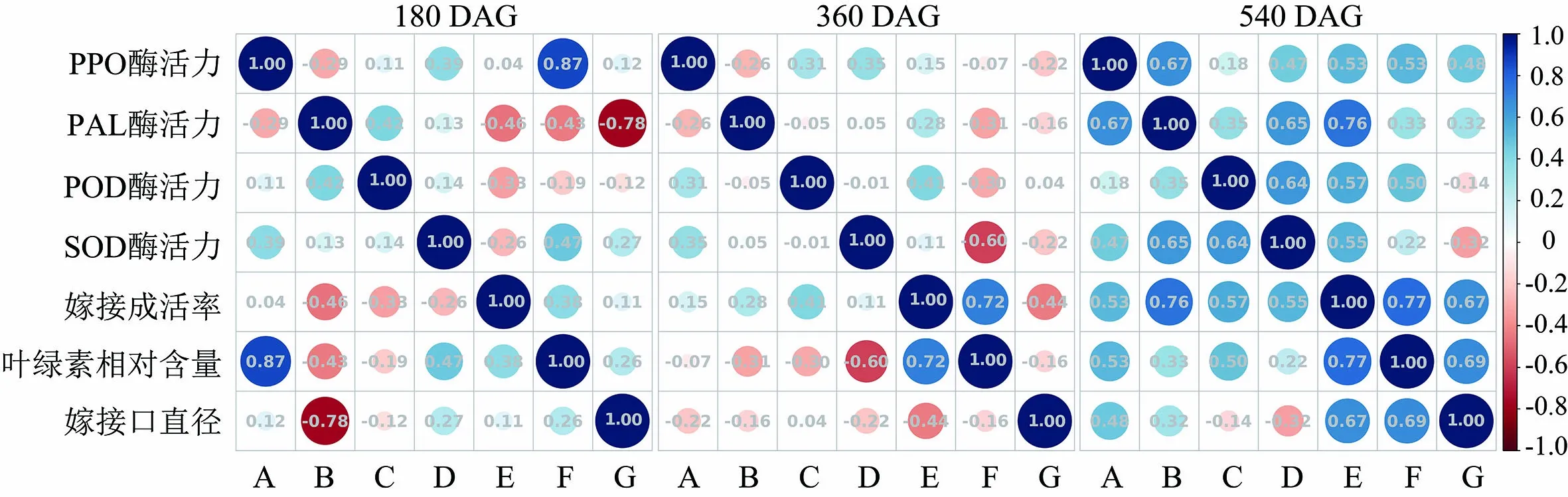
A:PPO酶活力;B:PAL酶活力;C:POD酶活力;D:SOD酶活力;E:嫁接成活率;F:叶绿素相对含量;G:嫁接口直径
3 讨 论
砧穗嫁接体能否愈合并成活是由多因素共同决定的,亲和力高是嫁接成功的前提,虽然通过嫁接后不同时间点的成活率可以判断亲和力的强弱,但在果树等木本植物中存在后期不亲和现象[10,22,25-26].本研究中,‘实大’和‘奥尼尔’为接穗的蓝莓不同砧穗组合从180 DAG到540 DAG大部分表现出成活率下降,这说明蓝莓嫁接过程中很可能存在后期不亲和现象;虽然后期不亲和情况随着嫁接时间的延长趋于稳定,但嫁接在兔眼蓝莓上的‘奥尼尔’后期不亲和情况比‘实大’更严重.‘奥尼尔’是‘实大’的父本,二者遗传背景相似[17-18],嫁接后‘奥尼尔’接穗叶片中叶绿素相对含量和嫁接口直径的平均值小于‘实大’,暗示‘实大’与兔眼蓝莓的亲和性更高.
嫁接过程中植物遭受切割伤会释放多酚类物质,酚类物质可以促进愈伤细胞分化[3,7,10-11,27],影响生长素的合成及运输,从而在嫁接体发育过程中通过促进形成层的发育,影响新木质部和韧皮部的分化过程及木质化过程[7,8,10-11].PPO基因受机械损伤的诱导,PPO在生物体酶促褐变、体内色素合成的过程中起关键性作用[28-29],PAL是催化苯丙烷代谢途径的第1个关键酶[30-31].苯丙烷代谢途径可调控下游木质素的合成,当植物遭受机械损伤时,这些物质可参与伤口愈合,在昆虫和病原菌入侵时也可以起到免疫作用[21,31-32].POD催化过氧化氢参与的各种氧化反应,它可以通过氧化嫁接面的吲哚乙酸调节生长素的含量,PAL和POD都可促进木质素合成和管状分子的形成[32].SOD能够清除生物氧化过程中产生的超氧自由基[22],荔枝中的研究表明,嫁接后9个月的穗叶片中PPO,POD和SOD酶活力越高,其组合的嫁接亲和性越高,对应的嫁接成活率也越高[22],但这种相关性受到植物砧穗愈合时间的影响.本研究在蓝莓不同砧穗组合中发现180 DAG时,PPO,PAL,POD和SOD酶活力与嫁接成活率没有显著相关性,在360 DAG时,接穗叶片中PPO和POD酶活力与嫁接成活率存在一定程度的相关性,但未达到显著水平;‘实大’叶片中PAL酶活力与嫁接成活率存在显著正相关关系,在540 DAG时,4种酶的活力与嫁接成活率才表现为显著正相关.本实验室在田间对嫁接后1年的高丛蓝莓和兔眼蓝莓进行应力测试时,发现折断的砧穗愈合处仍保留着完整的嫁接切口,且仅在切口周围由少量韧皮部和木质部连接砧木和接穗(图片未呈现),暗示高丛蓝莓和兔眼蓝莓嫁接后维管组织完全重连可能需要更多时间.
砧木对嫁接植株的高度和冠幅等生长势有影响,嫁接亲和性高的植株生长势更强[4,33-38],这与本研究结果一致.蓝莓砧穗组合嫁接成活率越高,其嫁接口直径越大,嫁接在‘杰兔’‘蓝沙’砧木上的‘实大’和‘奥尼尔’的嫁接口直径都显著大于其他组合.嫁接后的蓝莓树体比组培繁育的自根苗生长更加旺盛,对土壤环境有更好的适应能力,可以使得接穗蓝莓提早开花和结果[16].砧木与接穗之间的亲和力强弱还会造成接穗叶片中的叶绿素含量存在显著差异,进而对接穗叶片的光合能力产生影响[38-39].嫁接在‘杰兔’‘蓝沙’上的‘实大’和‘奥尼尔’在嫁接后不同时期叶片中叶绿素相对含量变化量较小,且平均含量显著高于其他砧穗组合,这进一步说明‘杰兔’‘蓝沙’与‘实大’‘奥尼尔’亲和性高,可以用作高丛蓝莓的栽培砧木.
4 结 论
兔眼蓝莓‘杰兔’和‘蓝沙’均可作‘实大’和‘奥尼尔’的嫁接砧木.‘实大-杰兔’和‘奥尼尔-杰兔’组合嫁接成活率、嫁接后接穗中叶绿素相对含量和嫁接口直径的生长量显著高于其他组合.蓝莓砧穗组合的完全愈合需要较长的周期,且存在后期不亲和现象.在嫁接第540 d之后,接穗叶片中PAL,PPO和POD酶活力与最终的嫁接成活率存在正相关关系,PAL酶活力可以用作蓝莓砧穗嫁接亲和力强弱的检测指标.
猜你喜欢
现代园艺(2022年11期)2022-06-09 05:06:28
河北果树(2021年4期)2021-12-02 01:15:08
河北果树(2021年4期)2021-12-02 01:14:40
中国生态农业学报(中英文)(2021年8期)2021-07-28 10:32:52
中国果业信息(2020年12期)2020-12-15 10:03:38
天津外国语大学学报(2020年4期)2020-08-24 06:59:18
英语文摘(2020年1期)2020-08-13 07:25:46
音乐教育与创作(2020年2期)2020-06-19 06:45:46
商周刊(2017年5期)2017-08-22 03:35:22
作文周刊·小学一年级版(2017年26期)2017-08-10 20:31:15
