
长江中游宜昌段浮游动物群落结构特征与环境因子相关性分析
2022-10-22 02:55伍遇普田波蔡志宇吴益平黄盼君刘千凡周志国
黑龙江水产 2022年5期
向 浩,林 枫,伍遇普,田波,蔡志宇,吴益平,黄盼君,刘千凡,周志国
(1.长江湖北宜昌中华鲟自然保护区管理处,湖北宜昌 443000;2. 武汉水科苑科技有限公司,湖北武汉 430000;3.湖北省漳河工程管理局,湖北荆门 448156 )
水生态系统动态平衡是生物群落与环境因子之间相互影响和制约的结果,水生生物群落结构受水环境变化影响,水生生物群落结构及动态变化也会反映出水体环境变化[1-2]。浮游动物是一类悬浮于水中营浮游性生活的微型动物(原生动物、轮虫、枝角类和桡足类),具有体型微小、数量多和代谢活动强等特点[3-4]。浮游动物主要以细菌、碎屑和浮游植物为食,属于水生生态系统食物链中的初级消费者,同时又是其他高级营养级水生生物的天然饵料。浮游动物是联系初级生产者和次级消费者的重要一环,在生态系统中物质循环和能量流动过程中起到承上启下的作用[5],同时对于维持食物网丰富性和稳定性具有重要的调控作用[6-8]。
宜昌位于湖北省西南山区与江汉平原的过渡地带,属亚热带季风性湿润气候,水资源丰富,大小河流和水库占全境面积5.6%,孕育了丰富的水生生物资源[9]。近年来,宜昌段对水生生物的研究大多集中在渔业资源[10-13],对浮游动物的生物多样性有过研究,但时间较早,且采集的周期较短[14],对浮游动物群落结构及环境因子有关研究未见报道。因此,为填补宜昌段浮游动物群落结构特征及与环境因子相关性等方面研究空白,该研究在宜昌江段进行了为期1年的野外采样调查,分析了浮游动物群落结构特征及其时空变化;结合水质分析数据,并通过相关分析和冗余分析(RDA)研究了浮游动物群落结构与环境因子的相关关系,以期为长江宜昌段生态系统变化研究积累资料。
1 材料与方法
1.1 采样断面
在长江中游宜昌段设置了11个调查断面(图1),顺水流方向从上至下以依次为:柏临河口(BLHK)、磨盘溪(MPX)、古战场(GZC)、工贸码头(GMMT)、唐家大湾(TJDW)、清江口(QJK)、白洋镇(BYZ)、枝城大桥(ZCDQ)、车阳河(CYH)、百里洲(BLZ)和七星台(QXT),分别于2021年春季(4月)、夏季(7月)、秋季(9月)和冬季(12月)进行了浮游动物群落与生境调查。

图1 长江中游宜昌段采样断面示意图
1.2 样品采集与处理
参照《内陆水域渔业自然资源调查手册》,在各采样断面分左岸、河中、右岸3个采样点进行浮游动物的定性和定量样品采集。原生动物和轮虫的定性样品采用25#浮游生物网在采样断面水面下10cm处呈“∞”形拖拽采集,拖动速度不超过0.3m/s;定量样品则采用5L有机玻璃采水器,在上、中、下水层取混合水样1L,加入1.0%~1.5%鲁哥氏液固定,定置沉淀24hrs,浓缩至 30mL。
枝角类和桡足类定性样品则采用13号浮游生物网采集,采集的样品用4%~5%甲醛溶液固定;定量样品用采水器在水面上、中、下三层处取 20L 混合水样,用 25#浮游生物网过滤浓缩至 50mL,现场加入4%~5%甲醛溶液固定。采集的样品运送至实验室,沉淀浓缩后在显微镜下进行鉴定和计数。样本的鉴定及生物量的换算参考《中国淡水藻类》《中国淡水生物图谱》和《淡水浮游生物研究方法》等。
采样现场采用GPS记录经纬度,使用塞氏透明度盘测定透明度(SD);使用便携式水质分析仪(YSI)测定水温(Water Temperature,WT)、电导率(Conductivity,Cond)、pH和溶解氧(Dissolved Oxygen,DO);使用流速仪(FP111)测定流速(CV)。总氮(Total Nitrogen,TN)、总磷(Total Phosphorus,TP)等水质参数测定参照《水和废水检测分析方法》进行。
1.3 数据处理与分析
对长江中游宜昌江段浮游动物的Shannon-Wiener多样性指数(H)、Margalef种类丰富度指数(D)、Pielou均匀度指数(J)及优势度(Y)进行计算。相关计算公式如下:

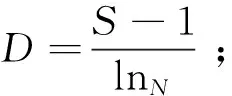
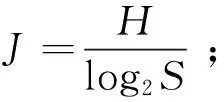

式中:Ni为第i种的个体数;N为样品中的总个体数;fi为第i种在各样点出现的频率;S为种类数。取优势度Y≥0.02的物种作为优势种。
对浮游动物的丰度、生物量、Shannon-Wiener 指数(H)、Pielous 均匀度指数(J)、Margalef 丰富度指数(D)以及TN、TP、pH和DO等,进行标准差标准化处理后,采用SPSS 26.0软件对浮游动物与环境因子做Pearson相关性分析,其中R值表示相关性大小,P值表示两个变量间的显著性,一般要求P<0.05有统计学意义。
采用 Canoco 4.5 软件对浮游动物丰度和环境因子进行数据分析。先对浮游动物相关数据进行去趋势对应分析(Detrended correspondence analysis,DCA),当排序轴最大梯度长度大于3 时选择典范对应分析(canonical correspondence analusis,CCA),当排序轴最大梯度长度小于3时选择冗余分析(Redundancy analysis, RDA),最终用物种与环境因子的双序图表示浮游动物与环境因子的关系[15]。采样点的绘制结合使用 ArCIS10.8 和 BigMap完成。
2 结果
2.1 水质理化指标
各调查断面水体理化指标显示水体环境因子呈现明显的季节变化(表1)。2021年采样断面水温季节变化范围14.53℃~24.68℃,年均水温19.67℃,四季水温差异较大,夏季最高,冬季最低。采样断面流速季节变化范围为0.23m/s~0.65m/s,夏季最大,春季最小,平均流速0.41m/s。水体总体呈弱碱性,pH值范围为7.93~8.25,平均pH为8.11,季节差异不明显。电导率的范围为0.16ms/cm~0.34ms/cm,平均电导率为0.25ms/cm,季节差异显著(P<0.05),其中冬季最高为0.34ms/cm,夏季最低为0.16ms/cm。溶解氧的季节变化表现为秋季最高,夏冬季节次之,春季最低,平均溶解氧为8.66mg/L。水体 TP在夏季明显低于其他季节,其他季节差异不明显。水体TN秋季最高,其他季节差异不大;COD四季变化不明显。
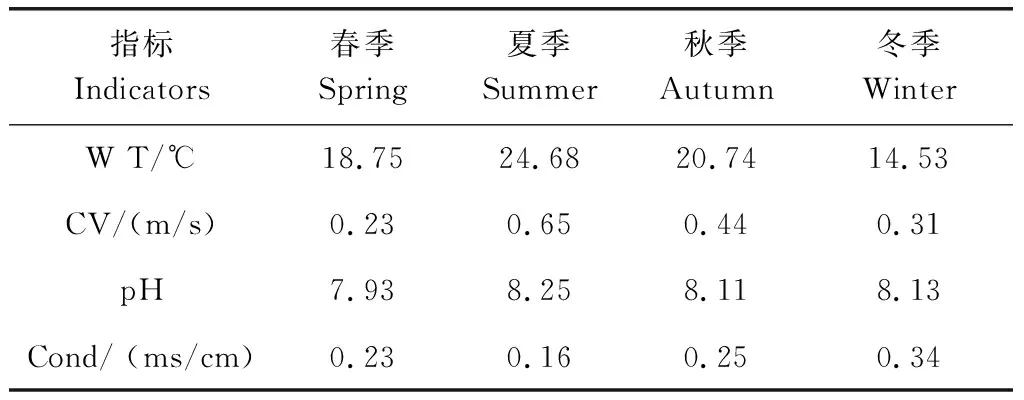
表1 水体理化指标季节变化(均值)

续表
2.2 种类组成及优势种
对11个断面不同季节采集的样本进行鉴定,结果显示:2021年共鉴定出浮游动物71种,其中轮虫类种类数最多,为28种,约占总数的39.4%;其次为桡足类的16种,约占总数的22.5%;枝角类15种,约占总数的21.1%;原生动物种类数最少,为12种,约占总数的16.9%(图2)。

图2 浮游动物种类组成
从季节上来看,浮游动物种类组成的季节变化差异较明显,夏季种类数最多,为62种,其次为秋季和春季,分别为49种和44种,冬季种类数最少为40种(图3)。

图3 不同季节浮游动物种类组成
经统计分析,采样断面浮游动物优势种共有15种(Y>0.02)(表2)。其中,原生动物2种,包括球形砂壳虫(Difflugiaglobulosa)和叉口砂壳虫(Difflugiagramen);轮虫6种,包括萼花臂尾轮虫(Brachionuscalyciflorus)、壶状臂尾轮虫(Brachionusurceus)、角突臂尾轮虫(Brachionusangularis)、剪形臂尾轮虫(Brachionusforficula)、螺形龟甲轮虫(Keratellacochlearis)和矩形龟甲轮虫(Keratellaquadrata);枝角类3种,包括短尾秀体溞(Diaphanosomabrachyurum)、长额象鼻溞(Bosminalongirostris)和直额弯尾溞(Camptocercusrectirostris);桡足类4种,包括广布中剑水蚤(Mesocyclopsleuckarti)、透明温剑水蚤(Thermocyclopshyalinus)、闻名大剑水蚤(Macrocyclopsdistinctus)和汤匙华哲水蚤(Sinocalanusdorrii)。
从季节上来看,夏季优势物种种类数最多为7种,冬季优势种最少为1种,数量大小顺序为夏季(7种)>秋季(5种)>春季(3种)>冬季(1种)(表2)。

表2 浮游动物优势种季节变化
2.3 密度和生物量变化
各采样点浮游动物的密度变化范围为176.4ind./L~1703.0ind./L,平均密度为674.1ind./L(表3)。从类群上来看,轮虫的密度最大(1497.2ind./L),其次为原生动(1006.9ind./L),枝角类最小(86.6 ind./L)(表4)。从季节上来看,浮游动物密度不同季节之间存在较大差异(图4-a),密度大小顺序总体为:夏季>秋季>春季>冬季;其中,夏季的浮游动物平均密度最高为1161.8ind./L,显著高于春、冬季的425.2ind/L和268.8ind/L(P<0.05);春季和冬季之间的差异性不显著。从空间上看,浮游动物密度分布也存在较大差异,各个季节总体表现为下游江段相对较大,而上游江段相对较小。
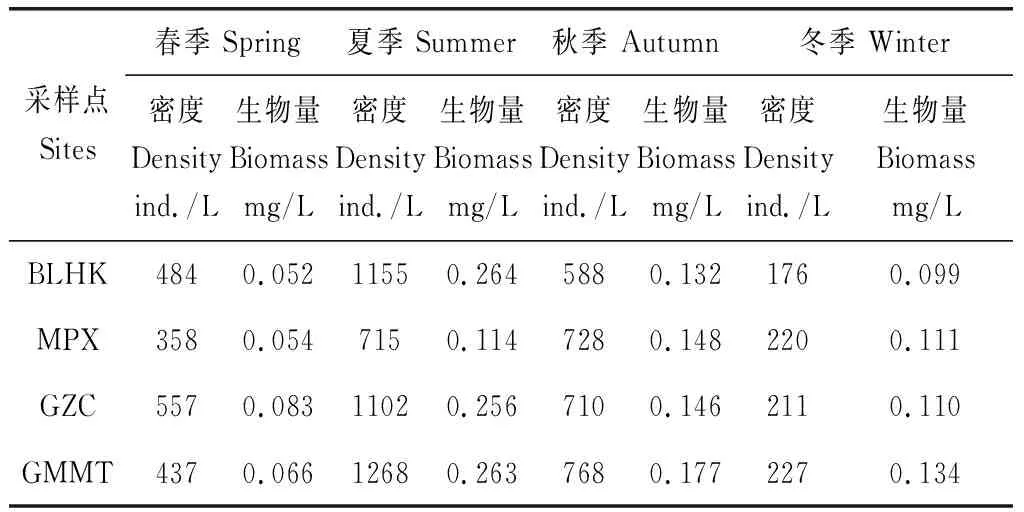
表3 各采样点不同季节浮游动物密度和生物量

续表
各采样点浮游动物生物量范围为 0.052mg/L~0.365mg/L,平均生物量为0.154mg/L。从类群上来看,枝角的生物量最大(0.34mg/L),其次为桡足类(0.22mg/L),原生动物最小(0.01mg/L)。从季节上来看,各采样点生物量季节变化趋势与密度基本一致(图4-b),即:夏季>秋季>春季>冬季。夏季的浮游动物平均密度最高为0.232mg/L,显著高于春的0.085mg/L(P<0.05)。从空间上看,宜昌江段下游生物量相对较大,上游江段相对较小,与密度变化趋势一致。

图4 浮游动物密度(a)和生物量(b)季节变化

备注:a代表春季;b代表夏季;c代表秋季;d代表冬季
2.4 生物多样性特征
对11个采样断面浮游动物生物多样性进行统计(见表5和图6),结果表明:2021年宜昌江段各断面浮游动物Shannon多样性指数(H)变化范围为 0.95~4.25,年均值为2.35;H季节变化差异较大,其中夏季H值与冬季H值之间存在显著性差异(P<0.05),冬季与春季之间无显著性差异。Pielou 均匀度指数(J)的波动范围为 0.25~0.77,年均值为0.45,但季节之间变化差异不显著。Margalef指数(D)范围为0.11~1.94,年均值为0.93,季节变化之间差异性不显著。不同断面上,Shannon多样性指数(H)、Pielou 均匀度指数(J)和Margalef指数(D)差异较大,总体上下游江段各指数值大于上游江段。

表5 长江中游宜昌段各样点4个季节多样性指数

备注:a代表春季;b代表夏季;c代表秋季;d代表冬季
2.5 浮游动物与环境因子分析
对浮游动物密度、生物量和多样性指数等与环境因子进行Pearson 相关性分析,结果表明:浮游动物的密度和生物量,与温度、pH之间呈显著的正相关关系(P<0.05),水温和pH对浮游动物的生长与繁殖起着重要的调节作用;与流速呈显著负相关(P<0.05),在流速较快的水域,浮游动物密度和生物量较低;与总氮、总磷和溶解氧呈正相关,与电导率呈负相关,但相关性均不显著。浮游动物的Shannon多样性指数(H)与流速和溶解氧呈正相关,与其他环境因子呈负相关,但是相关性均不显著;Margalef种类丰富度(D)与电导率呈显著负相关(P<0.05),与其他因子之间关系不显著;Pielou均匀度指数(J)与水温呈负相关,与pH、流速等其他因子呈正相关,但均不显著(表6)。

表6 宜昌江段浮游动物与环境因子相关性分析
对15种优势物种进行去趋势对应分析(DCA),排序轴最长梯度小于3,说明浮游动物群落结构接近线性模型,该研究采用冗余分析(RDA)探究浮游动物与水体理化指标(环境因子)的关系。结果显示,温度、pH、流速与浮游动物群落结构密切相关。其中水温、pH和TN与第一排序轴为正相关;水温与第一排序轴为最大正相关,流速与第二排序轴呈最大负相关。就优势物种与环境因子关系而言,球形砂壳虫、叉口砂壳虫、壶状臂尾轮虫、长额象鼻溞、直额弯尾溞、广布中剑水蚤与pH、水温呈正相关,与电导率及流速呈负相关;角突臂尾轮虫、矩形龟甲轮与总氮和总磷呈正相关关系(图7)。
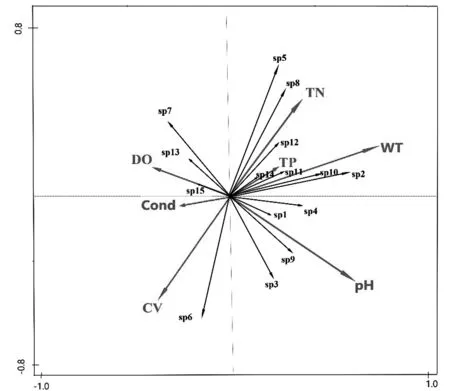
备注:SP1为球形砂壳虫;SP2为叉口砂壳虫;SP3为萼花臂尾轮虫; SP4为壶状臂尾轮虫;SP5为角突臂尾轮虫;SP6为壶状臂尾轮虫;SP6为剪形臂尾轮虫;SP7为螺形龟甲轮虫;SP8为矩形龟甲轮虫;SP9为短尾秀体溞;SP10为长额象鼻溞;SP11为直额弯尾溞;SP12为广布中剑水蚤;SP13为透明温剑水蚤;SP14为闻名大剑水蚤;SP15为汤匙华哲水蚤。
3 讨论
3.1 浮游动物群落结构特征
长江宜昌段位于中亚热带与北亚热带的过渡地带,属亚热带季风性湿润气候,浮游动物种类为亚热带水体常见种。研究期间,共鉴定出浮游动物4门71种,其中轮虫类种类数最多(总数39.4%);其次为桡足类(占总数22.5%),枝角类(占总数21.1%);原生动物种类数最少(占总数16.9%)。调查结果显示,宜昌段浮游动物主要以轮虫为主,与长江干流及大多数河流浮游动物研究结果一致,具有典型的河流浮游动物群落结构特征[14,15-17]。这与轮虫的生物学特性有关,轮虫独特的孤雌生殖方式使其能在短时间内进行大量繁殖,加上轮虫适应能力很强,能够在河流水生态系统中快速占据优势地位[18];轮虫类以浮游植物和有机碎屑等作为食物来源,宜昌段轮虫类食物来源丰富,导致轮虫种类和数量较多。该研究与2013年[14]的调查结果相比,浮游动物的种类数差异不大(分别为71种和69种),但是采集到的物种差异较大,本次调查轮虫种类和枝角类数增加,而原生动物种类数减少,这可能与调查范围、调查周期长短等差异有关。
从季节角度来看,浮游动物种类组成的季节变化差异较明显,夏季种类最多(62种);其次为秋季(49种)、春季(44种);冬季种类数最少(40种),这说明同一河流不同季节受到不同因素的影响,浮游动物群落结构存在差异。从水平分布来看,中下游采样断面的浮游动物种类数高于上游种类数,这可能与流速有关。根据水体理化因子监测结果,宜昌段上游采样断面接近葛洲坝,河道较窄,流速较高,中下游河段河道变宽、流速缓慢。浮游动物主要以细菌及藻类为食,浮游动物的分布与细菌及藻类等密切相关,而细菌及藻类的主要受流速的影响[19]。有关研究表明,葛洲坝截流后水的流速减缓,有利于细菌、藻类等的繁殖,浮游动物的密度和多样性均显著增加[20],这与该研究中浮游动物生物量和多样性与流速呈显著负相关的结果一致。
3.2 浮游动物密度和生物量变化
浮游动物密度和生物量的变化大致可以反映出生物群落的变化[21]。研究结果表明,轮虫的密度最大为1497.2ind./L,占总量的55.51%;其次是原生动物,为1006.9ind./L,占总量的37.33%;桡足类和枝角类密度较小,分别为86.6ind./L和106.5ind./L,共占总量的7.16%。原生动物和轮虫体型较小,较体型比较大的桡足类和枝角类而言,其相对湿质量偏低,导致该类群密度与生物量在占比上存在不一致性。轮虫具有孤雌生殖特性,能够在较短时间内完成生活史,并且适应能力很强,使其在能够在河流水生态系统中快速占据优势地位,因此该研究中轮虫的密度较大。
调查结果显示,浮游动物密度和生物量季节之间存在显著性差异,大小顺序总体为:夏季>秋季>春季>冬季,夏季的浮游动物平均密度最高(1161.8ind/L),显著高于春、冬季的平均密度(425.2ind/L和268.8ind/L)(P<0.05),可能是因为夏季浮游植物生物量的提高为浮游动物提供了充足饵料。浮游动物密度和生物量在水平分布上也存在较大差异,各个季节总体表现为下游江段相对较大,而上游江段相对较小。
3.3 浮游动物群落与环境因子关系
在河流生态系统中,浮游动物群落结构及时空分布受水温、浮游植物、营养盐的上行效应和鱼类捕食的下行效应以及种内竞争等因素的影响,是多种环境因子在时间和空间序列上共同作用的结果[22-23]。郭欧阳[24]研究认为,水温、溶解氧、总氮、总磷、盐度、总溶解性固体是长江下游干流浮游动物群落结构的主要影响因素;郭杰等[16]研究认为,温度、溶氧和氨氮是影响长江中游浮游动物群落结构的主要因素。同一河流影响浮游动物的环境因子出现异同,除了与河流的水文、水物理、水化学的变化以及人为干扰程度有关外,还与采样地点、采样时间、物种鉴别水平等存在差异有关[25]。
Pearson 相关性和RDA结果表明,水温对宜昌江段的浮游动物群落结构具有显著影响,与浮游动物种多样性、丰度、生物量呈显著性正相关。相关研究表明,水温升高不仅提升浮游动物的饵料生物浮游植物密度和生物量,也会提高浮游动物的摄食效率,是影响浮游动物群落结构的重要环境因子[26]。该研究显示,宜昌段浮游动物的密度和生物量存在季节性差异,这可能与宜昌段属于亚热带季风性湿润气候有关,夏秋季节具有良好的光照和水温条件,而春冬季温度相对较低,宜昌水温条件的季节差异为浮游动物群落结构的季节性变化创造了条件。此外,宜昌段冬春季水温偏低,夏秋季水温较高,因此夏季的密度及生物量均高于其他季节。
浮游动物的种类、数量变化与也与水体pH值密切相关。高原等[27]研究表明,轮虫和桡足类适宜生活在偏碱性水体中,枝角类则在酸性、中性及微碱性水体中生活。Yin 等[28]对轮虫在酸碱水体中的适应研究认为,酸性水体中轮虫种类多、数量少,在碱性水体中则相反。调查期间,各采样断面的 pH值范围为7.93~8.25,平均pH为8.11,属于典型的弱碱性水体,适合轮虫和桡足类等生存繁衍。宜昌段夏季浮游植物等光合作用增强,溶解态CO2大量消耗使水体酸碱平衡向碱性偏移,导致夏季pH值较高。该研究表明,宜昌段浮游动物的密度和生物量与 pH 呈显著的正相关,夏季pH值最高,浮游动物的密度及生物量最大。
浮游动物游泳能力相对较弱,不能有力地抵抗水的冲击,喜欢在静止或流速较小的水体中生活。Saunders等[29]研究表明,流速增大,浮游动物的种类数及生物量减少。该研究显示,随着夏季降雨增多,水流增大,流速与浮游动物种类丰度存在负相关性,与Saunders等[29]研究结果相似。溶解氧和氮磷营养盐也是影响浮游动物群落特征的重要因素,氮、磷等营养盐不会对浮游动物群落结构造成直接影响,但会通过调节浮游植物而间接影响浮游动物群落[30]。该研究结果显示,溶解氧和氮磷等营养盐与浮游动物密度和生物量呈正相关,但相关性不显著。
猜你喜欢
党员生活·下(2022年1期)2022-04-23
科学养鱼(2020年10期)2020-11-23
学生天地(2020年31期)2020-06-01
科学养鱼(2020年12期)2020-04-22
课外语文·中(2020年2期)2020-03-24
智富时代(2019年8期)2019-09-23
智富时代(2019年8期)2019-09-23
小天使·一年级语数英综合(2019年8期)2019-08-27
知识窗(2019年4期)2019-04-26
小学阅读指南·低年级版(2016年6期)2016-05-14
