
鸡IGF1R基因遗传变异与其体尺和屠宰性状的相关性
2022-10-21 11:13杨礼杨蛟陈磊杜勇张胜国牟慧蓉冉隆权张依浴
畜牧兽医科学 2022年7期
杨礼,杨蛟,陈磊,杜勇,张胜国,牟慧蓉,冉隆权,张依浴
(1.贵州省沿河县农业农村局,贵州 铜仁 565300;2.贵州省铜仁市农业农村局,贵州 铜仁 554300;3.贵州大学,贵州 贵阳 550000)
0 引言
世界各地的许多鸡品种不仅需要通过选择培育去提高它的生长特性,而且要提高屠宰和肉质属性。传统的育种方法得到了各种分子遗传学工具的支持,以提高生产性状的遗传增益率,加速育种目标的实现。在标记辅助选择中,使用对这些特性具有显著和积极影响的适当分子标记可以改善胴体和肉质性状。
胰岛素样生长因子1受体(Insulin-like growth factor receptor 1,IGF1R)是一种酪氨酸激酶受体,由异四聚体跨膜糖蛋白的两个细胞外α亚基(135 kDa)和2个跨膜β亚基(90 kDa)组成[1-2]。IGF1R属于细胞调节剂家族,由3种激素组成,包括胰岛素样生长因子1(Insulin-like growth factor 1,IGF1),IGF2和胰岛素(Insulin),以及另一种称为胰岛素样生长因子2受体的受体(IGF2R)和6个胰岛素样受体生长因子结合蛋白(Six insulin-like growth factor binding proteins,IGFBP1-6)[3-4]。这些蛋白质充当内分泌、自分泌和旁分泌刺激物,在胚胎发生、细胞生长和分化、有丝分裂、存活和代谢调节中起重要作用[5]。当IGF1,IGF2或胰岛素中的任何一种与受体结合,就会发生IGF1R信号,而这种信号可介导并促进肌肉肥大的途径,抑制有利于肌肉萎缩的生物途径,进而对动物的生长发育起调控作用。由于具有上述功能,IGF1R基因已被认为多种动物生长和胴体性状的功能候选基因[6]。
在鸡体内,IGF1R基因定位于10号染色体上,含有21个外显子和20个内含子,跨域基因组140 315个bp(NCBI RefSeq:NC_006097.5,区域范围为17,124,349-17,264,663)编码一个900个氨基酸多肽(NCBI RefSeq:NM_205032.1)。目前有关IGF1R基因对鸡的体尺性状和大多数屠宰性状的影响的相关研究较为匮乏,特别是以贵州沿河铁叫鸡为研究动物的研究信息则不存在。本研究目的是进行IGF1R基因的多态性调查,并且评估其多态性及其基因型对鸡体尺性状和屠宰性状的影响,以期为鸡的科学选育培育提供参考。
1 材料与方法
1.1 试验动物
研究在贵州省沿河县铁叫鸡育种场选取同批出壳、健康无病和同一饲养环境下的20周龄鸡100只,每只鸡翅静脉采取血液2 mL用于总DNA的提取。
1.2 体重与体尺测定
在鸡禁食12 h后称量其活重,按照NY/T823—2004规定测定体尺[7]。
1.3 屠宰性能测定
在测定完体尺指标后将鸡颈静脉放血致死,利用湿拔毛法,根据NY/T823—2004规定测定沿河铁叫鸡的屠宰性状。
1.4 引物的设计和基因组DNA提取
按照大连宝生物血液基因组DNA提取试剂盒(TaKaRa,Dalian,China)说明书提取基因组DNA。根据NCBI GeneBank已公布鸡IGF1R基因(NCBI RefSeq:NC_006097.5)序列。Primer 3(美国Applied Biosystem公司)设计多态引物[8],由上海生工生物工程股份有限公司合成,见表1。

表1 试验所用引物
1.5 DNA池PCR扩增及测序分型
PCR反应体系30.0 mL:15.0 mL PCR MasterMix,正、反向引物各2.0 mL,DNA模板1.0 mL,12.0 mL ddH2O。按预变性、退火、延伸、终延伸程序进行PCR扩增,产物经检测后送至生工生物工程(上海)股份有限公司测序。结果经比对分析,筛选出单核苷酸多态性(SNP)位点。通过PCR扩增获取产物继续送至生工生物工程(上海)股份有限公司测序,进行基因分型。
1.6 统计分析
在线软件SHEsis(http://analysis.bio-x.cn/)构建单倍型和双倍型;PopGene 软件(ver.1.32)计算等位基因频率和等位基因型;PowerMarker 软件(ver.3.25)对多态信息含量(PIC)进行分析;采用PASW Statistics 18.0软件对屠宰性能和体尺测定数据进行有效性检测,剔除不符合正态分布个体。一般线性模型(GLM)分析屠宰性能和体尺与IGF1R基因的关联性,差异显著(P<0.05)。
2 结果与分析
2.1 鸡IGF1R基因多态性分析结果
使用MegAlign分析鸡IGF1R基因多态性,发现在鸡IGF1R基因的第3外显子(Exon-3)有3个SNPs位点,即g.105384 C>T、g.105333 A>C和g.110681 A>C。3个SNPs位点均存在3种基因型,突变点为同义突变,编码氨基酸未改变。见图1、图2、图3。
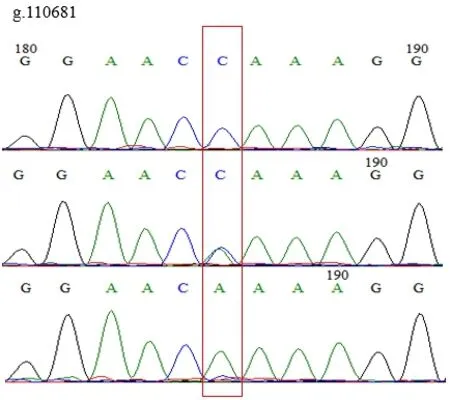
图1 g.110681
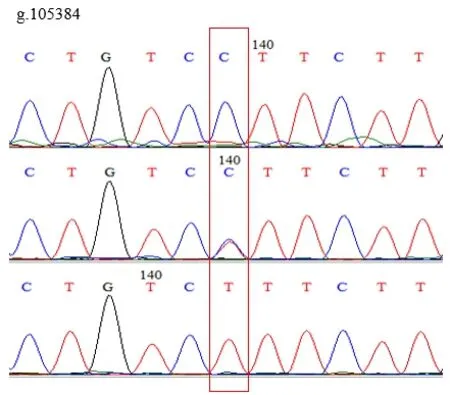
图2 g.105384
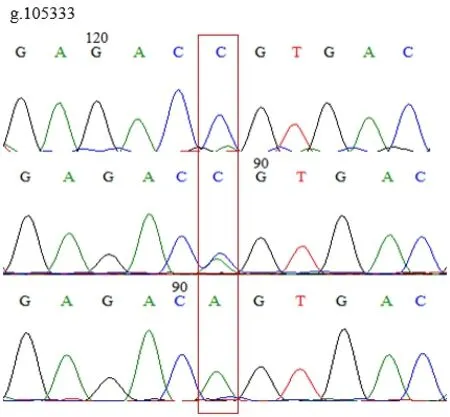
图3 g.105333
2.2 鸡IGF1R基因群体遗传特性分析
鸡IGF1R基因群体遗传特性分析见表2。g.105384 C>T、g.105333 A>C和g.110681 A>C这3个SNP位点均为中度多态(0.25<PIC<0.5);优势基因型分别为CT、AC和AC,频率均是0.580;优势等位基因和频率分别为C(0.647)、C(0.647)、A(0.735)。χ2检验结果显示,g.105384 C>T、g.105333 A>C和g.110681 A>C均未偏离Hardy-Weinberg平衡(P>0.05)。
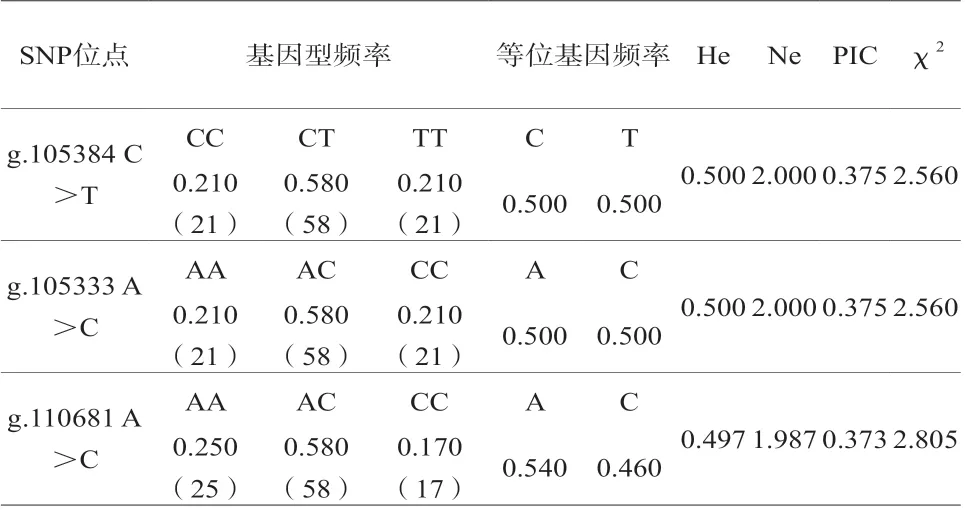
表2 SNP位点群体遗传信息分析
2.3 鸡IGF1R基因SNP单倍型和双倍型分析结果
在线软件SHEsis(http://analysis.bio-x.cn/)构建单倍型和双倍型。结果显示(表3),在试验沿河铁叫鸡群体中,存在4种单倍型(H1,H2,H3,H4),9种双倍型(H1H1、H1H2、H1H3、H1H4、H2H2、H2H4、H3H3、H3H4和H4H4)。优势单倍型H1(73),频率0.365,其次是H4(67),频率0.335;优势双倍型为H1H4(40),频率0.400,其次是 H1H3(12)和H3H4(12)频率均为0.120。
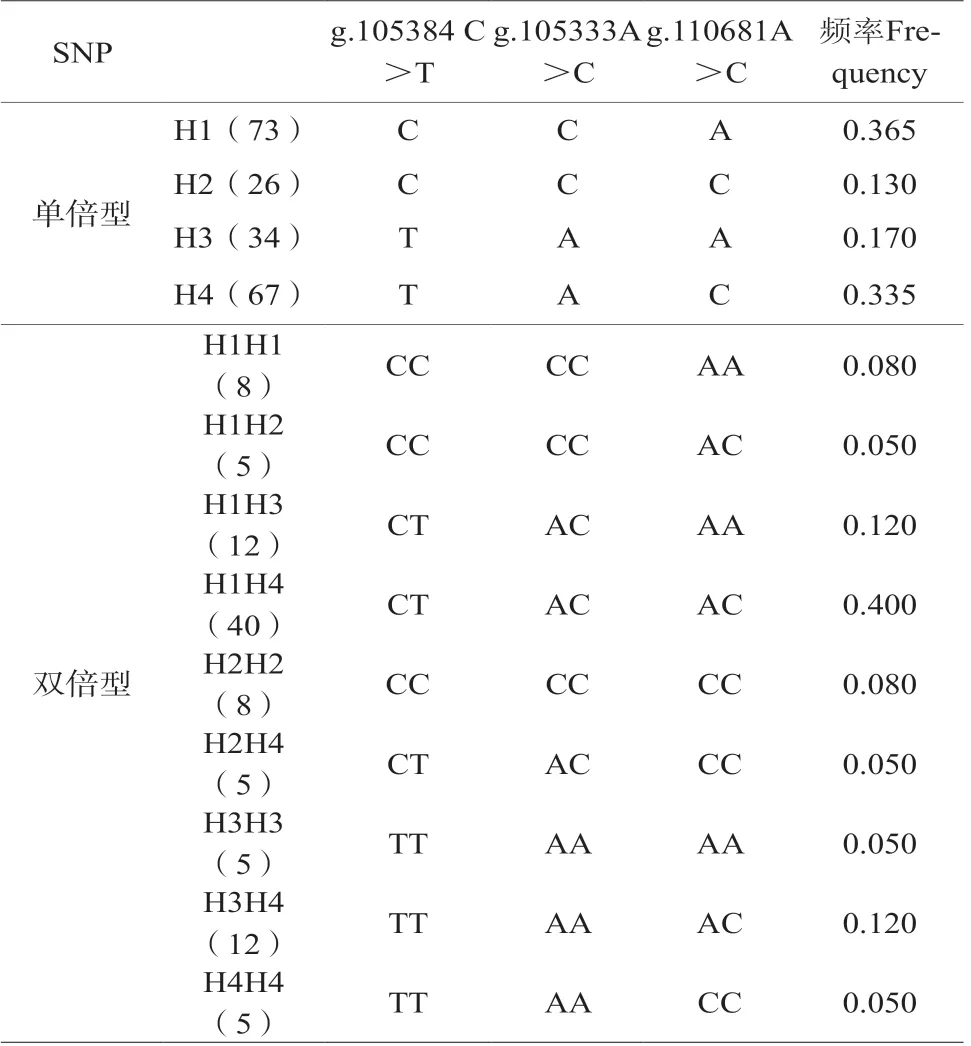
表3 SNPs的单倍型和双倍型分析
2.4 鸡IGF1R基因SNP与体尺的关联分析结果
一般线性模型(GLM)分析了IGF1R基因多态与体尺的关联性(表4)。结果显示,g.105384 C>T突变位点CC和CT基因型鸡的体斜长显著大于CC基因型(P<0.05);g.105333 A>C位点AA和AC基因型体斜长显著大于CC基因型(P<0.05),g.110681 A>C位点AA基因型髋骨宽显著大于AC和CC基因型(P<0.05)。
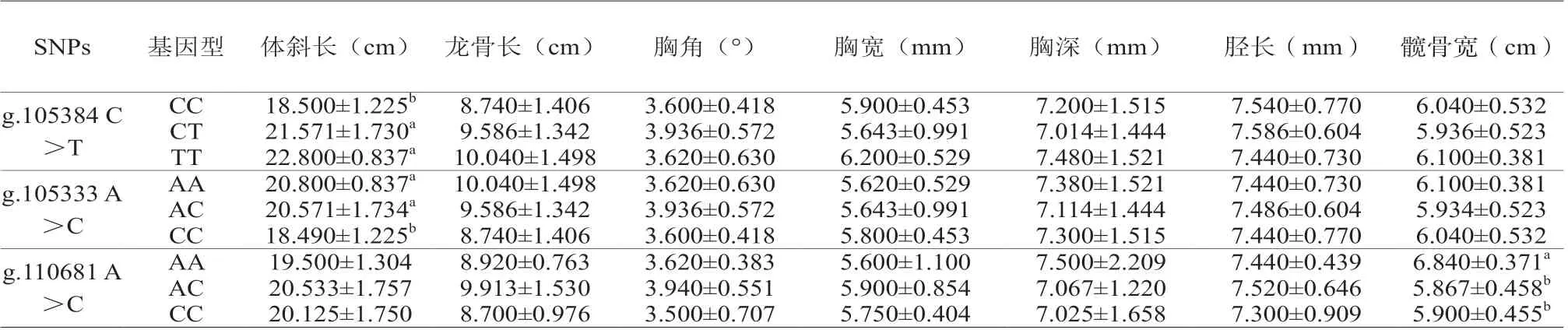
表4 IGF1R基因多态与体尺的关联性分析
2.5 鸡IGF1R基因SNP与屠宰性能的关联分析结果
试验分析IGF1R基因多态与体尺的关联性,详见表5。结果表面,g.105384 C>T位点TT基因型半净膛和全净膛重显著高于CC基因型(P<0.05),g.105333 A>C位点AA基因型半净膛和全净膛重显著高于CC基因型(P<0.05),g.110681 A>C突变位点CC基因型腿肌重显著高于AA和AC基因型。
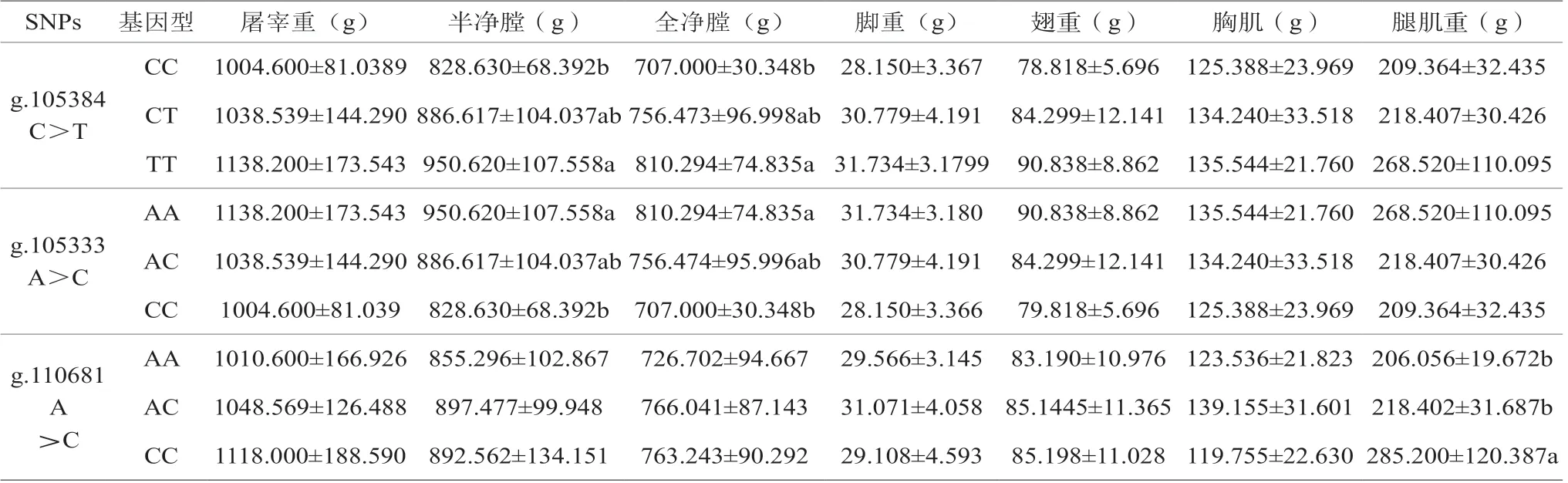
表5 IGF1R基因多态与屠宰性状的关联性分析
3 讨论
IGF1R基因已被研究为多种动物体尺和胴体性状的功能候选基因。在水牛、牦牛、山羊、猪和鸡身上发现了与许多这些特性的显著关联[9-12]。虽然在鸡方面,高凤华等[13]研究IGF1R基因多态性对生长性状的影响,但鸡的衍生品种中进行研究,同时IGF1R基因多态性与鸡屠宰性状的关联相对较少。Ekegbu[14]检测了新西兰罗姆尼绵羊IGF1R基因从9号外显子到10号内含子片段中核苷酸变异,并发现其影响羊的屠宰性状。而Byun等[15]发现了绵羊IGF1R基因核苷酸变异与寿命之间的关联。鉴于上述IGF1R基因的生物学功能,并且IGF1R基因与许多其他候选基因一样,对多个性状具有多效性作用[16]。在这种情况下,鸡的生长、胴体和肌肉质量的改善也可能伴随着肉质的改善。因此,广泛和复杂的研究,包括广泛的生产性状,有可能提供适当的价值的分子标记育种。
研究对IGF1R基因跨越部分内含子和全部外显子的片段进行多态性分析,3个SNP位点g.105384 C>T、g.105333 A>C和g.110681 A>C分别在IGF1R基因的外显子3被检测到,3个SNPs基因型分布符合Hardy-Weinberg平衡。某个群体未来世代的基因型和等位基因频率,当前种群的平衡状态常用Hardy-Weinberg平衡预测,当一个符合孟德尔群体之间交配随机,并且无选择、突变、迁移和遗传漂变等事件发生时,那么基因频率将保持不变,即这个群体被称为处于Hardy-Weinberg平衡[17]。根据关联分析法,3个SNPs可联合组成8种单倍型和36双倍型,但本试验只找到了4种单倍型和9种双倍型,与理论有差距,可能与试验群体小或其他单倍体个体在人工选育中被淘汰有关。
关联分析结果显示g.105384 C>T突变位点CC和CT基因型鸡的体斜长显著大于CC基因型(P<0.05);g.105333 A>C位点AA和AC基因型体斜长显著大于CC基因型(P<0.05),g.110681 A>C位点AA基因型髋骨宽显著大于AC和CC基因型(P<0.05)。g.105384 C>T位点TT基因型半净膛和全净膛重显著高于CC基因型(P<0.05),g.105333 A>C位点AA基因型半净膛和全净膛重显著高于CC基因型(P<0.05),g.110681 A>C突变位点CC基因型腿肌重显著高于AA和AC基因型。前人研究证实,同义突变SNP在IGF1R基因中的虽不直接改变蛋白的一级序列[18]。然而,它可以引起IGF1R蛋白表达、构象和功能的改变。而在羊中,3号外显子核苷酸c.654G > A突变,它编码了IGF1R的尿道亚基的一部分,子亚基包含1个富含半胱氨酸的区域,参与配体结合。因此,该区域的任何变异,例如由IGF1R基因特定片段的核苷酸替换引起的变异,可能会影响IGF1R与IGF1或胰岛素的相互作用,从而导致信号通路的改变。在绵羊IGF1R基因中,同义突变,c.654G > A突变可扰乱转录、剪接、共翻译折叠、mRNA稳定,从而导致其他功能相关的变化[19]。此外,同义替换可以影响蛋白质编码区域内的转录和剪接调节因子,从而调节基因表达。此外,最近研究揭示了非同义编码SNP和同义编码SNP在性状方面表现出相似的似然性和效应大小。g.105384 C>T、g.105333 A>C和g.110681 A>C位点一些基因型对鸡的体尺和屠宰性状有显著影响可能与此相关,不可忽视。在今后的选育培养过程中可根据实际需求选择培育。
猜你喜欢
分子催化(2022年1期)2022-11-02
作物学报(2022年12期)2022-10-14
中国农业科学(2022年16期)2022-09-19
渔业科学进展(2022年4期)2022-09-05
中国典型病例大全(2022年9期)2022-04-19
安徽农学通报(2022年6期)2022-04-07
电脑报(2020年40期)2020-11-06
新课程·下旬(2018年8期)2018-11-10
电脑知识与技术(2018年19期)2018-11-01
中学生理科应试(2016年4期)2016-11-19
