
不同恢复年限红锥人工林群落结构特征及经营策略
2022-10-27 08:21陈富强黄明智叶秋慧刘菊秀
安徽农业大学学报 2022年1期
陈富强,郭 勇,黄明智,冯 蔚,叶秋慧,刘菊秀,李 旭,3*
(1. 广东省龙眼洞林场,广州 510520;2. 中国科学院华南植物园退化生态系统植被恢复与管理重点实验室,广州 510650;3. 中国科学院大学,北京 100049)
全球森林正进行大规模的恢复计划和工程,如封山育林和集约化人工造林[1]。正确理解自然或人为干扰发生后森林的恢复过程是生态学领域和全球森林可持续管理的核心内容,尤其是在持续退化的热带和亚热带森林[2-3]。有研究表明,无论有无人为协助森林恢复,其物种的多度、多样性和生态系统功能等在停止干扰恢复30 多年后仍低于各自的参考水平[4-5]。由于物种多样性和生态系统功能在扰动后的恢复过程中可能呈现非线性关系[6],因此,及时把握森林生态系统恢复的现状和程度,正确评估退化生态系统恢复过程中生态系统结构和功能的动态变化显得愈加重要。生物多样性的分布格局和维持机制一直是群落生态学研究的热点与核心问题。物种多样性是反映该群落内物种组成状况的重要指标,即物种丰富度和均匀度[7-8]。研究植物群落物种多样性特征对揭示群落、生态系统结构和功能演变具有极为重要的意义[9-12]。
森林作为陆地生态系统的主体碳库,在应对全球气候变化具有独特的功能,在全球碳平衡中具有重要的作用[13]。人工林因成林迅速,已成为恢复森林增长、抵减森林消耗的一个关键措施。目前我国人工林呈现持续增长的态势,人工林面积继续保持世界首位[14],为全球绿化做出了巨大贡献[15]。然而,由于我国人工林普遍树种单一、结构简单、生态功能脆弱等缺陷,导致随着林龄的增加,会出现植物生物多样性下降、养分限制加剧、土壤微生物活性降低等生态系统功能失调方面的问题[16],可能会严重影响其固碳潜力。因此,了解处于不同林龄阶段植物群落结构和发育进程显得尤为重要。
红锥(Castanopsis hystrix)是壳斗科锥属常绿阔叶高大乔木,也是热带亚热带地区生长较快的珍贵用材树种[17],红锥人工林是我国南亚热带种植较为广泛的人工林。已有学者对红锥人工林的生长规律[18]、生理生态[17]、土壤理化性质及生态化学计量比[19]、微生物生物量、植物、土壤和微生物因子等[20]以及红锥人工林生物量和碳储量进行了评估[21-22],但很少关注不同林龄阶段的人工林结构特征与森林经营的关系,以及如何基于不同林龄阶段来开展人工林经营模式和后续经营作业措施[23-25]。因此,有必要对林分进行科学的森林经营管理[26]。本研究中,作者以广东省龙眼洞林场的不同林龄红锥人工林为研究对象,采用空间代替时间的方法,建立了不同林龄阶段样地并调查数据,尝试从经营的方向和群落动态来阐述不同林龄阶段的经营目标和预期状态,以期为红锥人工林经营作业法设计提供目标和过程的参考性对象依据。
1 材料与方法
1.1 研究地概况
研究地位于广东省广州市东北郊广东省龙眼洞林场,地理坐标为 113°21'~113°27' E,23°10'~23°18' N,属亚热带季风气候,年平均温度21.7 ℃,年平均降水量1 694 mm。现有林地面积1 622 hm2,生态公益林面积约占932 hm2为63%,海拔在100~538 m 之间,土壤类型为赤红壤,主要由花岗岩和砂页岩发育而成。该区人工林林分结构简单,乔木以红锥为主。
历史上该区域的南亚热带常绿阔叶林遭受破坏、形成裸地后,通过人工种植速生树种马占相思(Acacia mangium)进行初步的人工造林。后期经过皆伐后的林分改造,形成了处于不同林龄阶段的次生红锥群落。由于种植5 年后统一停止人为干扰(在种植第3年和第5 年进行抚育),逐渐出现林下植被,在一些林龄较高的红锥人工林中,三桠苦(Melicope pteleifolia)和鼠刺(Itea chinensis)因适应性强等优势而成为最先侵入的优势木本植物,其他乔木主要包括银柴(Aporosa dioica)、山油柑(Acronychia pedunculata)、罗伞树(Ardisia quinquegona)、水锦树(Wendlandia uvariifolia)、红花荷(Rhodoleia championii)、锥(Castanopsis chinensis)、木荷(Schima superba)、毛冬青(Ilex pubescens)、秤星树(Ilex asprella)和黄桐(Endospermum chinense)等。林下主要的灌木物种有:九节(Psychotria asiatica)和假鹰爪(Desmos chinensis)等;主要的草本物种有:乌毛蕨(Blechnum orientale)、扇叶铁线蕨(Adiantum flabellulatum)、团叶陵齿蕨(Lindsaea orbiculata)、芒萁(Dicranopteris dichotoma)、淡竹叶(Lophatherum gracile)、露籽草(Ottochloa nodosa)、狗脊(Woodwardia japonica)和酸模芒(Centotheca lappacea)等。
1.2 样地设计
为探索不同恢复年限红锥人工林植被的动态和恢复趋势,本研究采用空间代替时间的方法。在前期的调查和多次踏查的基础上,选择了具有代表性的7 个不同林龄序列的红锥人工林,分别为6、10、15、20、25、30 和34 a,且每个样地按以下3 个标准进行选择:(1)立地条件(包括地形和土壤)相对一致;(2)都是在皆伐地上发育的林地;(3)在皆伐之前,林地的树种均为马占相思人工林。所有样地的红锥人工林在生长的第3 和第5 年均进行了一定的人工抚育(包括施肥和去除林下植被),人工抚育完成以后即让林地自然生长,并停止人为干扰。每个林龄设置3 个20 m × 20 m 样方,共21 个样方。为了尽可能将空间自相关最小化,不同林龄样地间的距离至少1 km,以避免空间自相关,且所有的样地距林缘至少20 m 以减少林缘效应。所有调查均在2020 年8 月完成。对每块不同林龄的红锥人工林内选取的3 个样地(高度> 4 m 或胸径> 2 cm 的所有乔木)进行每木检尺,采用测高仪(SUUNTOPM 5/1520 PC)实测样方内树高,记录植株的种名、胸径、树高、生活力和干材质量等信息,调查灌木层和草本层的种类、株数、株高和盖度(表1)。
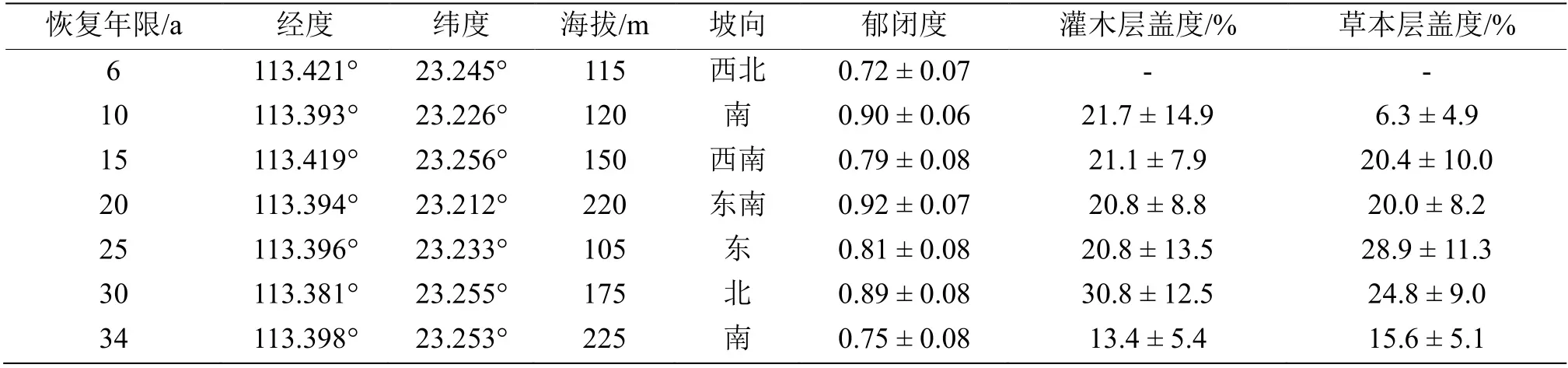
表1 样地基本信息Table 1 Information of sampling sites
1.3 多样性的测度
根据样地调查数据,分别计算各林龄阶段α 多样性指数,本研究采用物种丰富度(R)、Shannon-Wiener 指数(H')、Simpson 指数(D)和Pielou 指数(E)4 个常用指数测度物种多样性[7-8],具体公式计算如下:

式中,S为样方中物种数,Pi为群落中第i个物种的重要值。乔木层物种的重要值(important value,IV)用来反映各个物种在群落中的相对重要性,计算公式为:
重要值IV=(相对频度+相对密度+相对优势度)/3
1.4 数据处理与分析
结合二元立木材积表及胸径和树高,计算单木材积,然后相加推算林分蓄积量。采用LSD 多重比较分析了各参数(平均树高、平均胸径、林分密度、物种丰富度、林上树种组成、林下灌木覆盖度、林下草本覆盖度、胸高断面积、D2H、单株生物量和林分蓄积量等)在不同林龄阶段的差异显著性,显著性水平设定为α= 0.05。采用单因素方差分析(ANOVA)判断多样性、径级、材积和蓄积量的显著性差异。所有数据均采用Microsoft Excel 2020 和SPSS 22.0软件进行数据处理和分析,采用GraphPad Prism 8 进行作图。
2 结果与分析
2.1 不同林龄阶段群落组成
在广东省龙眼洞林场红锥人工林7 个林龄阶段的调查结果中,我们发现乔木层植物共有20 科25属27 种。其中,壳斗科(Fagaceae)、大戟科(Euphorbiaceae)、桑科(Moraceae)、芸香科(Rutaceae)、樟科(Lauraceae)、山茶科(Theaceae)和茜草科(Rubiaceae)植物比例较大,壳斗科物种重要值在不同林龄梯度的重要值最大。林分密度从6 a 到10 a 到林龄阶段显著上升,密度最大的两个林龄阶段为10 a 到20 a 分别为(1 391.7 ± 376.1)和(1 308.3 ± 364.3)株·hm-2(P< 0.05),但随着恢复年限的增加,在34 a 林龄阶段物种数又降低为(633.3 ± 80.4)株·hm-2(P< 0.05)(表2)。群落中的物种数从6 a 到10 a 林龄阶段后物种数开始增长迅速(增加4 种),最大值出现在15 a 林龄阶段(物种数为11 种),但随着恢复年限的增加,在34 a 林龄林龄阶段物种数又降低为6 种(表3)。6 a 到10 a林龄阶段基本为红锥纯林,重要值分别为100 和82.04,群落中仅有少量如三桠苦和黄桐等阳生灌木或小乔木,为典型的恢复初期群落特征。15 a 林龄阶段红锥重要值降至63.40,群落中出现了大量红花荷以及少量其他植物物种,表现为红锥-红花荷混交林。20 a 到25 a 林龄阶段红锥在群落中重要值有一定回升,红锥的重要值分别为70.33 和72.73,而此前大量出现的红花荷却消失殆尽,群落中开始出现如九节和银柴等,乔木物种相对较为单一。30 a 林龄阶段红锥重要值趋于减少,红锥的重要值为49.07,群落中除此前出现的九节和银柴外,出现了较多的鼠刺以及部分阴生灌木。34 a 林龄阶段红锥群落重要值再次出现增加,红锥的重要值为62.89,且上一林龄阶段出现的鼠刺退出群落,而九节和银柴的数量有一定的增加(表3)。

表2 样地林分特征Table 2 Characteristics of the studied stands (mean ± SD, n = 3)

表3 不同恢复年限各类树种的重要值分布Table 3 Tree species with important values of different restoration years
2.2 不同林龄阶段物种多样性特征
不同林龄阶段的乔木物种丰富度(R)在人工恢复后随恢复年限的增加呈现“M”型变化,其两个丰富度指数顶点分别出现在15 a 和30 a 阶段,分别为5.7 ± 3.8 和6.0 ± 1.0;乔木层物种的Simpson优势度指数和Shannon-Wiener多样性指数同样也呈现出“M”型变化,两个极大值均出现在15 a 和30 a阶段;乔木层物种的Pielou 指数虽也表现出类似前3 个指数的变化规律,但Pielou 指数在6 a 到10 a阶段表现出快速的上升趋势,此后10 a ~ 34 a 阶段之间虽有一定的波动,但却并不明显,这也是与前3 个指数变化的不同之处(图1)。
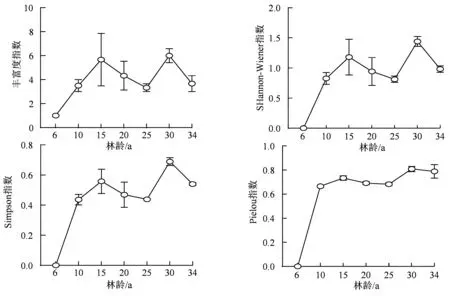
图1 不同林龄阶段α 多样性指数Figure 1 α diversity indexes in different forest ages
2.3 不同龄林阶段的生长动态
不同林龄阶段树木的平均胸径、平均树高和林分径级分布存在显著差异(表2)。随着恢复年限的增加,树木的平均胸径整体上表现出随恢复年限的增加先减小后增大的趋势,平均胸径最值出现在34 a林龄阶段,其次为6 a 林龄阶段,最小值出现在20 a林龄阶段,且在30 a 林龄阶段显著低于25 a 林龄阶段(P< 0.05),随后开始继续增大至34 a 林龄阶段(表2)。林分平均树高总体上呈现出随恢复年限的增加而增高的趋势,最大值出现在34 a 林龄阶段,其次为25 a 林龄阶段,最小值出现在6 a 林龄阶段,且在20 a林龄阶段显著低于15 a 林龄阶段(P< 0.05),30 a林龄阶段显著低于25 a 林龄阶段(P< 0.05)(表2)。林分胸高断面积则表现出先增加后减少再增加的趋势,其中最低值和最高值分别出现在6 a 林龄阶段和34 a 林龄阶段,且在20 a 林龄阶段显著低于15 a林龄阶段(P< 0.05),30 a 林龄阶段显著低于25 a 林龄阶段(P< 0.05)(表4)。6 a 林龄阶段中I 和II 径级的树木占个体总数的98%,随着恢复年限的增长,IV 径级的树木占比逐渐增大,在10 a 和15 a 林龄阶段分别达到13%和22%,而20 a 林龄林龄阶段却主要以II 径级树木为主,所占比例高达65%,IV 径级树木基本消失,直至25 a 林龄阶段开始重新恢复至个体总数的27%,并在此时林分中开始出现V 径级的树木,在30 a 和34 a 林龄阶段中,IV 径级的树木不断减少,在30 a 和34 a 林龄阶段间分别达到了10%和28%,V 径级的树木则没有明显的增加(表4)。
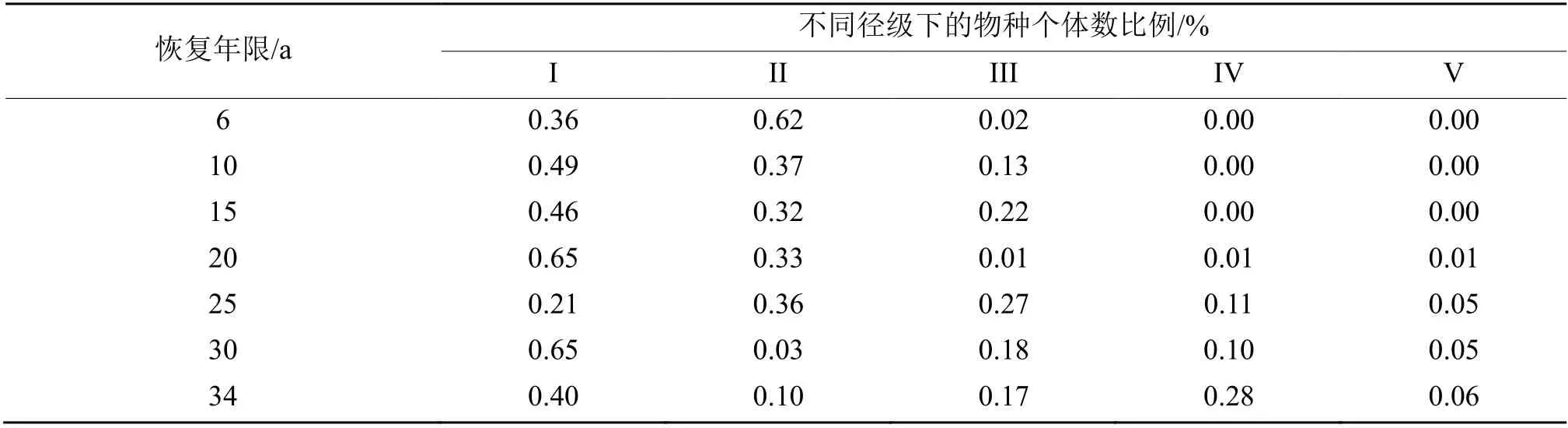
表4 不同恢复年限径级物种个体分布Table 4 BDH distribution of trees in plantations tree species with important values of different restoration years
2.4 不同龄林阶段的材积分布变化
不同恢复年限阶段林分D2H、单株生物量和林分蓄积量表现出显著差异,随着林分林龄的增长D2H、单株生物量和林分蓄积量呈现增加趋势,从6 a 到34 a 林龄阶段D2H 由(0.08 ± 0.00)m3显著增加到(1.19 ± 0.20)m3(P< 0.05),单株生物量由(42.51± 2.13) kg 显著增加到(379.79 ± 53.66)kg(P< 0.05),林分蓄积量由(49.95 ± 1.77)t·hm-2增加到(288.10 ±11.39)t·hm-2(P< 0.05),但30 a 林龄阶段却出现D2H、单株生物量和林分蓄积量的下降,分别为(0.66 ±0.11)m3、(218.36 ± 32.38)kg 和(227.46 ± 4.50)t·hm-2,并在34 a 林龄阶段时重新表现为增加(表5)。林分蓄积量的增加主要发生在20 a 至25 a 和30 a 至34 a这两个林龄阶段(P< 0.05),且单株生物量的增加幅度要大于蓄积量的增加,这也反映出林分密度在不同林龄阶段的变化为先上升后下降,最后整个林分仅表现在单株生物量的增加,植物株数变化不明显(表5)。
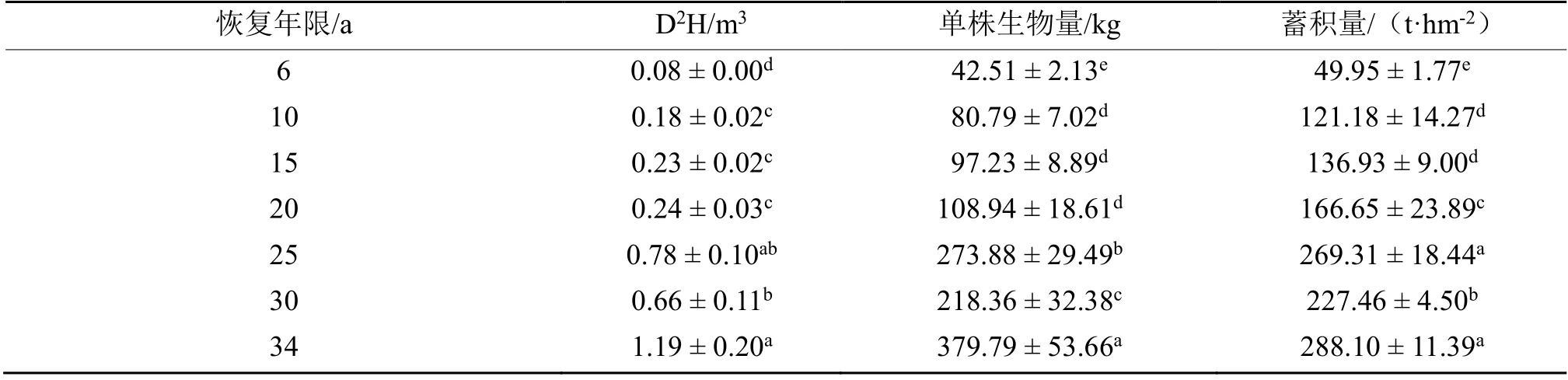
表5 不同恢复年限红锥人工林样地树种的材积与蓄积量Table 5 Volume growth of the the trees with different restoration years in the observational plots
3 讨论
在广东省龙眼洞林场红锥人工林7 个林龄的调查结果中,我们发现乔木层植物20 科25 属和27种,其物种组成较少,群落结构较为单一。主要是壳斗科、大戟科、桑科、芸香科和茜草科植物比例较大,壳斗科物种重要值在不同林龄梯度的重要值最大,为主要建群树种。Steffan-Dewenter 等[6]指出物种多样性和生态系统功能在扰动后的恢复过程中可能呈现非线性关系。本研究发现:不同林龄阶段的乔木物种丰富度(R)、Simpson 优势度指数和Shannon-Wiener 多样性指数同样也呈现出“M”型变化,两个极大值均出现在15 a 和30 a 阶段;乔木层物种的Pielou 指数虽也表现出类似前3 个指数的变化规律,但Pielou 指数在6 a 到10 a 阶段表现出快速的上升趋势,此后10~34 a 阶段之间虽有一定的波动,但却并不明显,这与姜俊等[30]的研究结果类似。有研究发现,随着林龄的增长,群落逐渐由南亚热带乡土树种常树种入侵[27]。由于该地区位于我国南亚热带地区,气候的顶级群落为南亚热带常绿阔叶林(季风常绿阔叶林)[28],因此,推测该区域红锥人工林的林龄发展阶段可能为:从初期的红锥人工纯林到中期的红锥-常绿阔叶混交林再到最终的南亚热带常绿阔叶林,此红锥林龄序列在南亚热带地区的森林恢复过程中较为常见[29-30]。事实上,从红锥人工林到南亚热带常绿阔叶林的发展过程中,从红锥人工纯林到出现红锥-常绿阔叶混交林的间隔并不长,也许只有15 a 甚至更短,但红锥-常绿阔叶混交林的存在可能超过百年的时间,因为随着林龄的增加,红锥不断衰老死亡以及自我更新的周期较长导致其优势度逐渐降低。红锥-常绿阔叶混交林逐渐在林冠层占优,但物种多样性恢复尚需一段时间。随着年龄的增长,林木占据的资源空间变大,种子库中的实生苗迅速萌发,促进了林下灌木层乔木幼苗的繁衍和扩张。不同林龄阶段红锥人工林群落的多样性变化是环境条件、竞争关系以及区域过程共同影响的结果[31-32]。
研究发现:随着恢复年限的增加,树木的平均胸径整体上表现出随恢复年限的增加先减小后增大的趋势,平均胸径最大值出现在34 a 林龄阶段,其次为6 a 林龄阶段,最小值出现在20 a 林龄阶段;林分平均树高总体上呈现出随恢复年限的增加而增高的趋势,最大值出现在34 a 林龄阶段,其次为25 a 林龄阶段,最小值出现在6 a 林龄阶段(表2)。林分胸高断面积则表现出先增加后减少再增加的趋势,其中最低值和最高值分别出现在6 a 林龄阶段和34 a 林龄阶段(表4)。究其原因可能是因为恢复初期(6 a)由于定值的都是统一规格,基径树高基本一致的红锥幼苗,随着恢复年限的增加,新长出的乔木(高度> 4 m 或胸径> 2 cm)数量增多,致使林分的平均树高、平均胸径和胸高断面积降低或增加幅度降低,之后随着原有红锥林的不断生长逐渐发育成为大径材以及新生乔木的不断生长,使得林分的平均树高、平均胸径和胸高断面积重新回升,林分密度(表2)和林分大径级树木比例(表4)可以佐证这一推论。此外,我们发现,不同恢复年限阶段林分D2H、单株生物量和林分蓄积量表现出显著差异,随着林分林龄的增长D2H、单株生物量和林分蓄积量呈现增加趋势,从6 a 到34 a 林龄阶段D2H、单株生物量和林分蓄积量均显著增加(P<0.05),但30 a 林龄阶段却出现D2H、单株生物量和林分蓄积量的下降,并在34 a 林龄阶段时重新表现为增加(表5)。由于林分材积的增加主要发生在20 a~25 a 和30 a~34 a 这两个林龄阶段,且单株生物量的增加幅度要大于蓄积量的增加,这反映出林分密度在不同林龄阶段的变化为先上升后下降,最后整个林分仅表现在单株生物量的增加,植物株数变化不明显,也就是说在恢复过程中林分发生自疏现象。
针对我国人工林经营中存在的问题和新时期社会对森林期望与需求的新变化,我国人工林经营发展战略和对策在全球气候变化大背景下正在发生改变和调整。从单纯的注重人工林面积扩张逐渐转变为人工林生态系统服务质量和效益提升,发展可持续的多功能人工林已经渐渐成为一种主流趋势[16]。从森林经营和管理的角度看,我们需要利用人工林生态系统内部自然生长规律,根据不同的林龄阶段和林相来实施不同的经营策略[35]。以广东省龙眼洞林场不同林龄红锥人工林为例,10 a 林龄阶段是林分的初始阶段,主要特点是林冠密闭,到15 a 林龄阶段时早期入侵种的林下植被和小径级(胸径小于10 cm)的树木数量逐渐增加。该阶段是群落物种组成和结构变化的开始,由对光需求的物种和林冠下更新所驱动的[34]。因此,在林分初始阶段应适当筛选出适地适树改造的乡土物种和目标树种,依据更新树种对光照、土壤和生物学特性,采取人工补植和播种等调整混交比例等措施,改善林分的树种组成和结构[32]。其次,20 a 林龄阶段既是林分密度较高的阶段,也是林下植被覆盖度最高的阶段,短寿命的植物死亡率的增加和慢生优势树种分化,开始出现红锥-常绿阔叶混交林的林相,但分布的是干型细长且数量多的幼树,缺乏光照且自身竞争力较弱,更多的林木逐渐转向径向生长,较大胸径的林木个体比例增加,对资源的竞争会逐渐加剧[35]。因此,建议该阶段林分上层疏开,以改善林内的光照条件并缓解个体间竞争压力。最后,30 a 林龄阶段以后主要特征是红锥幼苗和占据优良生态位的红锥数量相当,耐阴树种和长寿命树种占据上层,结构复杂、物种丰富以及林下植被更新萌蘖能力强,主要原因是光照条件的改善和竞争效应的减弱使林冠的活力及其光合作用得到增强,促进了近熟林阶段林木的迅速生长和物质积累[36],对于南亚热带常绿阔叶林的过渡起到了重要作用,成为在整个林分发育进程里重点经营阶段。因此,建议该阶段林木应避免生长趋势良好的母树受灾害和机械损伤影响,在选择目标树后,每10 a 林龄阶段一次促进目标树生长抚育间伐,直到达到目标直径的树木,在获得森林收获物质的同时又保证林分稳定性,以实现林地持续发挥森林生态服务功能[37]。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
农业工程学报(2022年10期)2022-08-22
现代园艺(2022年6期)2022-04-08
安徽农学通报(2020年7期)2020-05-26
农民致富之友(2020年4期)2020-04-03
农业与技术(2017年9期)2017-05-28
安徽农学通报(2017年6期)2017-04-11
绿色科技(2017年3期)2017-03-14
安徽农学通报(2014年18期)2014-10-23
安徽农学通报(2014年9期)2014-06-23
