
板蓝根多糖抗病毒作用及其机制研究进展
2022-10-14 12:31:10李咸慰宋沁洁杨新荣李国峰李东辉吴红伟李越峰
中草药 2022年19期
李咸慰,宋沁洁,杨新荣,李国峰,李东辉,吴红伟,李越峰, 2
板蓝根多糖抗病毒作用及其机制研究进展
李咸慰1,宋沁洁1,杨新荣1,李国峰1,李东辉1,吴红伟1,李越峰1, 2*
1. 甘肃中医药大学,甘肃 兰州 730000 2. 甘肃省中药制药工艺工程研究中心,甘肃 兰州 730000
板蓝根多糖是板蓝根的主要成分,具有多种药理作用,其中抗病毒作用较为突出,可通过多种方式实现,如直接杀灭病毒和抑制病毒吸附、复制及蛋白表达,还可通过促进免疫细胞增殖分化、促进细胞因子释放、降低促炎因子水平以及参与红细胞免疫等方式增强机体的免疫作用,Toll样受体3(toll-likereceptor-3,TLR3)通路和Janus激酶(Janus kinase,JAK)/信号转导子和转录激活因子(signal transducer and activator of transcription,STAT)通路是其通过免疫调节方式抗病毒的重要信号通路,免疫佐剂的应用可为板蓝根多糖与其他抗病毒制剂的联合使用提供一定思路。主要综述了板蓝根多糖抗病毒作用及其机制的研究进展,以期为板蓝根多糖的临床应用提供参考。
板蓝根;板蓝根多糖;抗病毒;免疫;信号通路;作用机制
板蓝根是十字花科植物菘蓝Fort.的干燥根,具有清热解毒、凉血利咽的功效[1]。研究表明,板蓝根具有抗菌、抗炎[2]、抗病毒[3]、抗肿瘤[4]、免疫调节[5]、抗氧化[6]、调血脂[7]、保肝[8]等药理作用,因其抗病毒作用显著,在医药及畜牧业应用广泛。病毒作为一种常见的病原微生物,具有种类多、致病性强[9]、传染性强[10]、变异性强[11]和诱发其他疾病[12]等特点。研究发现,多糖作为板蓝根的主要成分,抗病毒效果显著且应用范围广泛,对流感病毒[3]、肝炎病毒[13]、伪狂犬病病毒[14]等多种病毒具有抑制作用。板蓝根多糖的抗病毒作用与其对机体免疫力的增强密切相关,也可通过对病毒的直接杀灭、干扰其感染、增殖过程等多种方式共同发挥作用。本文主要对板蓝根多糖抗病毒的药理作用及其机制的研究进展进行综述,以期对其在相关方面的使用提供参考。
1 直接杀灭作用
研究表明,板蓝根对病毒可起到直接杀灭的作用,一般通过体外细胞实验实现,将病毒与药液混合物同时接种于培养细胞,同时设置正常细胞和病毒感染细胞对照组,以四甲基偶氮唑蓝(methyl thiazolyl tetrazolium,MTT)比色法和空斑减少法检测细胞活性体现其直接杀灭病毒的效力。杨子峰等[15]提出,板蓝根提取物S-03主要成分为多糖,以空斑减少法评价其对不同亚型流感病毒的抗病毒作用,发现板蓝根提取物S-03对8种流感病毒均有一定程度抑制作用,直接作用效果显著且呈浓度、剂量相关性。研究表明板蓝根多糖可直接杀灭猪流感病毒H3N2,MTT法检测发现在药物质量浓度0.312 5~1.25 mg/mL时,抑制率为2.65%~6.59%[16];同时,发现板蓝根多糖可抑制、阻断和直接杀灭体外抗猪伪狂犬病病毒(pseudorabies virus,PRV),其中直接杀灭作用更为显著[17]。Tong等[18]研究发现,0.078 125、0.625 mg/mL板蓝根多糖对PRV的直接杀死率分别达到了32.214%、67.422%。此外,将猪繁殖和呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)制剂直接与不同浓度的板蓝根多糖混合,2 h后测定其与未混合板蓝根多糖病毒组的半数组织培养感染剂量,结果发现,板蓝根多糖可通过直接杀灭病毒以降低PRRSV的传染性,且呈浓度相关性[19]。综上,板蓝根多糖对多种病毒具有直接杀灭作用,效果显著且具有一定临床意义。
2 抑制病毒复制及吸附
病毒在体内发挥作用需经历多个环节,如吸附、脱壳、复制、成熟、释放等[16],除直接杀灭作用外,板蓝根多糖对病毒的复制过程也有抑制作用。乔艳艳[17]采用细胞实验,将板蓝根多糖接种于提前感染病毒的细胞中,以MTT法检测其与病毒对照组的细胞活性,发现0.625 mg/mL板蓝根多糖对PRV的抑制率可达30.84%。与张鹏宇[20]的研究结果一致,且张鹏宇[20]的研究还显示,板蓝根多糖单独使用时的半数抑制浓度为250 μg/mL,与黄芪多糖、干扰素同时使用时,抑制率可提升至70.26%。研究发现0.156~0.625 mg/mL板蓝根多糖对PRV复制的抑制率可达14.674%~30.840%[18]。以上研究表明,板蓝根多糖对病毒复制的抑制效果,与其浓度有关。
病毒血凝素是一种抗原性糖蛋白的血球凝集素,它可以使病毒与被感染的细胞相互吸附结合,Yang等[21]研究认为,板蓝根粗多糖可干扰病毒血凝素和细胞受体之间的结合,以此发挥对禽流感病毒H6N2、H7N3、H9N2及人季节性流感病毒H1N1、H3N2等不同亚型的抑制作用,且由于作用方式与M2抑制剂的不同,病毒不易出现耐药性;同时还推测,血凝素的构象差异导致其抑制人和禽病毒的复制时,对人季节性流感病毒效果更显著。另有研究指出,板蓝根多糖对人及禽流感病毒血凝素具有特异抑制作用,表现在对3型副流感病毒血凝素并无抑制[15]。
综上,板蓝根多糖对病毒的复制及吸附抑制作用显著,对病毒血凝素的抑制是其发挥阻断病毒吸附作用的重要方式。同时,李永亮[16]指出,板蓝根多糖对病毒吸附和穿入的阻断作用,还可能是其增强了细胞膜的稳定性,从而保护受感染细胞。
3 抑制病毒RNA和蛋白质的合成及表达
板蓝根多糖对PRRSV的抗病毒作用除抑制其与上皮样细胞MARC-145的黏附,还包括抑制病毒RNA和蛋白质的合成[19],使用聚合酶链式反应和酶联免疫吸附法分别测定板蓝根多糖对病毒RNA和GP5蛋白的影响,在板蓝根质量浓度为80 g/mL时,抑制率分别达57.6%、80.0%。Wang等[22]研究表明,当板蓝根多糖质量浓度超过50 μg/mL时,人乙型肝炎病毒(hepatitis B virus,HBV)感染细胞中,HBV DNA水平明显下降。秦静霖[23]研究指出,板蓝根多糖可抑制马立克氏病病毒(Marek’s disease virus,MDV)基因的表达及其gB蛋白的表达,且呈时间、浓度相关性。此外,板蓝根多糖可能通过抑制病毒mRNA的表达和病毒繁殖所需生物的合成过程,实现对猪流感病毒H3N2的抑制作用[16]。除此之外,对病毒RNA和蛋白合成与表达的抑制,板蓝根多糖也可通过免疫调节方式间接发挥作用。
4 免疫调节
4.1 促进免疫细胞的增殖
周寒鹏等[24]研究表明,板蓝根多糖可显著增强脾脏T细胞的增殖能力,ip板蓝根多糖对于小鼠的免疫功能有明显促进作用,还可增加正常小鼠的脾质量、淋巴细胞数及白细胞总数,对于小鼠抗体形成及细胞功能的增强有积极作用[25]。赵凤杰[26]研究同样显示ip板蓝根多糖可使正常患者的脾质量、淋巴细胞数、白细胞总数明显增加,同时,对于氢化可的松诱导的免疫功能抑制所致的脾指数、淋巴细胞数和白细胞总数的下降具有显著的改善作用。另有研究指出,板蓝根多糖可恢复环磷酰胺模型小鼠的胸腺指数,促进其脾淋巴细胞增殖[27]。此外,板蓝根多糖对猪脾淋巴细胞的增殖同样有促进作用[28]。
板蓝根凝集素是一种特殊的板蓝根多糖,可通过与细胞表面的糖蛋白结合,从而促进小鼠胸腺发育及胸腺细胞的增殖,进而促进T淋巴细胞、胸腺上皮细胞对胸腺素和细胞因子的分泌,提高机体免疫力[29]。另有文献指出,板蓝根凝集素的抗病毒作用与板蓝根血凝活性呈正相关[30]。以上研究表明,板蓝根多糖对免疫受损及正常机体的免疫器官增殖和免疫细胞总数的提升具有显著作用。
4.2 诱导免疫细胞分化
免疫细胞的分化是机体免疫作用的重要环节,受多种细胞因子的介导。白细胞介素(interleukin,IL)参与多种细胞的免疫过程,辅助T细胞(helper T cell,Th)对免疫细胞的诱导激活及免疫应答具有重要作用[31],在免疫反应中充当中间角色,其增殖分化受细胞因子、转录因子及代谢等多种因素影响[32]。乔艳艳[17]研究发现板蓝根多糖可通过抑制mRNA对Th1细胞应答的削弱作用和促进IL-12转录或翻译,以此实现Th0型细胞分化为Th1型细胞的诱导,使Th1细胞的免疫应答效应增强。张弩[33]研究也表明板蓝根多糖可显著提高小鼠淋巴细胞对IL-10的分泌,促进TH0细胞分化为TH2细胞,继而促进B细胞分化、成熟和增殖,使免疫球蛋白E(immunoglobulin E,IgE)和IgG类抗体大量生成,增强机体的体液免疫作用。
4.3 增强免疫细胞活性和促进抗炎因子分泌
研究发现,0.32 g/kg板蓝根多糖可增加IL-2和γ干扰素(interferon-γ,IFN-γ)的分泌,并提高巨噬细胞吞噬指数[34]。左娅等[35]研究指出,ip板蓝根多糖溶液可提升受流感病毒H1N1感染小鼠血清中抗体IgG的水平,提高小鼠脾脏细胞IFN-γ的水平,使小鼠抗流感病毒能力得到显著增强;此外还提出,板蓝根多糖可使小鼠外周血CD4+、CD8+细胞升高而达到抗病毒的目的。研究显示,板蓝根多糖可显著提高小鼠NK细胞的吞噬能力[33]。这与徐艺铨[36]的研究结果相一致。肿瘤坏死因子(tumor necrosis factor,TNF)是一种引起细胞死亡(凋亡)的细胞因子,耿婵娟[37]研究表明,用含板蓝根多糖100 mg/kg的饲料喂食后,小鼠体内IL-2水平显著提升,且小鼠血清中的IL-2和TNF-α水平与板蓝根多糖的使用剂量呈正相关,使用剂量越大,小鼠体内抗炎因子水平越高,免疫力越强。同时,板蓝根多糖对荷瘤小鼠的TNF-α、INF-γ和IL-2均有上调效应[4]。板蓝根多糖还可提高肠黏膜免疫球蛋白(IgA、IgM、IgG)的含量,抵抗肠道黏膜处的病毒入侵[38],对肠黏膜免疫具有积极作用。
4.4 抑制促炎因子及趋化因子的表达
促炎因子与趋化因子在免疫过程中具有诱导免疫细胞分化、募集免疫细胞等作用,病毒感染引起促炎因子及趋化因子的异常表达,导致机体产生炎症反应[39],如发热、疼痛、组织损伤等[40-41],其表达水平也成为判断病毒感染严重与否的重要标志物[42]。板蓝根多糖可通过抑制IL-6的表达降低受MDV感染细胞的炎症反应[24],还可抑制流感病毒诱导的促炎因子如干扰素诱生蛋白-10(interferon-inducible protein-10,IP-10)、趋化因子(C-X-C基元)配体5[chemokine (C-C motif) ligand 5,CCL5]等的释放[43]。
4.5 通过免疫调节抗病毒的重要信号通路
4.5.1 Toll样受体3(toll-likereceptor 3,TLR3)通路 由病毒激活的TLR3促炎信号通路可导致机体炎性反应增加和其他并发症的产生[44]。板蓝根多糖能有效抑制人流感病毒PR8/H1N1诱导的TLR3蛋白水平的上调,同时经板蓝根多糖干预后TLR3通路下游促炎因子(IP-10、IL-6、CCL5和CCL9)表达水平显著降低。推测板蓝根多糖可能是通过抑制TLR3通路上游的蛋白表达,降低机体促炎因子的分泌[43],见图1。
4.5.2 Janus激酶(Janus kinase,JAK)/信号转导子和转录激活因子(signal transducer and activator of transcription,STAT)通路 干扰素诱导的JAK/STAT通路参与多种免疫性疾病、炎症反应和感染性疾病[45]。其下游的抗病毒蛋白2′,5′-寡腺苷酸合成酶(2′,5′-oligadenylate stythetase,OAS)和黏病毒抵抗蛋白A(myxovirus resistance protein A,MxA)是其发挥作用的物质基础,同时,细胞因子信号转导抑制因子-1(suppressors of cytokine signaling-1,SOCS-1)和SOCS-3的过度表达可抑制其中间过程STAT的磷酸化及后续蛋白的表达[46],见图2。
板蓝根多糖可显著增加细胞内IFN-α水平,且呈剂量和时间相关性;同时,经板蓝根多糖干预后,细胞STAT-1、STAT-2、JAK1和Tyk2蛋白总量不变,但其磷酸化水平升高,SOCS-1和SOCS-3的蛋白表达被抑制,OAS和MxA表达水平显著增加[22],表明激活由干扰素诱导的JAK/STAT信号通路,是板蓝根多糖抗病毒的重要途径。
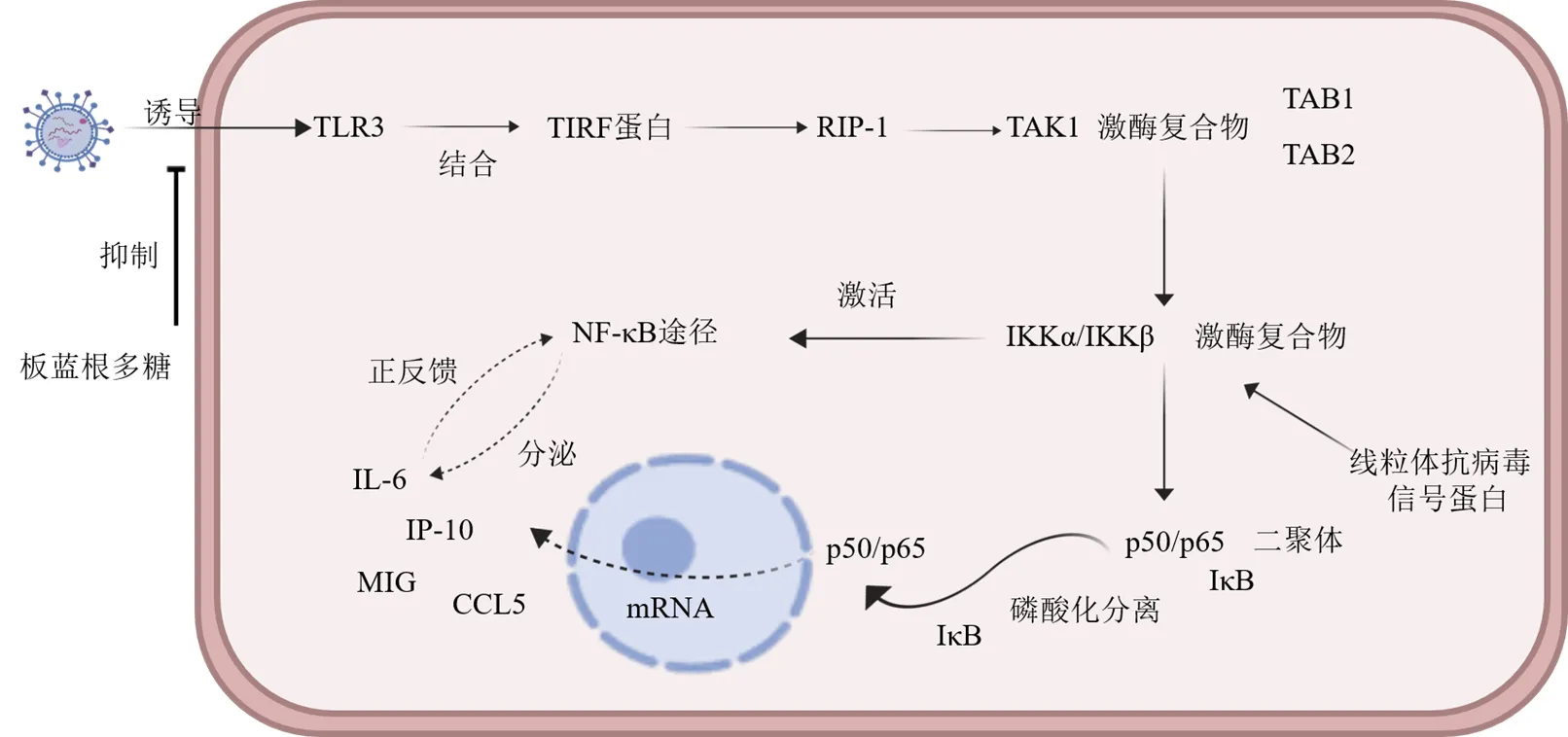
TIRF-含TIR结构域的接头分子 RIP-1-受体相互作用蛋白-1 TAK1-转化生长因子β激酶1 TAB-TAK1结合蛋白家族 NF-κB-核因子-κB IKK-IκB激酶 IκB-NF-κB抑制蛋白

图2 板蓝根多糖通过调节JAK/STAT通路发挥抗病毒作用机制
4.6 免疫佐剂
免疫佐剂作为免疫应答的辅助物质,可提升疫苗的使用效果。中药在疫苗佐剂中的应用也十分广泛,对于禽流感、狂犬病、口蹄疫、乙肝等多种病毒的疫苗具有较好的免疫辅助作用,见表1。
4.7 其他免疫调节作用
“红细胞免疫”是指红细胞(red blood cell,RBC)具有识别、储存、呈递抗原及清除免疫复合物(immune complex,IC)等功能,其细胞膜上的C3b受体,是发挥免疫功能的物质基础[53]。C3b受体与免疫复合物中的补体、酵母菌或抗体可黏附结合为花环状,以结合数量计算得出的红细胞花环率是评价其免疫功能强弱的主要方法[54]。研究发现,板蓝根多糖可使青脚麻雏鸡RBC-C3bR花环率明显提高[55],对兔RBC-C3b受体花环率和RBC-IC花环率也有明显升高作用[38]。
板蓝根多糖通过多途径调节免疫发挥抗病毒作用机制见图3。
表1 板蓝根多糖对不同疫苗的佐剂应用
Table 1 Adjuvant application of Isatidis Radix polysaccharide to different vaccines
多糖疫苗实验动物作用效果及机制文献 处理方式剂量/μg名称剂量/μg 水提醇沉(质量分数8.75%)150单纯疱疹病毒2感染的DNA疫苗pgD100雌性BALB/c小鼠脾脏T细胞增殖能力显著增强、IFN-γ和IL-10含量显著增加,血清中特异性IgG的抗体水平显著增加24 粗多糖提纯(IIP-A-1和IIP-2)200狂犬病重组病毒RCVS-11-G50 μL雌性BALB/c小鼠增强小鼠抗体滴度、增强免疫细胞分泌、诱导B细胞分化、增强多糖对血液中B细胞和T细胞募集的作用47 200口蹄疫病毒灭活疫苗Aftosa2雌性BALB/c小鼠IIP-2具有免疫原性,分别与Aftosa疫苗、H1N1病毒裂解疫苗和HbsAg作用免疫小鼠,均产生抗IIP-2IgG1型抗体13,48 200H1N1病毒裂解疫苗3 200乙肝病毒重组亚单位蛋白乙肝病毒表面抗原HbsAg2 IIP-A-1100H1N1流感疫苗3雌性BALB/c小鼠促进特异性抗体的生成、总抗体及亚类抗体滴度升高、促进脾脏、胸腺免疫细胞增殖及抗炎因子分泌。激活巨噬细胞,促进TNF-α的分泌49 水提醇沉10 mg∙mL−1新城疫Lasota弱毒疫苗0.3 mLAA肉鸡抗体效价与脏器指数显著升高50 多糖(质量分数≥10%)200 mg∙L−1鸡新城疫、传染性支气管炎二联活疫苗 黄羽肉鸡(雌雄均有)抗体滴度显著增强、血清IgA含量提高、促进外周淋巴细胞增殖、免疫器官指数上升51 粗多糖精制(质量分数69.7%)20、10 mg∙kg−1PRRSV灭活疫苗 二元杂交断奶仔猪增强疫苗免疫效果,且与剂量呈正相关28 水提醇沉4、8 mg∙mL−1新城疫病毒IV株疫苗 罗曼白鸡显著促进淋巴细胞增殖、血清抗体滴度显著增强52

图3 板蓝根多糖通过多途径调节免疫发挥抗病毒作用机制
5 结语与展望
中药抗病毒作用报道较多,但因其成分复杂,作用机制体现出多途径、多靶点的特点。板蓝根作为常用清热解毒中药,相关复方及中成药在生活中广泛使用。在严重急性呼吸综合征病毒及新型冠状病毒大规模传染时期,其具有一定疗效的宣传颇多,然而,媒体的过度“捧杀”反而使民众对板蓝根及传统中药的认知适得其反,只有对相关机制深入研究,才能合理指导临床用药。
板蓝根多糖既可以直接作用于病毒及其感染过程,在病毒感染细胞初期起到阻断及抑制作用,又可在感染病毒后,以多种途径参与机体免疫调节,间接地起到抗病毒作用,抗病毒方式的多样性与复杂性可能与其多种成分和对多靶点的共同作用有关。板蓝根多糖对多种病毒具有较好的阻断及抑制作用,其抗病毒作用具有广泛性。另外,板蓝根提取物及其制剂在抗病毒研究中的作用路径也与抑制病毒感染、提高免疫等方式有关[56],可推测与板蓝根的多糖成分相关,提示在使用板蓝根多糖抗病毒时,可根据药效物质基础,合理搭配其他成分及药物,相互补充作用,利用其抗病毒的广谱性及有效性,发挥更好的效果。
利益冲突 所有作者均声明不存在利益冲突
[1] 中国药典[S]. 一部. 2020: 214.
[2] 郝敬友, 霍墨涵, 梁蒙, 等. 基于抗菌及抗炎作用探究板蓝根微粉对雏鸡沙门菌病的防治效果[J]. 中国兽医学报, 2021, 41(12): 2468-2474.
[3] 何立巍, 杨婧妍, 侯宪邦. 板蓝根正丁醇部位抗病毒活性组分及相关化学成分研究 [J]. 中草药, 2017, 48(14): 2843-2849.
[4] 李吉萍, 朱冠华, 袁野, 等. 板蓝根多糖体内抗肿瘤作用与免疫功能调节实验研究 [J]. 天然产物研究与开发, 2017, 29(12): 2010-2016.
[5] 陈冉, 王婷婷, 李开铃, 等. 免疫调节抗病毒中药的特性与应用 [J]. 中草药, 2020, 51(6): 1412-1426.
[6] 农石生, 龚子龙, 周金花, 等. 板蓝根抗氧化成分及抗氧化性能研究 [J]. 中国野生植物资源, 2017, 36(3): 18-22.
[7] 张文友, 袁野, 李吉萍, 等. 板蓝根水提物预防大鼠高脂血症 [J]. 中成药, 2017, 39(11): 2225-2230.
[8] 胡天骄, 姜振, 张文友, 等. 板蓝根水提物对糖尿病大鼠早期肝损伤的影响 [J]. 中国现代应用药学, 2017, 34(2): 196-199.
[9] 金松, 孟宪臣, 薛亚梅, 等. 我国部分地区黄羽肉鸡群鸡传染性贫血病病毒的流行病学调查及致病性分析 [J]. 中国家禽, 2022, 44(2): 29-35.
[10] 杨孝坤, 李昱, 赵宏婷, 等. 新型冠状病毒感染不同阶段的传染性研究进展 [J]. 中华流行病学杂志, 2021, 42(1): 33-38.
[11] 姚晓文, 周玉霞, 王蓉, 等. 2019新型冠状病毒变异株的研究进展 [J]. 中华传染病杂志, 2022, 40(2): 111-115.
[12] 孙宏磊. 病毒感染诱发肺损伤的作用机制和研究进展 [J]. 现代畜牧科技, 2018(12): 7-8.
[13] 李海霞, 刘坤璐, 贾培媛, 等. 板蓝根多糖IIP-A-1和IIP-2作为疫苗佐剂的免疫原性 [J]. 中国药理学与毒理学杂志, 2019, 33(1): 22-29.
[14] 刘芳. 板蓝根多糖调节伪狂犬病毒对树突状细胞成熟及功能影响的作用研究 [D]. 郑州: 河南农业大学, 2014.
[15] 杨子峰, 王玉涛, 秦笙, 等. 板蓝根水提物S-03体外抑制甲、乙型流感病毒感染的实验研究 [J]. 病毒学报, 2011, 27(3): 218-223.
[16] 李永亮. 板蓝根多糖对肺气虚证小鼠抗H3N2型猪流感病毒试验研究 [D]. 郑州: 河南农业大学, 2015.
[17] 乔艳艳. 板蓝根多糖对伪狂犬病毒感染猪树突状细胞的调节作用研究 [D]. 郑州: 河南农业大学, 2016.
[18] Tong C, Chen Z W, Liu F,. Antiviral activities ofpolysaccharide against pseudorabies virus in swine testicle cells [J]., 2020, 20(1): 48.
[19] Wei Z Y, Wang X B, Zhang H Y,. Inhibitory effects of indigowoad root polysaccharides on porcine reproductive and respiratory syndrome virus replication[J]., 2011, 16(3): 357-363.
[20] 张鹏宇. 板蓝根、黄芪多糖与干扰素对猪伪狂犬病病毒的体外作用研究 [D]. 郑州: 河南农业大学, 2015.
[21] Yang Z F, Wang Y T, Zhong S,.inhibition of influenza virus infection by a crude extract fromroot resulting in the prevention of viral attachment [J]., 2012, 5(3): 793-799.
[22] Wang T B, Wang X W, Zhuo Y,. Antiviral activity of a polysaccharide from(Fortune) against hepatitis B virus (HBV)via activation of JAK/STAT signal pathway [J]., 2020, 257: 112782.
[23] 秦静霖. 板蓝根多糖体外抑制鸡马立克氏病病毒增殖的作用研究 [D]. 郑州: 河南农业大学, 2021.
[24] 周寒鹏, 陶薇, 傅婷, 等. 板蓝根多糖对单纯疱疹病毒-2型DNA疫苗免疫效果的影响 [J]. 中国生物制品学杂志, 2020, 33(6): 609-613.
[25] 刘丽芳, 李方洲. 板蓝根化学成分、药理及质量控制管理 [J]. 中国卫生产业, 2018, 15(13): 36-37.
[26] 赵凤杰. 浅谈板蓝根的药理研究及临床应用 [J]. 中国保健营养, 2017, 27(11): 132.
[27] 薛瑞, 章激, 曹军华, 等. 板蓝根多糖对小鼠免疫功能的调节作用 [J]. 中医药导报, 2012, 18(9): 94-96.
[28] 夏俊. 新疆PRRSV感染调查及中药多糖、共轭亚油酸钙对仔猪免疫调节的研究 [D]. 石河子: 石河子大学, 2017.
[29] 何鑫. 板蓝根的主要药理作用机制 [J]. 东方食疗与保健, 2017(2): 6-7.
[30] 孙琴, 马丽, 李兰, 等. 板蓝根中红细胞凝集效应组分的谱效关系研究 [J]. 中草药, 2012, 43(1): 125-130.
[31] 石笑瑜, 周蓓蓓, 来艺青, 等. T细胞免疫与宫颈癌关系的研究进展 [J]. 东南大学学报: 医学版, 2021, 40(6): 869-874.
[32] 张笑梅, 刘春燕, 邵宗鸿. 辅助T细胞亚群分化调控机制研究进展 [J]. 中国免疫学杂志, 2020, 36(24): 3045-3054.
[33] 张弩. 板蓝根多糖的化学分离及其免疫增强活性的研究 [D]. 武汉: 华中科技大学, 2008.
[34] Zhao Y L, Wang J B, Shan L M,. Effect ofpolysaccharides on immunological function and expression of immune related cytokines in mice [J]., 2008, 14(3): 207-211.
[35] 左娅, 代明, 王志勇, 等. 板蓝根多糖对小鼠抗流感病毒感染的作用 [J]. 华西药学杂志, 2008, 23(6): 666-667.
[36] 徐艺铨. 板蓝根多糖的化学分离及其免疫增强活性的研究 [J]. 生物技术世界, 2015, 12(7): 140.
[37] 耿婵娟. 板蓝根多糖的提取及其免疫调节作用研究 [D]. 大庆: 黑龙江八一农垦大学, 2012.
[38] 郭兵, 王海凤. 板蓝根多糖对兔球虫黏膜免疫和红细胞免疫的影响 [J]. 黑龙江畜牧兽医, 2016(1): 191-193.
[39] 张瑞华. 细胞自噬在H9N2流感病毒致肺损伤中的作用及其机制研究 [D]. 呼和浩特: 内蒙古农业大学, 2021.
[40] Turner M D, Nedjai B, Hurst T,. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease [J]., 2014, 1843(11): 2563-2582.
[41] 方奕程, 张黄琦, 谢继承, 等. 不同临床分型新型冠状病毒肺炎淋巴细胞比率、IL-6、IL-10水平的比较及临床意义 [J]. 浙江医学, 2021, 43(7): 763-765.
[42] 马俊娥, 崔湘铧, 刘芮, 等. 新型冠状病毒肺炎患者血浆CRP, PCT和IL-6水平检测的临床诊断价值 [J]. 现代检验医学杂志, 2021, 36(5): 105-109.
[43] Li Z T, Li L, Zhou H X,.polysaccharides inhibit influenza a virus and influenza A virus-induced inflammation via suppression of host TLR3 signaling[J]., 2017, 22(1): E116.
[44] Chen Y J, Lin J H, Zhao Y,. Toll-like receptor 3 (TLR3) regulation mechanisms and roles in antiviral innate immune responses [J]., 2021, 22(8): 609-632.
[45] 张茹薏, 刘怀鄂, 彭玉娟, 等. SOCS1/3与慢性乙型肝炎疾病进展及抗病毒治疗疗效的关系 [J]. 现代医药卫生, 2021, 37(22): 3842-3846.
[46] Vlotides G, Sörensen A S, Kopp F,. SOCS-1 and SOCS-3 inhibit IFN-alpha-induced expression of the antiviral proteins 2,5-OAS and MxA [J]., 2004, 320(3): 1007-1014.
[47] Zhang W J, Zheng X X, Cheng N,.root polysaccharides as adjuvants for an inactivated rabies virus vaccine [J]., 2016, 87: 7-15.
[48] 王玉霞, 朱婷, 李倩, 等. 板蓝根多糖疫苗佐剂的候选药研究[A] // 2016全国多糖研讨会论文摘要集 [C]. 北京: 中国化学会, 2016: 13.
[49] 文松, 李倩, 贾培媛, 等. 板蓝根α-葡聚糖佐剂提高H1N1流感疫苗免疫小鼠体液免疫和细胞免疫功能 [J]. 中国药理学与毒理学杂志, 2014, 28(6): 850-856.
[50] 陈光明, 李巨银, 颜友荣, 等. 板蓝根多糖的制备及其对鸡新城疫Lasota疫苗的免疫效果研究 [J]. 安徽农业科学, 2014, 42(36): 12920-12921.
[51] 鲁振国, 李美娣, 武力, 等. 玉竹多糖和板蓝根多糖对环磷酰胺致雏鸡免疫抑制的调节作用研究 [J]. 中兽医医药杂志, 2021, 40(1): 74-77.
[52] Kong X F, Hu Y L, Rui R,. Effects of Chinese herbal medicinal ingredients on peripheral lymphocyte proliferation and serum antibody titer after vaccination in chicken [J]., 2004, 4(7): 975-982.
[53] 李悦, 石莉红. 红细胞免疫调控功能的研究进展 [J]. 中国细胞生物学学报, 2022, 44(1): 59-67.
[54] 高如意. 藏药秦艽花黄酮对小鼠红细胞免疫功能影响的实验研究 [D]. 成都: 西南民族大学, 2021.
[55] 王量, 范红结, 姚毅, 等. 板蓝根多糖对雏鸡红细胞免疫黏附功能的研究 [J]. 黑龙江畜牧兽医, 2014(6): 89-90.
[56] 李宁, 钱秀玉, 聂黎行, 等. 中药抗冠状病毒活性和机理方法研究进展 [J]. 药物分析杂志, 2020, 40(9): 1527-1535.
Research progress on antiviral effect and mechanism ofpolysaccharide
LI Xian-wei1, SONG Qin-jie1, YANG Xin-rong1, LI Guo-feng1, LI Dong-hui1, WU Hong-wei1, LI Yue-feng1, 2
1. Gansu University of Traditional Chinese Medicine, Lanzhou 730000, China 2. Gansu Research Center for Chinese Medicine and Pharmaceutical Technology Engineering, Lanzhou 730000, China
polysaccharide is the main component of Banlangen (), which has a variety of pharmacological effects. Among them, the anti-virus effect is more prominent, which can be achieved in a variety of ways, such as direct killing, inhibiting virus adsorption, replication and protein expression. It can also enhance the immune function of the body by promoting the proliferation and differentiation of immune cells, promoting the release of cytokines, reducing the level of proinflammatory factors and participating in erythrocyte immunity, TLR3 and JAK/STAT are important signal pathways for anti-virus through immune regulation. The application of immune adjuvant can provide some ideas for the combination ofpolysaccharide and other antiviral agents. Research progress on antiviral effect and mechanism ofpolysaccharide were reviewed in this paper, in order to provide reference for clinical use ofpolysaccharide.
;polysaccharide; antiviral; immunity; signal pathway; mechanism of action
R282.710.5
A
0253 - 2670(2022)19 - 6227 - 07
10.7501/j.issn.0253-2670.2022.19.028
2022-06-20
国家自然科学基金资助项目(81960713);国家自然科学基金资助项目(82160750);国家药品监督管理局中药材及饮片质量控制重点实验室项目(2020GSMPA-KL15);甘肃省教育厅产业支撑计划项目(2021CYZC-21);甘肃省中药制药工艺工程研究中心开放课题(ZYGY202003);甘肃省科技计划(创新基地与人才计划)基础研究创新群体项目(21JR7RA569)
李咸慰,男,硕士研究生,研究方向为中药及复方加工炮制机制及活性成分。E-mail: 2471446785@qq.com
李越峰,女,教授,博士生导师,博士后,研究方向为中药及复方加工炮制机制及活性成分。E-mail: 2126059063@qq.com
[责任编辑 崔艳丽]
猜你喜欢
肝博士(2022年3期)2022-06-30 02:48:52
世界科学技术-中医药现代化(2021年9期)2021-12-31 03:31:24
肝博士(2021年1期)2021-03-29 02:32:14
肝博士(2020年4期)2020-09-24 09:21:26
疯狂英语·初中天地(2020年3期)2020-05-21 03:36:52
天然产物研究与开发(2018年6期)2018-07-09 06:01:28
中成药(2018年3期)2018-05-07 13:34:45
益寿宝典(2018年35期)2018-01-26 17:00:09
中成药(2017年12期)2018-01-19 02:06:48
中成药(2014年11期)2014-02-28 22:29:50
