
生物炭对盐渍化土壤周年温室气体排放及冻融效应的影响*
2022-10-15 05:16张如鑫屈忠义刘祖汀王麒源王丽萍
中国生态农业学报(中英文) 2022年10期
张如鑫, 屈忠义,2, 杨 威, 刘祖汀, 王麒源, 王丽萍**
(1.内蒙古农业大学水利与土木建筑工程学院 呼和浩特 010018; 2.内蒙古农业大学寒旱区灌溉排水研究所 呼和浩特 010018)
大气中温室气体浓度的升高目前被认为是导致全球气候变暖的主要原因之一, 内蒙古作为我国的粮食主产区, 1961年以来内蒙古10年的平均增温达到0.36 ℃, 明显高于同期全球(0.12 ℃)和全国(0.23℃)的增温幅度, 是我国气候变暖最显著的地区之一。一方面, 河套灌区属于我国干旱、半干旱地区, 土壤盐渍化程度高、分布面积广。盐渍化土壤通过影响微生物活性进而影响土壤温室气体排放和吸收过程,此过程在干旱、盐渍化地区长期普遍存在。另一方面, 冻融对土壤盐渍化的影响十分显著, 是土壤盐碱化独特的形成机制, 且冻融所形成的盐渍化土壤会对作物生长发育产生巨大的威胁, 其中灌区土壤冻融期伴随着初冻期、冻结期和融解期等多个过程,其变化直接关系土壤理化性质和微生物活性, 而这些因素与土壤温室气体的排放密切相关, 据报道冻融期土壤温室气体排放量在全年总排放量占有较大的比重。因此, 研究河套灌区盐渍化土壤周年温室气体排放规律及冻融效应问题对于改善土壤性质、提高作物生产力、减缓环境压力具有重要意义。
生物炭作为一种新型环境功能材料, 在自然条件下较为稳定, 具有改善土壤理化性质、固碳减排等作用, 被广泛应用于农田土壤。成功等针对小麦()-玉米()轮作系统发现,施用16 t·hm的小麦秸秆生物炭降低了土壤氧化亚氮(NO)、二氧化碳(CO)年排放总量和甲烷(CH)的年吸收总量, 分别为38.7%、14.4%和50.1%。Barracosa等通过盆栽试验也发现生物炭与矿物相结合可使NO排放量降低36%, 全球增温潜势(global warming potential, GWP)值降低26%。但也有一些报道认为, 生物炭在温室气体减排方面具有一定的副作用, 原因可能是土壤类型、生物炭施加量、生物炭种类等其他环境因素不同而存在差异。Uchida等以野豌豆()为研究对象, 发现施加稻壳生物炭并没有减少NO累积排放量, 反而提前了土壤NO排放峰值。魏甲彬等研究冬季稻田也发现,施入水稻壳生物炭会显著增加潮泥土壤CO排放,但在水稻()生育期的提高效果不显著。随着冻融交替过程中农田土壤温室气体排放量的明显增加, 冻融期温室气体排放问题逐渐受到重视。刘翔等研究发现, 生物炭的添加会增加冻融过程中的土壤CO排放量, 但同时促进了土壤CH的吸收和NO的减排。Gao等研究同样发现, 在冻融作用下, 生物炭能够通过影响土壤水热条件促进土壤CO排放。以上研究大多通过短期试验研究生物炭对土壤温室气体排放的影响, 但有研究表明, 生物炭对温室气体排放的影响随施用时间会发生改变, 生物炭在生物和微生物的长期影响下会被老化, 其理化性质会发生改变。Spokas发现, 3年陈化后的生物炭较新施加的生物炭的CH氧化能力更高, 但其抑制土壤NO排放的能力有所下降。而吴震等研究水稻小麦轮作系统却发现, 陈化后的生物炭比新生物炭更能显著降低土壤综合温室效应和温室气体强度。可见, 不同生物炭在不同时期对不同土壤类型的温室效应具有不同的影响。
目前在生物炭对土壤温室气体排放影响方面,多数学者针对的是草地、酸性农田、湿地以及水田等非盐渍化土壤的生态系统, 且主要集中在作物生长期、播种期、冻融期以及室内试验等不连续的试验周期, 针对施入生物炭对盐渍化土壤周年温室气体排放的影响研究鲜有报道。鉴于此, 本文在内蒙古自治区巴彦淖尔市五原县“五万亩盐渍土壤改良示范区”进行小区田间试验, 探究施加生物炭对盐渍化土壤周年温室气体排放的影响, 为改善河套灌区盐渍化土壤性质、剖析周年温室气体排放机制、提出适宜的减排措施提供科学依据和技术支撑。
1 材料与方法
1.1 研究区概况和试验材料
试验于2019年11月-2021年4月在内蒙古巴彦淖尔市五原县“五万亩盐渍土壤改良示范区”(40°46′30″~41°16′45″N, 107°35′70″~108°37′50″E)进行。该地位于内蒙古河套平原腹地, 属于典型的温带大陆性气候, 气候干燥, 土壤封冻期长, 且冬春季土壤盐分表聚现象严重。多年平均气温6.1 ℃, 极端最低气温-36.7 ℃, 年均降雨量177 mm, 无霜期117~136 d。年均辐射量6418 MJ·m, 全年日照时数3231 h。冻融期日照时数占全年的44%。平均最低地温-12.3 ℃(1月), 冻土深度为100~130 cm, 土壤11月初进入初冻期, 冻层全部融化在翌年4月中旬左右。
试验开始前选取5个1 m×1 m样方, 去除表层植物碎屑等杂物, 使用直径为50 mm螺旋钻采集0~20 cm的土样。样品均匀混合后装进密封袋, 24 h内带回实验室, 经自然风干, 过2 mm筛备用。土壤质地为粉质壤土, pH为8.87, 电导率值为1.88 mS·m, 含盐量为3.59 g·kg, 阳离子交换量(CEC)为7.56 cmol·kg,有 机 质12.33 g·kg, 速 效 磷7.53 mg·kg, 速 效 钾351.50 mg·kg, 碱解氮58.91 mg·kg, 砂粒16.70%, 粉粒79.03%, 黏粒4.27%。供试生物炭为辽宁金和福农业开发有限公司的玉米秸秆生物炭产品, 该生物炭在360 ℃厌氧条件下热解处理制备而成, pH为8.75, 速 效 磷307.52 mg·kg, 速 效 钾786.50 mg·kg,碱解氮401.94 mg·kg, 碳含量364.07 g·kg, 氮含量7.56 g·kg, 磷含量0.17 g·kg, 碳氮比81.57。
1.2 试验设计
根据课题组多年探究生物炭对田间土壤理化性质、温室气体排放、作物产量的影响, 综合得出生物炭施用量为15 t·hm和30 t·hm对土壤固碳减排、保水保肥、作物增产等方面效果最佳。因此本试 验 设 置3种 生 物 炭 水 平: 0 t·hm(CK)、15 t·hm(B15)和30 t·hm(B30), 每个处理设置3个重复, 共9个小区, 完全随机区组排列。小区四周设置2 m宽的保护行。试验小区面积为160 m(40 m×4 m), 2019年10月15日将生物炭均匀撒施在试验小区上, 使用旋耕机将其与20 cm土层土壤均匀混合, 并在每个小区安装取气基座和土壤温度传感器, 取气基座安放在行与行的中间位置, 并保证每次取样前基座内除土壤外无积雪、植株等其他物质, 之后不再施加生物炭。
试验田2020年4月25日进行春季灌水压盐, 灌溉定额为225 m·hm, 灌水方式为地面灌溉。播前进行人工耕翻, 深度约30 cm, 并施入磷酸二胺(N质量分数18%, P质量分数46%) 450 kg·hm和复合肥(N质量分数15%, P质量分数15%, K质量分数15%)337.5 kg·hm作为底肥。供试作物为当地农户普遍种植的‘902’葵花品种, 采用覆膜种植, 人工点播, 行距60 cm, 株距50 cm。病虫草害化学防治以及其他田间管理措施同当地作物高产管理一致。
将试验研究期划分3个阶段: 第1年冻融期(1 a冻融期, stage Ⅰ)、作物生长期(stage Ⅱ)和第2年冻融期(2 a冻融期, stage Ⅲ)。为更准确分析冻融期各个阶段土壤温室气体排放量的差异, 参考往年气象数据, 特将冻融期具体划分3个阶段: 初冻期、冻结期和融解期。日最低温开始达到0 ℃以下, 土壤表层开始冻结至日最高温达到0 ℃以下阶段为初冻期;当日最高温均持续低于0 ℃, 冻结锋面快速向下移动至最大冻结深度, 且积雪不再消融阶段为冻结期; 当春季日最高温达到0 ℃以上积雪开始融化, 土层开始双向解冻至全部解冻阶段为融解期。具体试验阶段划分如图1所示。
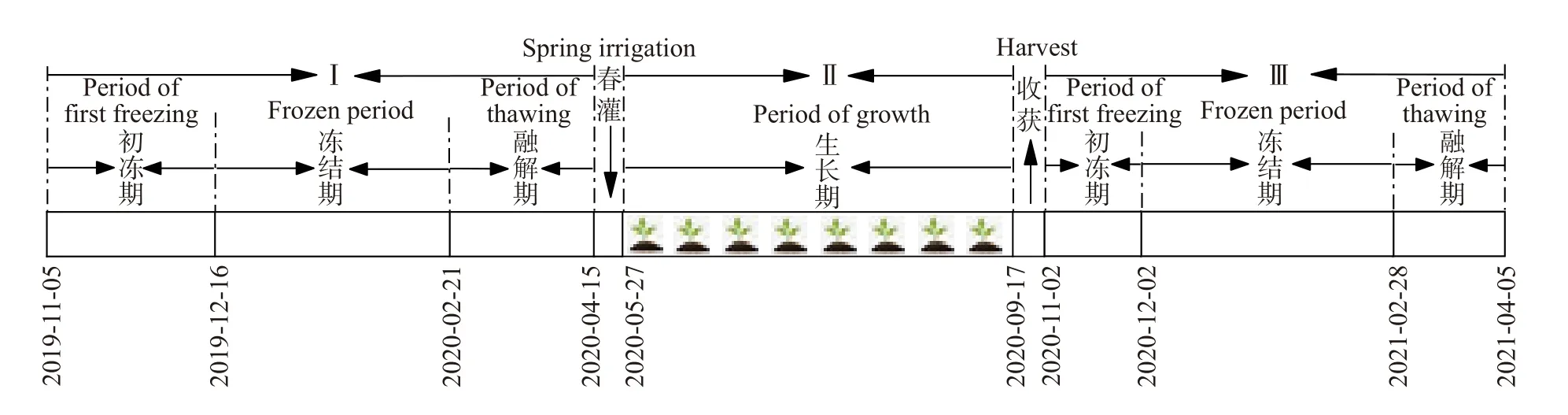
图1 试验期划分的阶段及其对应日期(年-月-日)Fig. 1 Stages of the trial period and their corresponding dates (year-month-day)
1.3 测定指标与方法
采用静态暗箱-气相色谱法进行温室气体(CH、CO和NO)定位观测。采集时间为上午9:00-12:00。采集频率设定为: 冻结期由于气候寒冷取样艰难, 故约每20 d取一次; 初冻期和融解期由于气候昼夜温差较大, 土壤处于冻融状态, 土壤温室气体排放波动较大, 因此采集气体频率设定约为每10 d一次; 作物生长期则按照葵花的生长阶段取样(苗期、现蕾期、开花期、灌浆期、成熟期)。静态箱由不锈钢制成,厚度为1 mm, 包括顶箱和基座两部分, 顶箱规格为40 cm×40 cm×40 cm, 底座的规格为40 cm×40 cm×15 cm。采样箱内装有小型风扇, 采样时通电持续转动, 以充分混匀箱内气体。在试验初期安放地箱,每个试验小区3个地箱(3个重复),并保证地箱内的土壤不受扰动。气体样品采用Picarro G2308分析仪(美国Picarro公司)测定CO、CH、NO含量。
箱体内所测样品的浓度()计算公式为:

式中:为标气浓度,为样品峰面积,为标气峰面积。
温室气体排放通量()计算公式为:
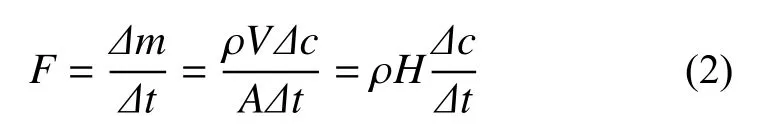
式中:为时间内箱内气体质量和混合比浓度的变化,为箱内气体密度,为箱底面积、体积、气室高度。
温室气体累积排放量()计算式为:

式中:为测量次数;F、F为第、+1次采样时温室气体的排 放通量, mg·m·h;t、t为第+1次采样的时间间隔, d。
综合增温潜势(GWP)是将各种温室气体的增温潜势换算为CO当量。100 a时间尺度的综合增温效应计算式为:
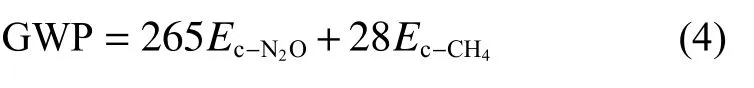
土壤样品采用土芯法采集, 使用手持式动力取样器采集深度0~20 cm土层土壤, 每10 cm为一层,每个处理重复3次。土样采集时间及频率与取气一致。冻融期有积雪覆盖时, 选取积雪厚度一致区域取样, 取样前用铲子将表层积雪清理干净, 取样结束后将积雪再次覆盖。取出原状土后立即将土壤样本放入铝盒内, 使用烘干法测定土壤质量含水率、采用pH计测定(土水比, 1∶2.5)土壤pH、采用电导率仪测定(土水比为1∶5)土壤电导率, 采用紫外可见分光光度计以及火焰光度计测定土壤速效磷和速效钾含量。采用土壤温度传感器(TM-03, 邯郸市若腾电子科技有限公司)对0~20 cm土层土壤温度进行监测, 在每个小区中间各布设一个土壤温度传感器,监测时间间隔设定为每4 h一次, 试验期末读取数据。
1.4 统计分析
数据采用Microsoft Excel 2010和SPSS 25.0 [运用LSD法进行差异显著性检验(<0.05)]进行整理分析, 使用Origin 9.1软件进行作图。
2 结果与分析
2.1 试验期温度、降水、积雪厚度和土壤水热变化
如图2所示, 2019-2021年试验区日最高、最低气温分别出现在7月和1月, 总体趋势为冻融期先降低后上升, 作物生长期先上升后下降, 整体呈现 “谷-峰-谷-峰”4个层次。试验期间累计降雨125 mm,主要集中在作物生长期(7、8月), 其中2020年8月11日出现一次强降雨, 降雨量达35.0 mm。通过测定试验期积雪厚度发现, 降雪主要集中在1月初, 其中1 a冻融期的降雪幅度比2 a冻融期明显增多。试验期土壤含水率受降雨量和积雪融化水的影响。10 cm和20 cm土层土壤含水率在1 a冻融期分别为16.74%和19.21%, 在作物生长期分别为22.35%和23.13%,在2 a冻融期分别为15.41%和17.54%。总体来看,3个时期的20 cm土层土壤含水率均高于10 cm土层土壤含水率。各土层土壤温度变化趋势与气温大致相同, 且初冻期和冻结期20 cm土壤温度高于10 cm土壤温度, 而融解期和生长期则相反。
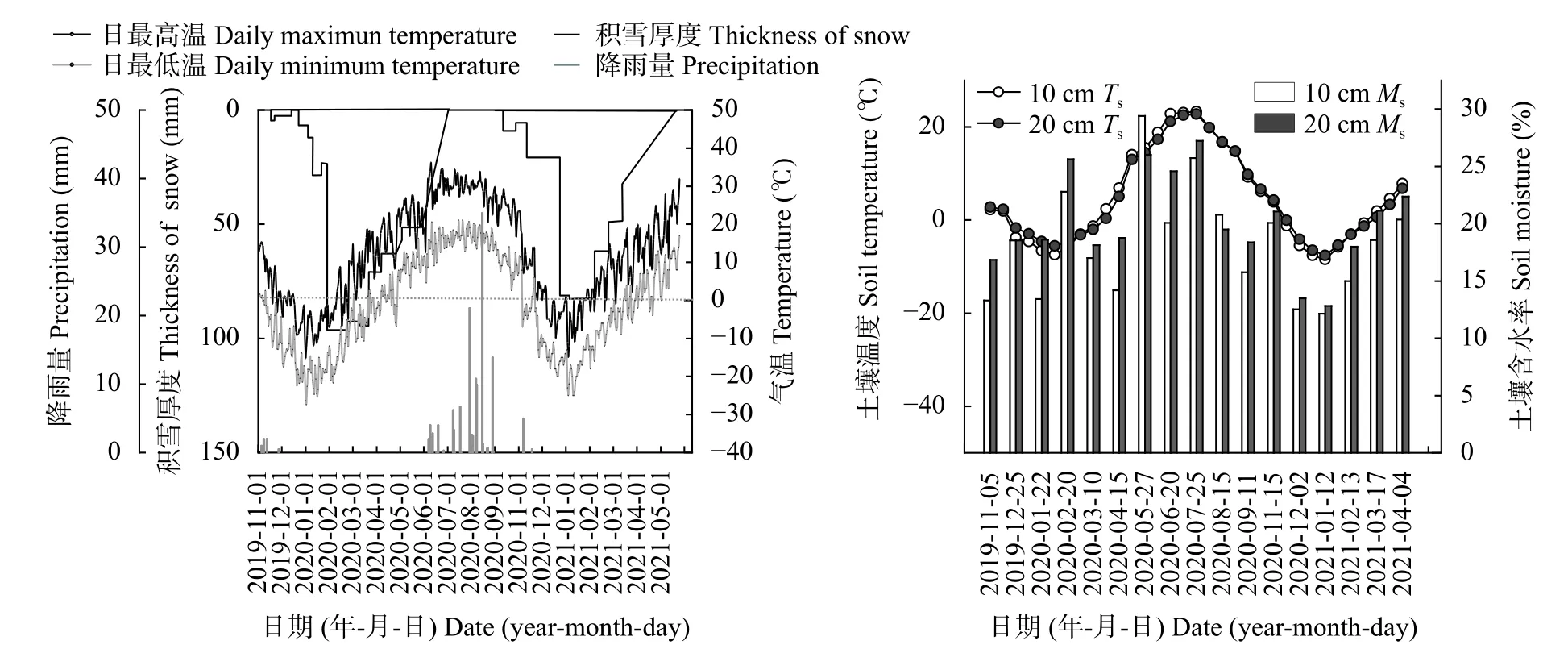
图2 试验期研究区气温、降水、积雪厚度和表层土壤温度(Ts)和含水率(Ms)的变化Fig. 2 Changes of air temperature, precipitation and thickness of snow, and surface soil temperature (Ts) and moisture (Ms) during the experimental period in the study area
2.2 土壤理化性质
与CK相比, 生物炭处理均提高了土壤pH、电导率、含水率、温度和速效磷、钾含量(表1), 此影响效果经过两年土壤冻融作用后仍然存在。生物炭显著提高了1 a冻融期、作物生长期和2 a冻融期的土壤电导率(<0.05), 其中, 与CK相比, B15处理使1 a冻融期、作物生长期和2 a冻融期的土壤电导率分别增加26.92%、31.71%和80.57%, B30处理分别增加39.23%、25.85%和63.51%。B30处理较CK显著增加了3个时期的土壤速效磷和速效钾含量(<0.05), 速效磷增幅为26.67%~36.77%, 速效钾增幅为12.17%~25.05%。分析各时期土壤含水率和温度发现, 生物炭处理仅对2 a冻融期的土壤含水率具有显著提高的效果, 对土壤温度也仅在1 a冻融期和2 a冻融期有显著影响, 可以看出生物炭的施入对各时期土壤水热状况的影响效果具有一定的差异性。整个试验期土壤pH在8.05~8.56间波动, 与CK相比, 生物炭处理均提高了土壤pH, 但效果不显著。可见无论是在冻融期还是作物生长期, 生物炭的添加均改变了土壤各理化指标, 但效果显著与否可能受环境以及生物炭老化等众多因素的影响。
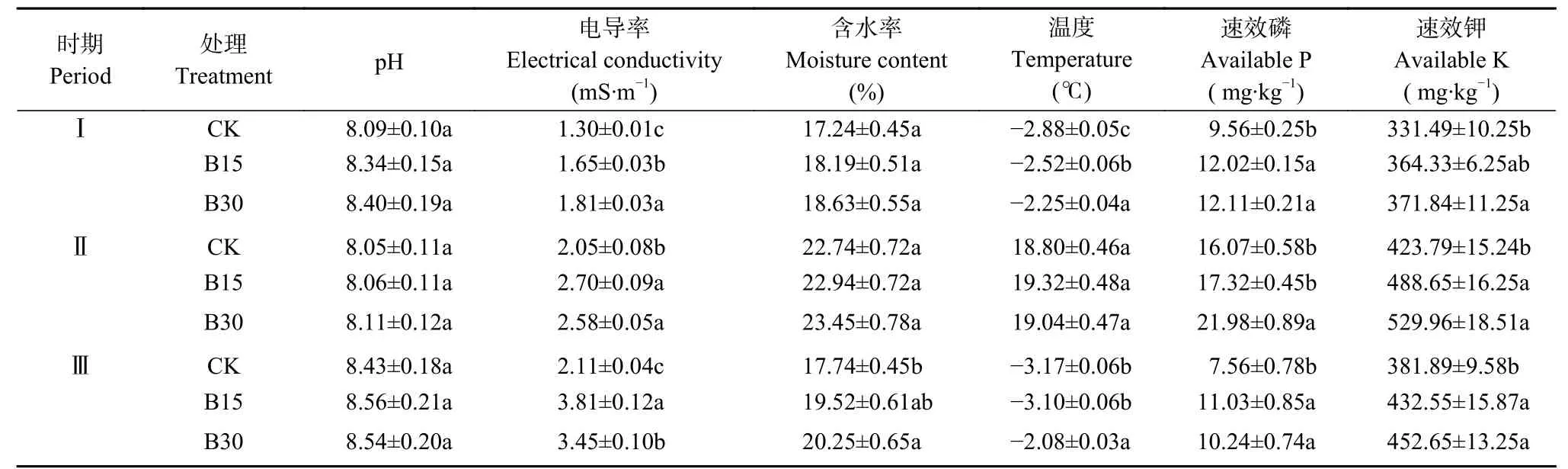
表1 不同生物炭用量下不同时期0-20 cm土壤理化性质Table 1 Physical and chemical properties of 0–20 cm soil in different periods under different biochar rates
2.3 施加生物炭对土壤温室气体排放的影响
2.3.1 CO排放通量
如图3所示, 各处理不同采样时期土壤CO通量具有明显的季节变化规律, 冻融期较低, 作物生长期较高, 整体与温度变化规律相似, 呈现“谷-峰-谷-峰”势态。初冻期(2019年11月14日-12月16日和2020年11月2日-12月2日)和 冻 结期(2019年12月16日-2020年 月2月22日 和2020年12月2日-2021年3月1日)温度低, CO通量由于冻结作用而固定于冻层中, 整体处于较低水平; 随温度上升,融 解 期(2020年2月22日-4月15日 和2021年3月1日-4月1日)和作物生长期(2020年5月27日-9月17日) CO被释放出来, 土壤CO通量处于排放高峰期。施加生物炭后土壤CO平均排放通量均低于CK。作物生长期各处理间差异显著(<0.05),B15和B30较CK分 别 降 低9.71%和14.88%; 1 a冻融期CK、B15、B30的CO排放通量分别为71.63 mg·m·h、69.21 mg·m·h和65.30 mg·m·h, 2 a冻 融 期 分 别 为44.02 mg·m·h、40.56 mg·m·h和40.62 mg·m·h, 与CK相比, 生物炭处理降低幅度分别为3.38%~8.84%和7.71%~7.85%, 但处理间无显著性差异。通过对比两年冻融期土壤CO平均通量发现, 2 a冻融期各处理CO平均通量较1 a冻融期降低39.26%, 可能是因为第1年冬季降雪较多, 积雪覆盖使土壤保温效果增强, 从而使土壤CO排放升高。
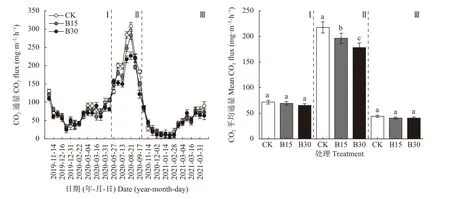
图3 不同生物炭用量下试验期土壤CO2排放通量差异Fig. 3 Differences of soil CO2 emission fluxes during the experimental period under different biochar rates
2.3.2 CH排放通量
炮弹鬼叫着飞过来时,五连长底柱眼皮都没抬一下,从一九三七年淞沪保卫战一直打到一九四二年衢州会战,侗族汉子底柱成了团里出名的老兵油子。打了六年仗,底柱身边的兄弟死了一茬又一茬,一起打淞沪的现在一个也不剩,说也怪,别人死的死,伤的伤,逃的逃,走的走,偏偏他底柱六年毫发无损,眼瞅着鬼子的子弹都打着他了,却不是被钢盔挡住就是被手榴弹挡住,就说手里这支汉阳造吧,188团一八八团上上下下谁人不识?见到枪就知道底柱来了:一发零点五八弹头大半根扎在枪托上,只露出半截屁股,屁股已被底柱大手磨得寒光闪闪。
如图4所示, 在融解期和作物生长期个别阶段均出现了土壤CH排放峰, 这可能是因为积雪融化、土壤解冻以及夏季降雨使土壤处于湿润状态, 形成了产生CH所需要的厌氧环境, 促进了土壤CH排放。其余时期各处理土壤均以吸收CH为主。处理CK、B15、B30土壤CH平均通量在1 a冻融期分别为-1.92 μg·m·h、-3.09 μg·m·h、-3.46 μg·m·h,作物长期分别为-1.47 μg·m·h、-1.92 μg·m·h、-2.13 μg·m·h, 2a冻融期分别为-3.20 μg·m·h、-3.47 μg·m·h、-4.58 μg·m·h。与CK相 比, 3个阶段生物炭处理降低土壤CH排放通量幅度分别为61.03%~80.58%、30.13%~44.89%、8.66%~43.15%。其中, 各处理在1 a冻融期和作物生长期差异达显著性水平。分析两年冻融期土壤CH平均通量发现, 在1 a冻融期, 生物炭处理与CK处理间均达显著性差异,而2 a冻融期仅B30处理与CK差异达显著水平, 可以看出生物炭处理抑制CH排放的能力有所减弱,且2 a冻融期各处理吸收CH强度大于1 a冻融期。

图4 不同生物炭用量下试验期土壤CH4排放通量差异Fig. 4 Difference of Soil CH4 emission flux during the experimental period under different biochar rates
如图5所示, NO排放主要集中在融解期(2020年2月22日-4月15日 和2021年3月1日-4月1日)和作物生长期(2020年5月27日-9月17日)。通常在土壤水分充足时土壤NO排放达到峰值, 最大峰值出现在春灌后-播种前期(2020年5月27日), 其中CK处理的NO排放峰值超过80 μg·m·h,其余峰值出现在2020年3月16日和2021年3月4日(大面积积雪融化后)以及2020年8月21日(强降雨后)。此外, 初冻期(2019年11月14日-12月16日和2020年11月2日-12月2日)和冻结期(2019年12月16日-2020年月2月22日和2020年12月2日-2021年3月1日)由于土壤水分和温度均较低, 土壤NO排放量较小, 1 a、2 a冻融期NO排放量分别在-7.87~5.66 μg·m·h和-7.84~7.53 μg·m·h间波动。分析各时期土壤NO平均通量发现, 施加生物炭抑制了土壤NO排放, 与CK相比, 1 a冻融期B15和B30分别降低23.14%和63.69%, 作物生长期分别降低19.29%和31.23%, 2 a冻融期分别降低41.27%和88.78%, 各处理间均达到显著性差异。分析两年冻融期土壤NO平均通量发现, 2 a冻融期土壤NO排放量大于1 a冻融期, 说明土壤冻融作用对NO排放影响较小。

图5 不同生物炭用量下试验期土壤N2O排放通量差异Fig. 5 Difference of Soil N2O emission flux during the experimental period under different biochar rates
2.4 生物炭对GWP及葵花产量的影响
如表2所示, 生物炭的施入对两年冻融期土壤CO累积排放量无显著影响; 作物生长期各处理间达显著性差异, 与CK相比, B15和B30分别降低9.86%和14.37%。各时期CH累积排放量均为负值,说明冻融期和作物生长期土壤均以吸收CH为主,与对照相比, 生物炭处理在1 a冻融期和作物生长期抑制CH累积排放量均达显著性差异, 而在2 a冻融期仅B30处理达显著性差异, 说明生物炭具有促进土壤吸收大气CH的作用。整个试验期, 生物炭处理抑制土壤NO排放效果显著, 其中3个时期的NO累积排放量大小均为CK>B15>B30。通过计算100 a尺度下增温潜势发现, 生物炭可显著降低冻融期和作物生长期综合增温潜势。1 a、2 a冻融期的GWP值均为负值, 说明在冻融期各处理土壤不具有增温效应, 且生物炭处理GWP值均显著低于CK, 增温潜势大小顺序为CK>B15>B30; 作物生长期, B15和B30的GWP值较CK分别降低15.74%和30.19%。就葵花产量而言, 与CK比, B15和B30显著提高葵花产量, 增幅分别为6.51%和9.44%。
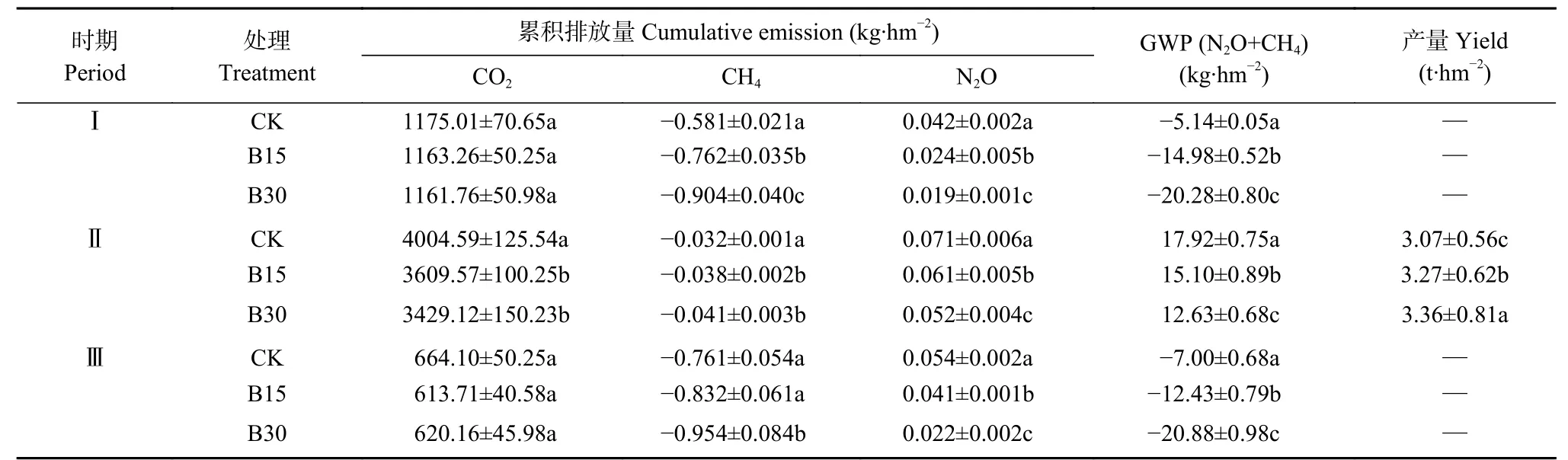
表2 不同生物炭用量下不同时期温室气体累积排放量、综合增温潜势(GWP)和葵花产量Table 2 Greenhouse gas cumulative emissions, global warming potential (GWP) and sunflower yield in different periods under different biochar rates
2.5 温室气体排放与环境因子的相关性
温室气体与环境因子的相关关系如表3所示。整个试验期的土壤CO通量与10 cm土壤温度有极显著正相关性(<0.01), 相关系数达0.80, 同时与20 cm土壤温度和10 cm土壤水分均呈显著正相关关系(<0.05), 相关系数分别为0.70和0.55。NO通量与10 cm土壤温度和20 cm土壤水分呈显著正相关关系(<0.05), 相关系数分别为0.58和0.59, 与其他因子无显著关系。CH通量则与土壤温度、水分、电导率、pH以及速效磷钾均无显著相关性。

表3 试验期温室气体排放通量与环境因子的相关性Table 3 Correlation between fluxes of greenhouse gases and environmental factors during the experimental period
3 讨论
3.1 施加生物炭对盐渍化土壤周年CO2排放通量的影响
微生物对土壤有机碳利用分解是产生土壤CO的主要原因, 土壤水分和温度是土壤CO产生的重要环境因子。从整个试验期来看, 土壤CO排放通量具有明显的阶段性差异, 作物生长期是CO排放高峰期, 而冻融期排放量较低。由于作物生长期土壤温度和水分较高, 土壤微生物利用作物根系分泌的营养物质及根表皮脱落物生长迅速繁殖, 使土壤CO排放量上升, 而冻融期土壤温度和水分较低,微生物活性较弱从而减缓了土壤CO排放速率。
本研究发现生物炭施入土壤中降低了试验期的土壤CO累积排放量, 主要原因是生物炭可以吸附土壤中的沉积物和土壤酶, 抑制有机碳转化和微生物活性, 降低了CO的排放; 其次, 生物炭提高了土壤pH, 相关性分析发现土壤CO与pH呈负相关关系, pH升高在一定程度上也会抑制土壤CO排放。这与刘翔等和Gao等的研究认为添加生物炭会增强土壤CO排放速率存在差异: 一方面原因是不同类型土壤内部有机质等组成成分可能不同, 微生物类型对土壤CO排放的影响存在差异; 另一方面原因是施加生物炭增强土壤CO排放仅存在短期效应, 初期生物炭本身含有大量土壤微生物利用的有机质, 从而提高土壤微生物的活性、增强了土壤呼吸作用, 后期生物炭促进了一些难以被土壤微生物分解的大分子物质形成, 如土壤腐殖质、碳水化合物等, 降低了微生物对有机质的利用量, 试验初期,在个别取样段同样监测到生物炭处理的土壤CO通量高于CK处理的现象, 但后期整体低于CK处理。此外, 随着生物炭添加量增大, 土壤的通气状况和营养状况得到进一步改善, 土壤呼吸也随着加强, 高量生物炭也会促进土壤CO排放。研究还发现, 添加生物炭对不同时间尺度上的土壤CO排放呈现出不同的效果。生物炭在作物生长期降低土壤CO累积排放量效果显著, 而在1 a冻融期和2 a冻融期无显著效果。苏德丽研究350 ℃温度条件下制备的生物炭(与本研究360 ℃制备的生物炭相近)发现,在冻融循环过程中该生物炭的中值粒径由198 μm减少到167 μm, 运用相关模型拟合后发现, 生物炭粒径和吸附系数有着显著的相关关系。因此推测本研究产生的结果可能是因为冻融作用极大影响了生物炭的吸附效果, 加之冻融期土壤温度和水分波动较小, 导致冻融期生物炭抑制土壤CO效果不明显。由于影响土壤CO排放的环境因素较复杂, 因此在研究土壤CO排放时除长时间监测外, 还需考虑生物炭陈化对土壤CO排放的影响。
3.2 施加生物炭对盐渍化土壤周年CH4排放通量的影响
农田土壤主要通过微生物厌氧发酵向大气中排放CH。本研究发现, 1 a冻融期、作物生长期和2 a冻融期土壤CH排放通量均处于较低的排放水平,并且大多表现为吸收状态。分析原因是盐渍化土壤含盐量较一般土壤高, 致使产甲烷细菌菌株活性会有所减弱, 而且也会降低土壤中的微生物活性, 从而抑制CH的产生。杨文柱等在内蒙古河套灌区同样发现, 不同盐碱程度的土壤均以吸收CH为主。从整个试验期来看, 生物炭处理均抑制了土壤CH的排放。生物炭本身具有较多的孔隙和具有使土壤中的氧化还原电位增加的能力, 从而改善了土壤的通透性和土壤C/N, 提高了甲烷氧化菌和嗜甲烷细菌的活性。同时本研究所用生物炭本身呈碱性(pH为8.75), 而产CH菌适合生存在酸性环境, 从而施加生物炭抑制了土壤CH的排放, 但CH产生环境中对产CH菌适宜的pH范围因土壤类型不同而不同, 并且CH排放通量与土壤pH间相关关系也不同。有研究发现, CH排放受土壤pH和微生物量碳影响, 且CH排放与pH呈现负相关。随老化过程持续进行, 生物炭pH降低, 则生物炭对CH减排效果有所削弱。本研究同样发现, 虽施加生物炭有效地抑制了土壤CH排放, 但抑制效果呈逐时期降低趋势, 通过分析土壤pH变化发现, 施加生物炭虽提高了土壤pH, 但效果不显著, 随着生物炭施加时间推移, 生物炭提高土壤pH效果逐渐降低。因此推测本试验生物炭可能还未达到老化程度, 但从长时间尺度来讲, 生物炭对CH减排效果会有所削弱, 因此生物炭对土壤CH产生与排放的影响机制仍需进一步探讨。
3.3 施加生物炭对盐渍化土壤周年N2O排放通量的影响
土壤产生和释放NO是由于硝化作用和反硝化作用机制。试验表明, 冻融期各处理土壤NO通量均处于较低水平: 一方面因为盐渍化土壤本身具有较高的pH, 使微生物NO还原酶降低, 抑制了土壤NO排放; 另一方面由于土壤温度降低, 土壤中微生物的硝化与反硝化能力随着土壤含水率减少而减弱, 使土壤NO通量逐渐降低。而在融解期和作物生长期NO排放达到高峰, 类似的研究结果在高寒草甸、落叶林等生态系统中同样存在。其原因是大量的融化水、土壤解冻水以及作物生长期雨水,使土壤处于厌氧环境, 主要以反硝化作用为主导, 因此NO排放增多; 加之作物生长过程中增加了土壤凋落物的输入量, 为NO产生菌提供了更为丰富的底物, 更有效地提高了NO产生率。
生物炭因其自身的化学性质与结构特点, 如较高的pH、巨大的比表面积及强吸附能力等, 直接或间接地影响硝化与反硝化过程, 从而影响土壤NO气体排放。本研究中施加生物炭显著减少了各时期的土壤NO累积排放量。其原因可能是生物炭能够吸附土壤中的无机氮, 降低了NO底物的可用性,且生物炭巨大的表面积, 更容易吸附导致NO增排的NH-N、NO-N和磷酸盐。也有研究表明, 施加生物炭会释放出更多的有机官能团和碳酸盐, 导致土壤pH增加, 在一定程度上降低了NO排放。本试验施加生物炭未显著提高盐渍化土壤pH, 且土壤NO通量与pH并无显著相关性, 说明土壤pH的变化并不是影响土壤NO排放的主要因素。但本试验却发现, 施加生物炭显著提高了各时期盐渍化土壤电导率, 使土壤盐分增加。根据已有研究表明,NO排放主要是在微生物的驱动下进行, 土壤盐分增加会抑制土壤酶活性和参与反硝化作用微生物活性,从而降低了NO排放。随着生物炭施入时间增加,生物炭经过2年冻融作用以及作物生长期降雨等自然老化后依然具有抑制土壤NO排放的作用, 这与de La等研究不一致, 该研究认为生物炭在田间自然老化过程中芳香碳含量下降、表面的含氧基团也会有所增加, 因此生物炭老化过程中NO减排效果会逐渐降低。分析原因可能是本试验周期内的生物炭仅是短时间尺度上的老化, 且土壤类型、生物炭种类等因素不同也会产生不同的结果。所以, 在今后的研究中需权衡生物炭老化进度和土壤NO排放与不同土壤类型、生物炭类型以及不同环境因子之间的关系。
3.4 施加生物炭对盐渍化土壤周年GWP和作物产量的影响
本研究发现, 生物炭的施入可显著降低盐渍化土壤周年温室气体综合增温潜势: 一方面因为单位质量CH和NO的全球增温潜势在100 a时间尺度上分别为CO的28倍和265倍, 通过表2可以看出施加生物炭均降低了土壤CH和NO累积排放量,从而使土壤GWP降低; 另一方面因为内蒙古河套灌区属于干旱、半干旱地区, 在旱地土壤中, 土壤NO累积排放量直接影响着GWP, 不同生物炭用量下不同时期间GWP的差异变化与土壤NO累积排放量差异一致, 因此施加生物炭对土壤具有一定的减排效果。研究还发现, 施入生物炭显著提高了河套灌区葵花的产量, 因为生物炭可以改善土壤的理化性质, 进而提高作物对土壤中氮素营养的吸收以及水肥利用效率, 从而提高作物产量。
4 结论
1)土壤周年CO排放通量具有明显的阶段性差异, 作物生长期是CO排放高峰期, 而冻融期CO排放量处于较低水平。与对照相比, 不同时期的生物炭处理抑制盐渍化土壤CO累积排放量效果存在一定性差异, 其中对作物生长期土壤抑制效果显著, 降幅达9.86%~14.37%, 但对1 a冻融期和2 a冻融期土壤抑制效果不显著。
2)整体而言, 试验期盐渍化土壤均以吸收CH为主, 且施加生物炭能够促进盐渍化土壤吸收CH。但随着生物炭施入时间的延长, 其降低土壤CH累积排放量的能力有所减弱, 降幅大小分别为2 a冻融期<作物生长期<1 a冻融期, 效果呈逐时期降低趋势。
3)不同处理周年NO排放通量的动态变化基本一致, 排放高峰均出现在土壤融解期和作物生长期,尤其作物生长期排放量占整个试验期排放量近一半。与对照相比, 生物炭处理对盐渍化土壤NO累积排放通量具有显著抑制作用, 3个时期的NO累积排放量大小均为CK>B15>B30, 降幅达14.08%~59.26%。
4)生物炭可显著降低盐渍化土壤周年温室气体综合增温潜势, 且2年冻融期各处理的综合增温潜势均为负值, 不具有增温效应。生物炭经过冻融作用后, 依然能够显著提高葵花产量, 其中以B30处理增产效果较优, 增幅为9.44%。
猜你喜欢
农业灾害研究(2022年1期)2022-05-07
环境(2021年5期)2021-06-20
文萃报·周五版(2020年37期)2020-10-12
看世界·学术上半月(2020年9期)2020-09-10
河北工业大学学报(2019年4期)2019-09-10
大气科学学报(2018年4期)2018-09-10
上海师范大学学报·自然科学版(2018年4期)2018-05-14
少儿科学周刊·儿童版(2015年12期)2016-05-16
农业工程技术·温室园艺(2009年4期)2009-07-24
农业工程技术·温室园艺(2009年11期)2009-05-17
