
齐穗后弱光胁迫对杂交籼稻节间非结构性碳水化合物积累转运的影响*
2022-10-15 05:16朱莜芸李秋萍何辰延袁玉洁黄小凡何宇欣任万军
中国生态农业学报(中英文) 2022年10期
周 星, 李 博, 朱莜芸, 李秋萍, 何辰延, 袁玉洁, 黄小凡, 何宇欣, 王 丽, 程 红, 任万军, 邓 飞
(四川农业大学农学院/西南作物基因资源发掘与利用国家重点实验室/农业农村部西南作物生理生态与耕作重点实验室 温江 611130)
水稻()产量与品质的形成取决于花后光合产物的积累以及花前营养器官储存物质的转运再利用, 其中茎鞘储存的非结构性碳水化合物(non-structural carbohydrates, NSC)是水稻籽粒灌浆的重要物质来源, 对籽粒的贡献率可达到40%左右。因此, 研究水稻茎鞘特别是节间中NSC的积累转运特性, 对于水稻丰产优质具有重要意义。
前人研究指出, 茎鞘NSC积累转运特性是品种和环境共同作用的结果, 逆境胁迫往往导致水稻NSC积累转运特性发生改变。高温胁迫下水稻茎鞘胼胝体大量积累, 胞间连丝受阻, 降低了茎鞘NSC的转运效率和转运贡献率, 导致营养器官积累较多的同化物。干旱胁迫加速水稻植株衰老, 叶片光合速率下降, 茎鞘NSC向籽粒转运增加, 籽粒灌浆时间缩短产量降低。低氮条件下水稻茎鞘淀粉合成与水解相关酶活性增强, 促进花后茎鞘NSC转运, 但因叶片同化物生产受到限制。此外, 不同节间NSC积累转运特性存在差异, 下部节间NSC转运量、转运率明显高于上部节间。
四川盆地属我国典型的弱光稻区, 具有“弱光、寡照”的生态特点。弱光胁迫下水稻叶片光合受阻, 进而扰乱了干物质的积累转运, 最终导致水稻产量大幅降低。前期研究发现, 弱光胁迫显著增加了抽穗后茎鞘NSC的转运量、转运率及转运贡献率,然而弱光胁迫对不同节间NSC积累转运特性的影响尚不明确。因此, 本试验以杂交籼稻品种‘宜香优2115’和‘C两优华占’为试验材料, 研究大田遮光对杂交籼稻不同节间NSC积累和转运特性以及产量的影响, 为四川盆地等弱光稻区耐阴品种筛选和栽培技术改良提供理论和实践依据。
1 材料与方法
1.1 试验地点与材料
试验于2020年在四川省成都市温江区公平镇四川农业大学惠和农场(30°43′N, 103°52′E)进行。水稻生长季节(3-9月)的降雨量为1040.6 mm, 平均气温为23.65 ℃。试验地土壤质地为中壤土, 0~20 cm土层pH为5.25, 有机质含量17.20 g·kg, 全氮含量1.83 g·kg, 全磷含量1.35 g·kg, 全钾含量3.66 g·kg,碱解氮含量187.60 mg·kg, 速效磷含量38.40 mg·kg,速效钾含量96.87 mg·kg。
供试品种为四川农业大学等单位选育的籼型三系杂交水稻‘宜香优2115’(宜香1A×雅恢2115)和北京金色农华种业科技股份有限公司选育的籼型两系杂交水稻‘C两优华占’(C815S×华占)。
1.2 试验设计
试验采用两因素裂区试验设计, 主区因素为光照处理, 设自然光强(CK)和遮光53%(SH)两个水平;副区因素为水稻品种, 设‘宜香优2115’和‘C两优华占’两个水平。共4个处理, 每处理重复3次, 小区面积为3.6 m×9.6 m。光强处理时间为齐穗期到成熟期, 在高约2.0 m的竹架上挂孔径为0.50 mm左右的白色棉纱布进行遮光处理, 纱布置于水稻冠层上方0.5 m左右, 以确保水稻群体良好的通透性, 避免引起温度、湿度等其他环境因素的变化。旱育壮秧, 4月17日播种, 30 d秧龄移栽(行穴距30 cm×24 cm),每穴栽3苗。施N180 kg·hm, 基蘖肥∶穗肥按6∶4施用, 其中基肥∶分蘖肥为7∶3, 促花肥∶保花肥为6∶4。按照N∶PO∶KO比例为2∶1∶2配施磷钾肥, 磷肥作基肥一次施用, 钾肥按底肥∶促花肥为5∶5比例施用。田间管理与病虫害防治等与当地高产栽培要求一致。
1.3 取样及测定方法
于齐穗期、齐穗后10 d、20 d和30 d和成熟期取样, 每小区取15根节间数相同、长势一致的单茎。其中, ‘宜香优2115’和‘C两优华占’茎蘖分别选取伸长节间数为6个和5个的单茎。各单茎去除叶片和叶鞘, 将着生稻穗的节间定义为第1节间, 其下的节间按从上到下的顺序依次为第2至6节间。按不同节间、稻穗分装, 105 ℃的烘箱杀青1 h后75 ℃烘干至恒重并称重。
参照Yoshida等方法, 采用蒽酮-硫酸法测定了水稻各节间可溶性糖和淀粉的含量, 并计算NSC积累转运相关指标:

成熟期每小区连续调查60穴计算有效穗数, 并按平均有效穗数从每小区取5穴, 考查每穗颖花数、结实率和千粒重等产量构成因素。各小区分别单打实收测产。
1.4 数据统计与方法分析
采用Microsoft Excel 2019处理数据, SPSS 27.0进行方差分析, Graphpad prism 8作图。
2 结果与分析
2.1 弱光胁迫对不同节间NSC含量的影响
由图1可知, 齐穗期下部节间(‘宜香优2115’第4~6节间, ‘C两优华占’第4~5节间) NSC含量明显高于上部节间(第1~3节间)。齐穗后除‘宜香优2115’第1、第2节间, ‘C两优华占’第2节间NSC含量呈明显的单峰趋势外, 各节间NSC含量均呈降低趋势,且齐穗后0~10 d节间NSC含量降低幅度最大, 而下部节间降低速率明显高于上部节间。除第1节间变化趋势不明显外, 齐穗后第2至第5节间NSC含量均表现为‘宜香优2115’>‘C两优华占’。对照‘宜香优2115’上部3节间NSC含量均呈先增加后降低趋势, 第4和6节间NSC含量则逐渐降低; ‘C两优华占’第2节间NSC含量在齐穗后20 d达峰值, 其余节间NSC含量在齐穗0~30 d阶段整体呈降低趋势, 在齐穗后30 d至成熟期各节间NSC含量则略有升高。除‘宜香优2115’第2节间齐穗后10 d NSC含量外,遮阴处理显著降低了齐穗后各节间NSC含量; 成熟期‘宜香优2115’第1~6节间NSC含量较对照显著降低20.98%~95.45%, ‘C两优华占’第2~5节间较对照处理则显著下降78.34%~86.13%。
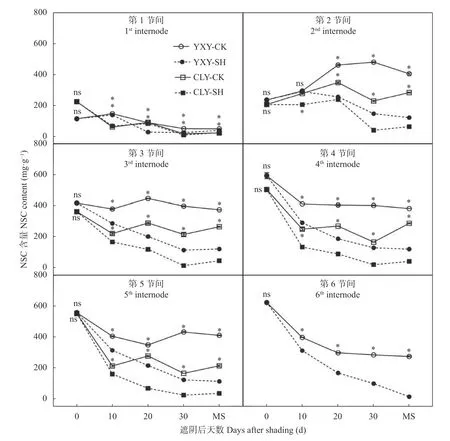
图1 弱光胁迫对水稻各节间非结构性碳水化合物(NSC)含量的影响Fig. 1 Effect of shading stress on the content of nonstructural carbohydrate (NSC) in each internode of rice
2.2 弱光胁迫对不同节间NSC积累量的影响
由图2可以看出, 齐穗期下部节间NSC积累量明显高于上部节间。‘宜香优2115’第4~6节间和‘C两优华占’第3~5节间NSC积累量逐渐降低, 且在齐穗后0~20 d降低幅度最大, 成熟期对照处理两品种第3~5节间NSC积累量略有回升。两品种第2节间及‘宜香优2115’对照处理第3节间NSC积累量均呈先增加后降低的趋势。各时期第2~5节间NSC积累量均表现为‘宜香优2115’>‘C两优华占’, 且‘C两优华占’第3~5节间NSC积累量在齐穗后0~10 d大幅降低, ‘宜香优2115’下部节间(第4~6节间)则在齐穗后0~20 d大幅降低。遮阴处理显著降低了两品种齐穗后各节间NSC积累量, 且下部节间NSC降低幅度明显高于上部节间。其中‘宜香优2115’第1~6节间降低36.60%~96.59%, 而‘C两优华占’第1~5节间下降17.39%~88.85%。
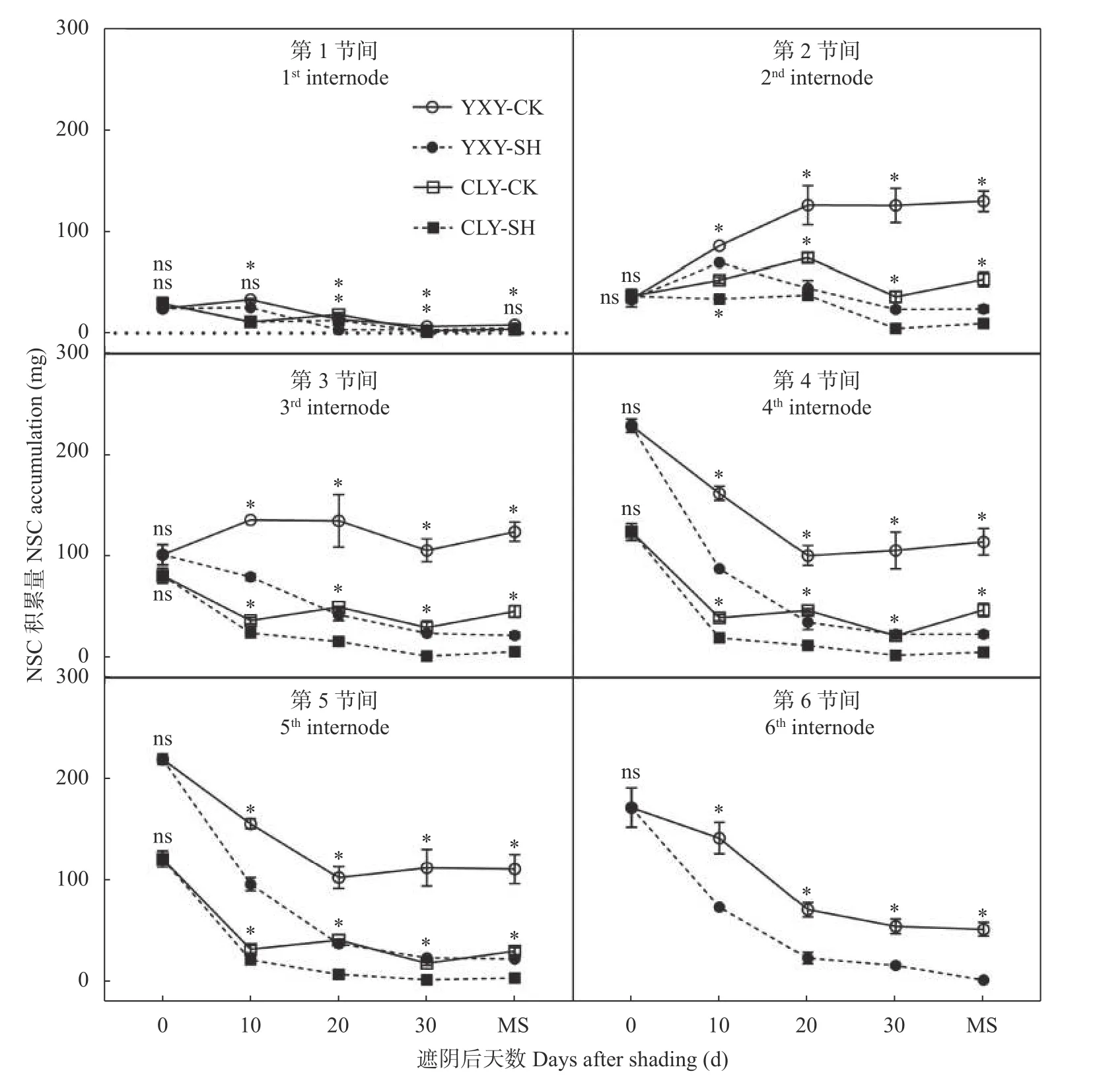
图2 弱光胁迫对水稻各节非结构性碳水化合物(NSC)积累量的影响Fig. 2 Effects of shading stress on the accumulation of nonstructural carbohydrates (NSC) in each internode of rice
2.3 弱光胁迫对不同节间NSC转运特性的影响
由表1可知, 品种、光照处理及二者互作极显著影响水稻第2~5节间NSC转运总量。‘宜香优2115’第4~6节间、‘C两优华占’第4~5节间NSC转运量明显高于第1~3节间。‘宜香优2115’上部节间转运量低于‘C两优华占’, 下部节间转运量大于‘C两优华占’, 进而导致其节间NSC转运总量显著高于‘C两优华占’。对照处理‘宜香优2115’第2、3节间和‘C两优华占’第2节间在齐穗后分别积累了95.86 mg、22.59 mg和16.44 mg, 遮阴处理使‘宜香优2115’第1~6节间NSC转运量显著增加19.03%~453.70%, ‘C两优华占’第1~5节间则增加3.37%~263.56%, 进而导致节间NSC转运总量分别显著增加182.86%和71.60%。此外, 遮阴处理‘宜香优2115’第2~5节间NSC转运量的增加量明显高于‘C两优华占’。
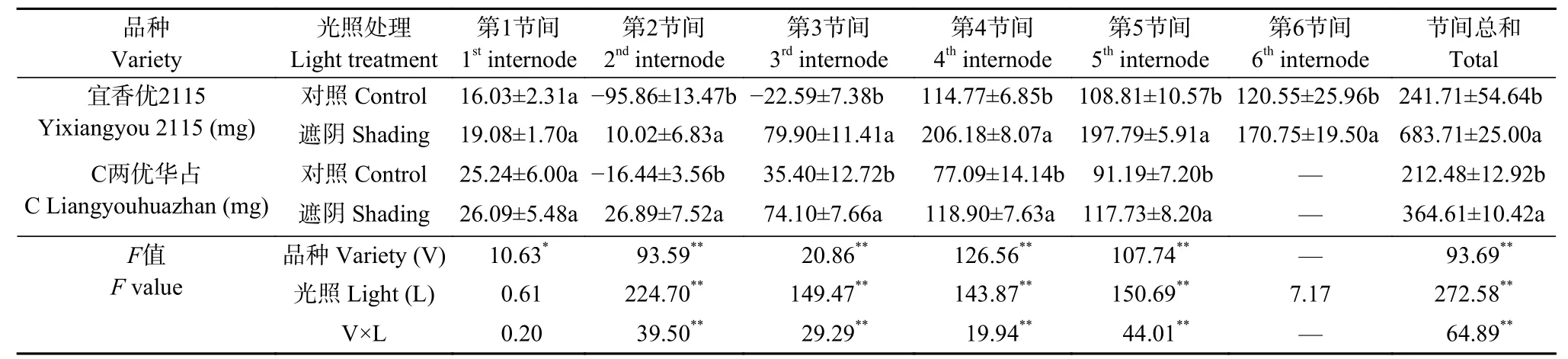
表1 弱光胁迫对水稻各节间非结构性碳水化合物转运量的影响Table 1 Effect of shading stress on the remobilization amount of stored nonstructural carbohydrates in each internode of rice
品种、光照处理主效应极显著影响水稻第1~6节间以及总节间NSC转运率, 二者互作则显著或极显著影响第1、2、3、5节间以及总节间NSC转运率(表2)。对照处理‘宜香优2115’第2、3节间和‘C两优华占’第2节间转运率为负值, 明显低于其他节间; 遮阴处理两品种第2节间NSC转运率明显低于其他节间。‘宜香优2115’第1~5节间转运率均明显低于‘C两优华占’, 进而导致其总节间NSC转运率下降。遮阴处理下‘宜香优2115’第1~6节间NSC转运率分别提高12.72~325.07个百分点, ‘C两优华占’第1~5节间转运率则增加3.25~118.47个百分点, 进而导致两品种节间NSC总转运率显著增加。遮阴处理下‘宜香优2115’第1~5节间NSC转运率较‘C两优华占’低5.88~44.59个百分点, 进而使其总节间NSC转运率降低5.51个百分点。
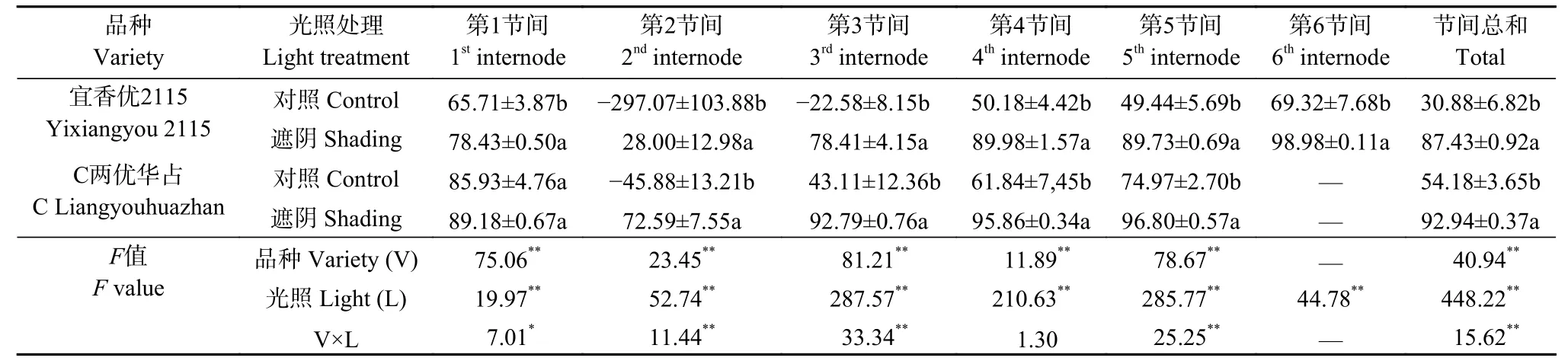
表2 弱光胁迫对水稻各节间非结构性碳水化合物转运率的影响Table 2 Effect of shading stress on the remobilization rate of stored nonstructural carbohydrates in each internode of rice %
表3表明, 品种主效应极显著影响第2、4、5节间以及节间NSC总贡献率, 光照主效应显著或极显著影响第1~6节间以及节间NSC总贡献率, 而光照与品种互作显著或极显著影响第2~5以及节间NSC总贡献率。对照处理‘宜香优2115’第2、3节间和 ‘C两优华占’第2节间NSC贡献率均为负值, 明显低于其他节间。遮阴处理两品种第1~2节间NSC贡献率明显低于其他节间。‘宜香优2115’上部节间贡献率低于‘C两优华占’, 但下部节间贡献率大于‘C两优华占’, 进而导致其节间NSC总贡献率显著高于‘C两优华占’。遮阴处理下‘宜香优2115’第1~6节间NSC贡献率分别提高0.63~9.18个百分点, ‘C两优华占’第1~5节间NSC贡献率则增加0.9~5.29个百分点, 进而导致两品种总节间NSC贡献率显著增加。遮阴处理‘宜香优2115’第3~5节间在NSC贡献率分别较‘C两优华占’提高0.26~5.99个百分点, 进而使其总节间NSC转运率提高20.49个百分点。
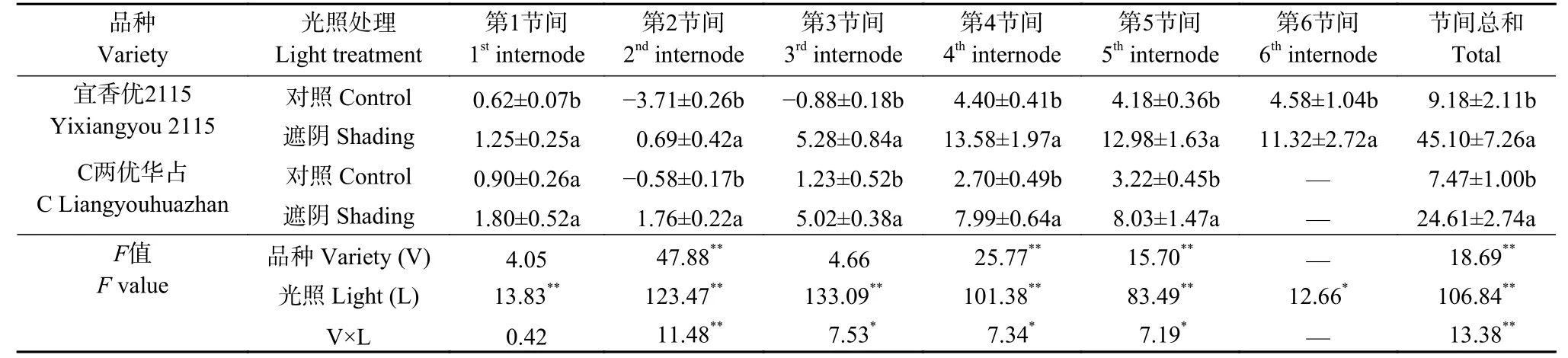
表3 弱光胁迫对水稻各节间非结构性碳水化合物转运贡献率的影响Table 3 Effect of shading stress on the contribution rate of remobilized nonstructural carbohydrates to panicle dry weight in each internode of rice %
2.4 弱光胁迫对水稻产量的影响
表4表明, 品种极显著影响水稻有效穗数、每穗颖花数、千粒重和产量, 而光照处理则显著或极显著影响每穗颖花数、结实率、千粒重和产量。‘C两优华占’有效穗数、每穗颖花数显著高于‘宜香优2115’, 导致其产量较‘宜香优2115’高27.76%~34.17%。遮阴显著降低两品种结实率, 以及‘C两优华占’的每穗颖花数, 加之两品种千粒重均呈降低趋势, 进而导致‘宜香优2115’和‘C两优华占’产量分别显著降低40.97%和43.79%。‘宜香优2115’结实率在遮阴处理下降低幅度更低, 进而导致产量下降幅度更小。
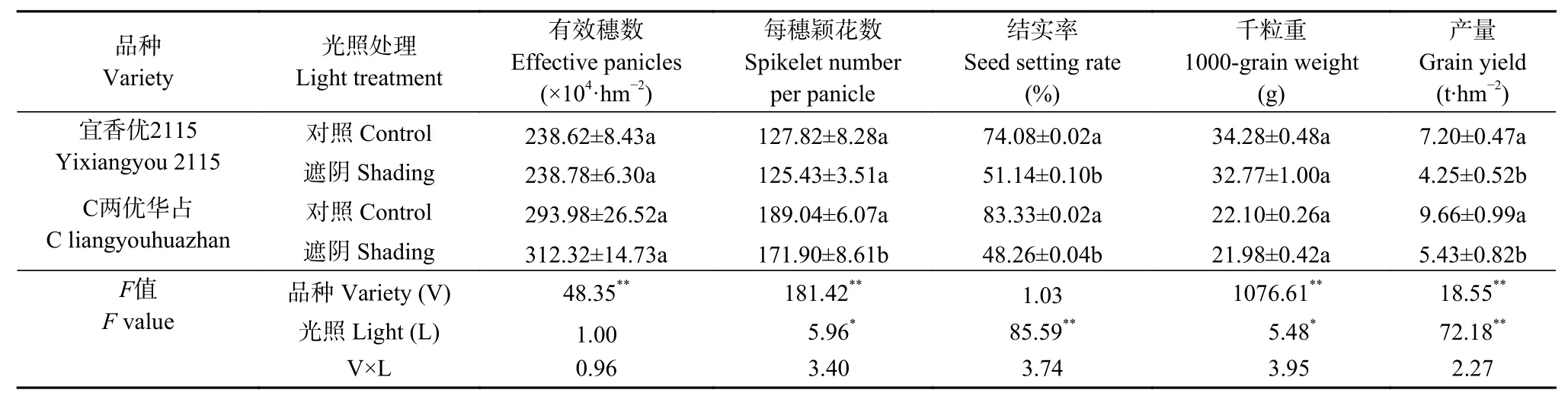
表4 弱光胁迫对水稻产量及构成因素的影响Table 4 Effect of shading on grain yield and yield components
2.5 NSC积累转运特性与产量的关系
以对照和遮阴处理间的产量差值, 与各节间对照和遮阴处理间的NSC含量、积累量、转运量和运转率的差值进行相关性分析(表5)。结果表明, 除齐穗后10 d第2节间NSC含量和积累量差值与产量差值呈正相关外, 齐穗后各节间NSC含量和积累量差值均与产量差值呈负相关关系。其中, 齐穗后20 d、30 d和成熟期第3节间和成熟期第5节间NSC积累量差值与产量差值相关性达到显著水平(<0.05)。此外, 产量差值还与第3和第5节间NSC转运量和转运率差值呈显著正相关关系(<0.05)。进一步以产量差值为因变量, 各节间NSC含量、积累量、运转率的差值为自变量进行多元回归分析, 得到回归方程=6.502+0.2229+0.061 53, 式中为第5节间NSC转运率差值,为成熟期第4节间NSC积累量差值, 回归方程决定系数为0.962, 且值达极显著水平(<0.01)。说明自变量与因变量之间具有显著相关性, 第5节间NSC转运率差值过大, 成熟期第4节间NSC积累量差值过高, 不利于缩小对照与遮阴处理间的产量差, 且第5节间NSC转运率差值对产量差值的影响远大于成熟期第4节间NSC积累量差值的影响。
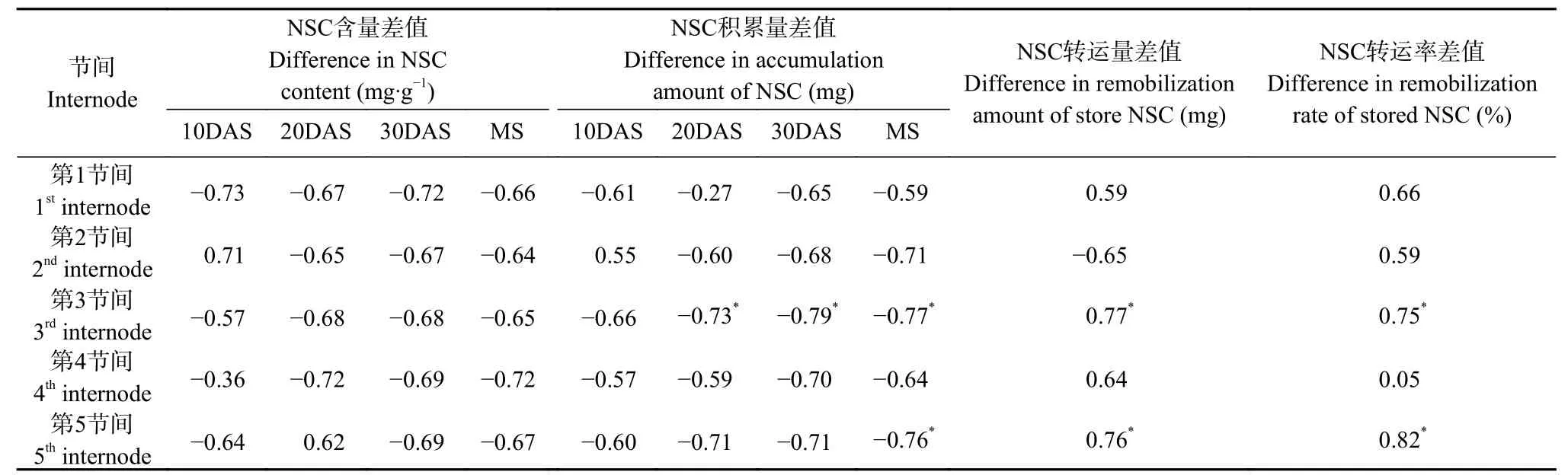
表5 水稻产量与节间非结构性碳水化合物(NSC)积累转运特性的相关性Table 5 Relationship between grain yield and the accumulation and remobilization characteristics of nonstructural carbohydrates(NSC) in internodes
3 讨论
3.1 水稻不同节间NSC积累转运特征
茎鞘储藏的NSC主要包括可溶性糖和淀粉, 是水稻光合作用的主要产物和呼吸作用的重要底物。NSC在植物生长发育和代谢中起着重要作用, 增加花前茎鞘NSC积累和花后NSC的转运有利于促进籽粒灌浆, 进而提高水稻产量。在开花期水稻茎鞘NSC积累量达峰值, 开花后茎鞘作为源器官, 其储藏的NSC向穗部进行转运, 用于籽粒灌浆充实。前人研究指出, 水稻茎鞘NSC积累转运是一个动态过程, 在籽粒灌浆盛期茎鞘NSC转运速率达最大值,而在灌浆后期, 由于光合同化产物供给超过了籽粒灌浆的需求, 茎鞘NSC重新积累。本研究结果表明, 齐穗后水稻节间中的NSC被转运到籽粒, 总节间NSC转运率为30.88%~92.94%, 其对每穗籽粒干重的贡献率为7.47%~45.10%, 与前人研究结果相似。有研究发现, 水稻、小麦()等作物不同节间的NSC积累转运特性存在差异。本试验研究发现, 第1节间(穗颈节)在NSC转运过程中主要起运输通道的作用, 其NSC含量、积累量及转运量均较低; 下部节间在齐穗期可积累较多的NSC,齐穗后节间中储存的NSC快速向籽粒转运, 以促进水稻结实灌浆; 上部节间NSC转运相对迟缓, 并且部分节间可出现NSC继续积累的现象, 其NSC转运量、转运率和贡献率均为负值。这主要是因为水稻节间的伸长是从下部节间到上部节间依次发生的, 这使得下部节间能够更早地作为库(源)器官发挥作用,而上部节间中的NSC向籽粒转运有延迟现象。而在光合产物供给充足的情况下, 部分节间主要作为库器官, 继续积累NSC。此外, 本研究同时发现,不同品种间NSC积累转运特性差异显著。较‘C两优华占’ ‘宜香优2115’在齐穗期具有更高的NSC含量和积累量, 而齐穗后0~10 d ‘C两优华占’第3~5节间则具有更高的NSC含量降低速率, 对照处理下其各节间NSC转运量、转运率均明显高于‘宜香优2115’。
3.2 弱光胁迫对节间NSC积累转运的影响
前人研究表明, 不同生育阶段弱光胁迫则导致水稻不同程度减产, 分蘖期遮阴会阻碍分蘖发生, 拔节孕穗期遮阴则主要影响稻穗和颖花的生长发育,开花灌浆期遮阴则使水稻授粉受精受限, 籽粒灌浆充实受阻, 进而导致结实率和千粒重均急剧下降。本研究结果表明, 齐穗后弱光胁迫显著降低结实率,加之每穗颖花数和千粒重均呈降低趋势, 进而使水稻产量显著降低。弱光胁迫下水稻叶片变薄, 叶绿素a、b含量增加, 叶绿素a/b值降低, 1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco)和Rubisco活化酶(RCA)的活性下降, 致使叶片净光合速率降低, 光合产物供应不足, 籽粒灌浆不良, 水稻产量大幅降低。水稻产量的形成主要由花后光合产物的直接供给和花前储藏NSC的转运再利用所决定。弱光胁迫下水稻可增强茎鞘中储存NSC的转运再利用, 以降低光合产物供给不足对产量的不利影响。本研究结果表明, 弱光胁迫显著降低了齐穗后水稻各节间的NSC含量和积累量, 进而使各节间NSC转运量增加3.37%~453.70%, 转运率提高3.25~325.07个百分点,最终使总节间NSC转运贡献率提高17.14~35.92个百分点, 从而降低了弱光胁迫对水稻产量形成的危害。对照与遮阴处理产量差值与成熟期第3和第5节间NSC积累量差值呈显著负相关, 与两节间NSC转运量和转运率差值则呈显著正相关关系。通过增强水稻各节间, 特别是第3和第5节间储存NSC的转运再利用, 可有效降低弱光胁迫对水稻产量的不利影响。本研究同时发现, 不同品种对弱光胁迫的响应存在明显差异。与‘C两优华占’相比, ‘宜香优2115’各节间在齐穗期具有更高的NSC含量和积累量, 遮阴处理各节间NSC转运量的增加量, 以及转运率和贡献率的增幅均明显高于‘C两优华占’, 最终使其结实率和产量降低幅度更小。可见, 筛选花前节间NSC积累量高, 花后转运量大的品种有利于降低弱光胁迫对水稻的不利影响。此外, 也可通过适当的栽培措施来调节茎鞘NSC的积累转运, 从而确保弱光稻区水稻丰产。
4 结论
水稻不同节间非结构性碳水化合物积累转运特性存在明显差异, 下部节间储藏的非结构性碳水化合物在齐穗后即被迅速转运利用, 而上部节间非结构性碳水化合物转运相对迟缓, 且部分节间可出现非结构性碳水化合物继续积累现象。弱光胁迫显著降低了水稻的产量, 而通过增强水稻各节间(特别是第3和第5节间)非结构性碳水化合物的转运再利用, 可一定程度弥补弱光胁迫下光合产物供给不足对水稻产量的影响。‘宜香优2115’在具有较高齐穗期节间非结构性碳水化合物积累量的同时, 拥有更大的花后节间非结构性碳水化合物转运量, 从而降低了弱光胁迫对结实率和产量的影响。因此, 生产上筛选花前非结构性碳水化合物积累量高, 花后转运量大的品种是弱光稻区水稻丰产的有效途径。
猜你喜欢
高中时代(2017年7期)2018-02-24
南方农业·下旬(2017年8期)2017-10-23
领导文萃(2017年11期)2017-06-12
山东农业科学(2016年12期)2017-01-21
高教探索(2015年10期)2015-10-29
山东农业科学(2014年7期)2014-09-22
摄影世界(2014年7期)2014-09-10
科技与创新(2014年8期)2014-07-17
