
气候变化和人类活动干扰下骆驼刺潜在分布格局变化特征
2022-10-12 13:50马倩倩吕光辉曾凡江
生态学报 2022年18期
尹 辉,田 聪,马倩倩,吕光辉,曾凡江,*
1 中国科学院新疆生态与地理研究所新疆荒漠植物根系生态与植被修复重点实验室,乌鲁木齐 830011 2 中国科学院新疆生态与地理研究所荒漠与绿洲生态国家重点实验室,乌鲁木齐 830011 3 新疆策勒荒漠草地生态系统国家野外科学观测研究站,策勒 848300 4 新疆大学资源与环境学院,乌鲁木齐 830046 5 中国科学院大学,北京 100049 6 新疆畜牧科学院草业研究所,乌鲁木齐 830000
气候变化和人类活动是全球变化的两个重要方面,它们对生物多样性和生态系统的可持续发展具有重要影响[1—2]。气候条件是决定物种分布的最重要原因,它影响着植物的生长、发育、物候等众多方面。不断加剧的气候变化有可能造成某些物种灭绝,从而引起生态系统失衡,严重威胁人类的生存[3]。反过来,人类活动尤其是过度开采自然资源、过度改变土地利用类型也会对物种生存产生严重的影响[4]。因此,利用气候变量和人类活动变量作为重要的环境因素能够对物种分布区的变化进行良好的预测。
生态位模型正是基于物种的存在数据和环境因素之间的相关性,通过一定的算法来推算物种的生态需求,从而预测物种的潜在分布区[5]。最大熵模型(MaxEnt)将物种和其环境因素看成一个系统,计算该系统具有最大熵时的状态参数,从而推测物种的潜在分布区[6—7]。最大熵模型自构建以来,对海洋、陆地、荒漠生态系统的大量动植物物种进行了广泛的预测。结果表明其预测出的潜在分布区结果较准确,并能在存在数据相对较少的情况下获得较准确的信息,因而广泛应用于濒危物种保护、入侵物种防御等方面[8]。在荒漠区植物的研究中,张华等人运用最大熵模型预测了气候变化对干旱区胡杨潜在分布区的影响[9],塞依丁等人运用最大熵模型预测了人类活动和气候变化背景下干旱区沙拐枣的潜在分布区的变化特征[10]。最大熵预测结果与实际情况较接近,获得了良好的效果[9—10]。
近年来,大量的研究报道指出,随着气候变化和人类活动的不断加剧,全球荒漠化进程显著加快,预计到21世纪末,全球旱地面积将达到陆地总面积的56%,而且旱地扩张以发展中国家为主[11]。骆驼刺是一种耐干旱、耐高温、耐贫瘠的典型的荒漠植物,它是我国干旱区的荒漠和戈壁上的优势物种,对维护干旱区生态平衡、防止沙漠扩张发挥着至关重要的作用[12]。骆驼刺对干旱环境具有极强的适应性[13],因而在旱地面积扩张的背景下,骆驼刺的适宜生境应当显著扩张。然而,杨霞等人2017年已对骆驼刺气候变化背景下的潜在分布区进行了预测,该研究表明,气候变化使得骆驼刺的适宜生境显著收缩[14]。值得注意的是,该研究用了19个生物气候变量对骆驼刺的潜在分布区进行预测,且未来气候情景运用国际耦合模式比较计划的第五阶段(Coupled Model Intercomparison Project 5,CMIP5)的RCP2.6和RCP8.5情景进行,模型运算时也未进行优化,因此模型预测结果有可能出现偏差。由于模型所用的环境因素及模型参数对模型预测结果具有重要的影响,近年来,涌现出大量对MaxEnt模型进行优化校准的文献资料,试图提高模型预测的准确度与精确度[15]。2020年,CMIP6的生物气候因素数据公布,CMIP6是目前参与的模式数量最多、设计的科学试验最为完善、所提供的模拟数据最为庞大的一次[16—17]。CMIP6充分利用上世纪末气候反转的因素,使得该气候变化模型在未来预测时更加贴合实际[18—19]。四个共享社会经济路径情景(SSPs:126,245,370,585)也给模型预测带来了新鲜血液。且众多研究指出,人类活动是造成骆驼刺生境被破坏的重要原因[12,20],故本研究预将人类活动强度的影响加入模型。相信新的环境数据的加入将对模型预测结果产生重要的影响。综上所述,本研究预对CMIP6的未来气候情景和人类活动情景下骆驼刺的潜在分布区进行预测。以揭示骆驼刺的潜在分布区的变化特征及其影响因素,确定其是否适合成为我国荒漠化防治的优良物种。
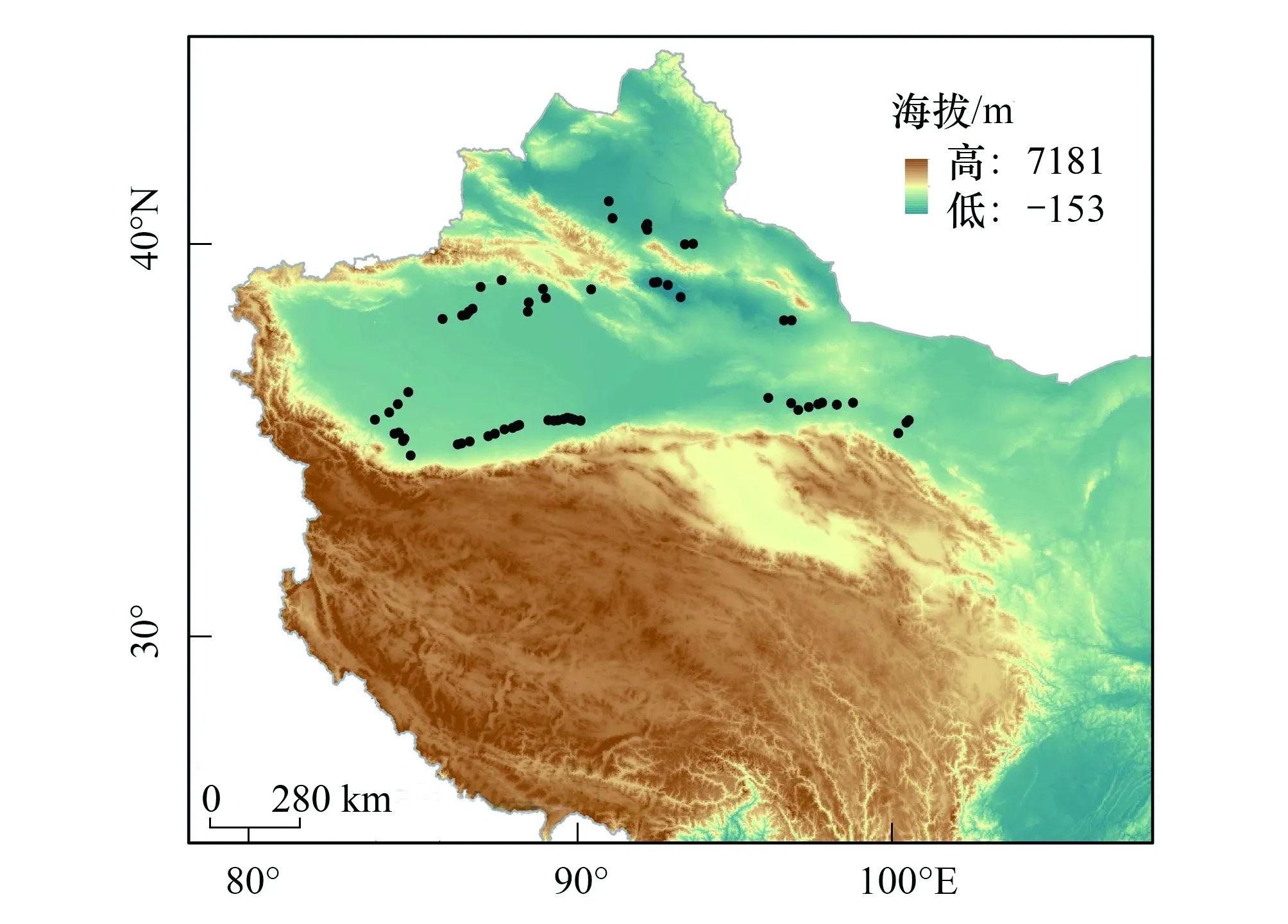
图1 骆驼刺发生点记录Fig.1 Occurrence records of A. sparsifolia
1 材料与方法
1.1 骆驼刺分布数据来源
为了获得骆驼刺详细的经纬度分布数据,通过查阅文献及全球生物多样性信息平台(GBIF,www.gbif.org),中国数字植物标本馆(CVH, http://v5.cvh.org.cn/),国家标本平台(NSII,http://www.nsii.org.cn/)等网站,运用googleearth软件推演标本位置的经纬度坐标。结合实地调查,获得了90个骆驼刺的发生数据。为了防止物种分布数据空间自相关带来的影响,并匹配2.5 arc-minutes分辨率的气候数据,本研究在直径5 km范围内仅保留一个数据点[21],最终获得61个骆驼刺发生点坐标(图1),用于最大熵模型的构建。
1.2 环境数据的来源与筛选
19个生物气候变量及1个高程数据,被认为是影响物种潜在分布区最重要的环境因子,常常用于生态位模型的构建[22]。分辨率为2.5 arc-minutes的19个生物气候变量(bio1—bio19)和高程数据(elev)来源于WorldClim2.0数据库。为了避免环境因子的多重共线性产生的过拟合现象,通过综合分析环境因子贡献率、环境因子间的相关性及环境因子的生态学意义,对19个生物气候因子进行筛选[23]。剔除相关系数高于0.8,且贡献率相对较小的环境因子[22],最后筛选出6个优化的环境因子用于建模。
针对未来气候因素,从WorldClim 2.0数据集中下载国家气候中心的气候系统模型(BCC-CSM2-MR)[16—17],CMIP6下的四个共享社会经济路径(SSP126,SSP245,SSP370,SSP585)。SSP126代表低辐射强迫情景,SSP245代表中等稳定情景;SSP370代表中高端强迫情景,SSP585代表高强迫情景[24]。本文欲展示不同时期骆驼刺潜在分布区的变化规律及其质心转移特点,故下载了2021—2040,2041—2060,2061—2080,2081—2100四个时期的19个生物气候因子的栅格数据,提取出与当代对应的6个优化的生物气候因子用于构建模型。
人类活动强度数据来源于国际地球科学信息网络中心(CIESIN)的人类足迹(human footprint)数据层。该数据集由9个数据层创建而成,涵盖人口密度、人类土地利用和基础设施建设(建筑区、夜间灯光、土地利用/土地覆盖)和人类访问(海岸线、公路、铁路、河流通航)等方面。能够客观、全面地反映人类活动强度及其空间分布[25]。该数据为Esri.Grid格式,将该数据置入ArcGis10.0软件中,经过坐标变换、裁剪、配准、格式转换等工作,使其与生物气候因子格式一致,用于MaxEnt模型的构建。
1.3 最大熵模型的优化、建立及评价
运用R3.6.3软件的“kuenm”包优化Maxent模型参数,其中倍频(regularization multiplier,RM)从0.5到4(增量0.5),特征组合(feature classes,FC)采用以下组合(L,LQ,H,LQH,LQHP,LQHPT,其中L:线性,Q:二次,H:铰链,P:积,T:阈值)[9];运用“kuenm”包对以上参数组合进行测试,通过Akaike信息量准则的delta AICc值评估模型的拟合度及复杂度,选择delta AICc最小的参数组合用于模型构建[26],并运用受试者特征曲线(Receiver Operating Characteristic curve,ROC)下面积(AUC值)评价和验证模型结果的准确性[19],AUC值小于0.6表示模型失败,0.6—0.7较差,0.7—0.8公平,0.8—0.9良好,大于0.9优秀[27]。最后将骆驼刺发生点的数据和优化后的环境因子导入MaxEnt模型中,运用优化后的模型参数组合,重复运行10次,获得骆驼刺当代潜在分布区预测结果。进行ROC检验,并运用刀切法获得环境因子贡献率。将预测结果置入ArcGis10.8软件中进行重分类,训练灵敏度加特异度的阈值被认为是一种更稳健的划分物种存在与否的方法[28],本研究的10次模型预测结果的训练灵敏度加特异度的阈值为0.19±0.06,故本研究将骆驼刺分布区划分为四个适生等级:非适生区(0—0.2),低适生区(0.2—0.4),中适生区(0.4—6),高适生区(0.6—1)。最后用ArcGis10.8对预测结果进行可视化展示。
未来气候条件下和人类活动条件下骆驼刺潜在分布区预测的数据处理与建模方法与当前气候条件的方法一致。
1.4 基于CMIP6未来气候情景的骆驼刺潜在分布区的变化及质心转移特点
将Maxent模型生成的ASCⅡ文件导入ArcGis10.8软件,转换成栅格数据后,计算出不同适生等级的骆驼刺分布区在未来不同的情景下与当代发生的面积变化,运用ArcGis10.8软件的SDM工具包比较未来与当代情景下适生区的变化区域与范围,获得骆驼刺在未来的扩张和收缩面积及地理位置。并以0.2为阈值,将其转化成二进制数据,运用SDM工具箱计算适生区的质心的位置及其变化方向。
2 结果与分析
2.1 生物气候因子的优化及模型精度检验
通过ROC检验,模型AUC值均大于0.9(图2),说明模型预测的准确性极高。

图2 有无人类活动干扰下,模型预测结果的ROC检验Fig.2 ROC curve and AUC values of the MaxEnt model without or without human disturbance
刀切法的结果表明,无人类活动条件下,年降水量对骆驼刺潜在分布区的贡献最高,达30.4%;最冷季度平均温次之,达22.5%;平均气温日较差、年均温和温度季节性的贡献率相仿,分别为16.4%,16.1%和12.7%。降水的季节性和高程的贡献率最小,为1.1%和0.9%(表1)。有人类活动条件下,年降水量、人类活动强度、高程是影响骆驼刺分布的最重要因素,其贡献率分别为:63.8%、17.4%和13.7%。

表1 建模的生物气候因子的相对贡献率排序
2.2 当代骆驼刺的适宜生境分布
无人类活动干扰的情景下,骆驼刺的潜在分布区主要位于新疆维吾尔自治区大部分地区,甘肃省北部,内蒙古自治区西部及青海省北部地区(图3)。高适生区主要在新疆南部地区。高适生区约40.06万km2,占总适生区的30.28%。中适生区面积约46.67万km2,占总适生区的35.26%。低适生区面积约45.56万km2,占总适生区的34.44%(表2)。人类活动干扰大大降低了骆驼刺的适生区,且适生区呈现破碎化状态(图3)。适生区以新疆为主,甘肃省北部和内蒙古西北部也有分布,青海省为非适生区。高适生区仅10.90万km2,占总适生区的15.28%。中适生区面积约26.76万km2,占总适生区的37.53%。低适生区面积约33.65万km2,占总适生区的47.19%(表2)。

图3 有、无人类活动干扰下,当代骆驼刺不同等级的潜在分布区Fig.3 The potential distribution areas of different grades of A. sparsifolia in current
2.3 影响骆驼刺潜在适宜分布的生物气候因子分析
根据环境因子响应曲线(图4),骆驼刺适生区的生物气候因子阈值为:年均温5.9—15.2 ℃,平均气温日较差大于13.4 ℃,温度季节变化106—161,最冷季度平均温-12.3—-0.2 ℃,年降水量18—145 mm,降水的季节性38—118。骆驼刺高适生区的范围很窄,在年均温9.8—12.2 ℃,平均气温日较差大于15.9 ℃,温度季节性变化1175—1281,最冷极度平均温度-6.9—-4.0 ℃,降水量30—49 mm,降水的季节性69—90的区域。

表2 不同等级的适生区面积及占总适生区面积的百分比

图4 骆驼刺的环境因子响应曲线Fig.4 Response curves for the probability of presence for A. spasifolia红线显示模型运行10次获得的平均值,蓝色的边界表示10次运算的标准差(SD)
2.4 CMIP6情景下,未来四个时段骆驼刺不同等级的适生区面积变化
四个共享社会经济路径下,骆驼刺各适生等级的面积变化趋势基本一致。高适生区面积显著增加,低适生区和中适生区面积减少(图5)。低强迫情景下,适生区面积几乎不随时间推移发生显著变化。而中高强迫情景和高强迫情景下,骆驼刺的低适生区面积随时间推移显著下降,而高适生区面积随时间推移显著增加(图5)。
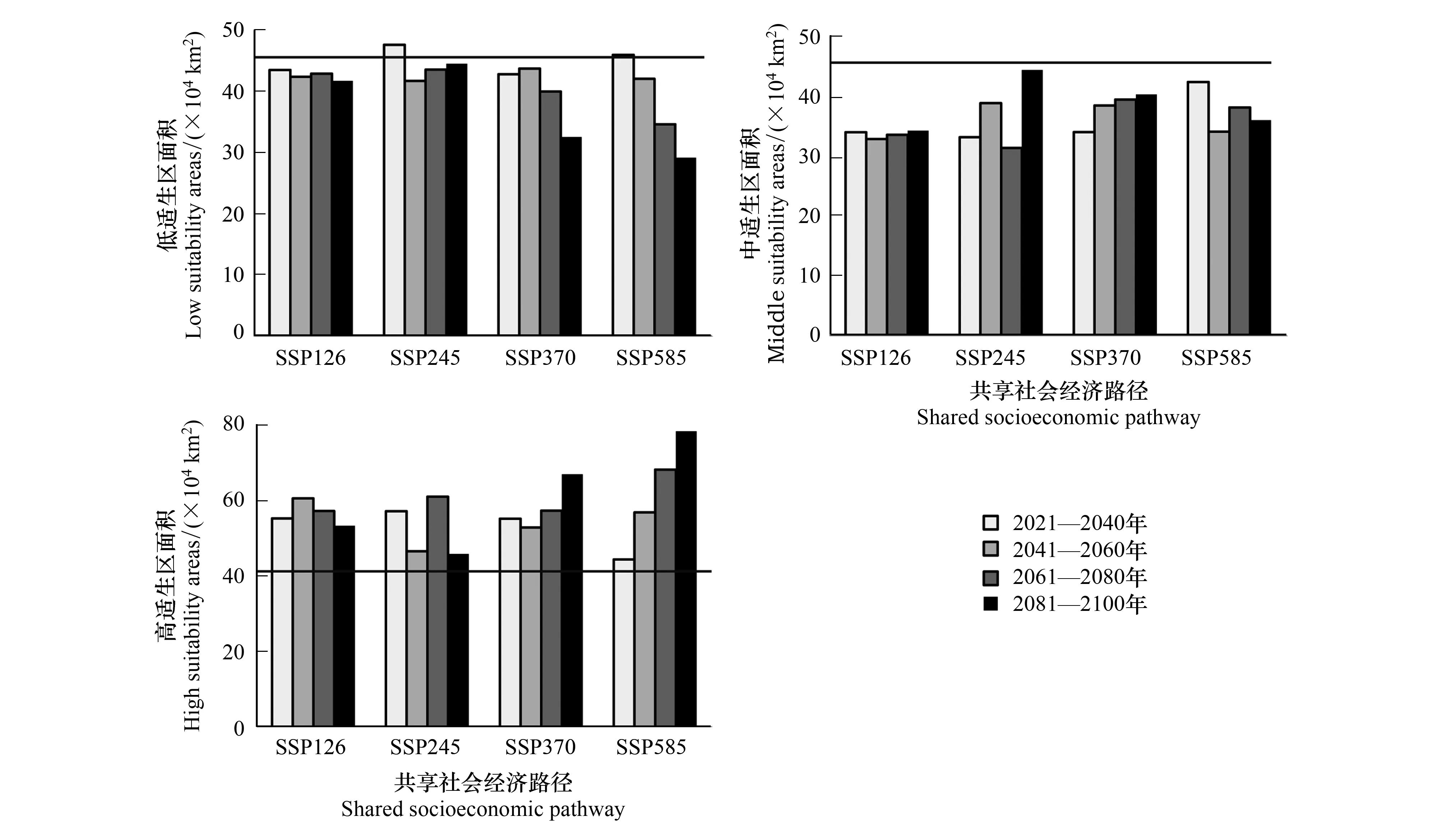
图5 四个共享社会经济路径(SSP126,SSP245,SSP370,SSP585)下,未来四个时间段,骆驼刺在我国各适生等级的面积变化Fig.5 Under four shared socio-economic paths (SSP126, SSP245, SSP370, SSP585), the area change of A. sparsifolia in each suitable level in China in the future four time periods 黑线表示当代骆驼刺的面积
2.5 CMIP6未来气候情景下,骆驼刺适生区的变化范围及质心转移特征
在SSP126情景下,2080之前的三个年代与当代相比,骆驼刺的适生区呈显著的增加趋势,而2081—2100年骆驼刺的适生区却显著下降。四个时间段的变化区域基本一致,扩张区主要位于青海省北部、甘肃省北部及内蒙古西北部地区,收缩区主要在内蒙古中西部地区,适生区的边缘。随时间推移,质心逐渐向西转移(图6)。
在SSP245情景下,随时间推移,骆驼刺的适生区呈现先增加后减少再增加的趋势。质心转移方向也呈现先东后西再转东的不规则变化特点。与当代相比,2041—2060年,骆驼刺在内蒙古的潜在适生区发生了大面积的收缩。而其他年代,骆驼刺的适生区以扩张为主,最显著的扩张区域为青海省北部和内蒙古的西北部(图7)。
在SSP370情景下,随时间推移,骆驼刺的适生区逐渐增加。骆驼刺向新疆北部、青海省北部及内蒙古西北部扩张。收缩区域仍以内蒙古中西部、骆驼刺适生区边缘为主。质心向西逐渐转移(图8)。
在SSP585情景下,骆驼刺的适生区逐渐增加,扩张区域仍旧以青海省北部、内蒙古西北部为主,新疆北部亦有较大面积的扩张,收缩区域仍旧以内蒙古中西部、骆驼刺适生区边缘为主。质心向西转移(图9)。

图6 SSP126情景下,4个时间段(2021—2040,2041—2060,2061—2080,2081—2100)与当代相比,骆驼刺适生区的变化范围及质心转移路径Fig.6 The variation range and centroid distribution transfer of fitness areas between four future periods (2021—2040,2041—2060,2061—2080,2081—2100) and current under SSP126 scenario for A. sparsifolia 质心转移路径用白色箭头的线表示。柱状图显示扩张区与收缩区的面积差

图7 SSP245情景下,4个时间段(2021—2040,2041—2060,2061—2080,2081—2100)与当代相比,骆驼刺的中、高适生区的变化范围及质心转移路径Fig.7 The variation range and centroid distribution transfer of fitness areas between four future periods (2021—2040,2041—2060,2061—2080,2081—2100) and current under SSP245 scenario for A. sparsifolia 质心转移路径用白色箭头的线表示;柱状图显示扩张区与收缩区的面积差

图8 SSP370情景下,4个时间段(2021—2040,2041—2060,2061—2080,2081—2100)与当代相比,骆驼刺的中、高适生区的变化范围及质心转移路径Fig.8 The variation range and centroid distribution transfer of fitness areas between four future periods (2021—2040,2041—2060,2061—2080,2081—2100) and current under SSP370 scenario for A. sparsifolia 质心转移路径用白色箭头的线表示;柱状图显示扩张区与收缩区的面积差

图9 SSP585情景下,4个时间段(2021—2040,2041—2060,2061—2080,2081—2100)与当代相比,骆驼刺的中、高适生区的变化范围及质心转移路径Fig.9 The variation range and centroid distribution transfer of fitness areas between four future periods (2021—2040,2041—2060,2061—2080,2081—2100) and current under SSP585 scenario for A. sparsifolia 质心转移路径用白色箭头的线表示;柱状图显示扩张区与收缩区的面积差
3 讨论
3.1 骆驼刺在当代的潜在适生区及关键的影响因素分析
年降水量是影响骆驼刺分布的最重要的生物气候因子,在有人类活动干扰的情况下,其影响作用更显著,年降水量、人类活动强度和高程对骆驼刺分布区的贡献率达94.9%。值得注意的是,骆驼刺适生区的年降水量范围非常狭窄,在18—145 mm范围内,高适生区的年降水量范围仅30—49 mm,如此低的降水范围内,其它物种往往很难生存。所以在年降水量低于36 mm的塔克拉玛干沙漠南缘和吐鄯托盆地,骆驼刺能够形成单种群群落。而在塔克拉玛干沙漠腹地,骆驼刺不能生存的环境下,便是寸草不生的沙漠。人类活动强度很好地勾画出了干旱区寸草不生的沙漠腹地。从模型结果来看,人类活动干扰大大降低了骆驼刺的潜在适生区面积,使其适宜生境碎裂化。这一现象有两个主要成因:一、人迹罕至的沙漠腹地寸草不生,只有在沙漠边缘的绿洲地带有植被覆盖,此处也为人类活动相对频繁的地带,因此加入人类活动强度使模型结果更合理。MaxEnt模型虽然是众多生态位模型中相对准确的一种,然而模型的模拟结果往往会比实际面积偏大[29],这与模型选取的环境变量关系很大,正如本研究中加入人类活动强度因素之后,适生区面积显著减小。对照中国植被地图,发现仅用生物气候因素和海拔因素预测的适生区内有大面积无植被区域,但该预测结果仍旧有很重要的指导意义。因为在广阔的无植被区域,其气候条件适宜骆驼刺的生长,可选用骆驼刺作为该区域的沙漠化防治、或沙漠公路保护的优良植被。二、人为因素,如人口压力、土地利用方式的转变、基础设施建设等均会对骆驼刺的分布产生重要的负面影响,可能造成骆驼刺适宜生境的丧失。研究指出,人类大量开采地下水造成地下水位下降,严重影响了骆驼刺的生存及自然更新[12]。且开荒种地直接造成骆驼刺栖息地减少或丧失。前人对干旱区沙拐枣的研究也得出相似的结论[10],说明干旱区人类活动可能对当地物种的适宜生境造成了显著的负面影响。因此在该区域的物种保护及合理开发的过程中,一定要密切关注人类活动的影响。
除去年降水量和人为因素的影响以外,最冷季度平均温、年均温、降水的季节性、温度日较差均是影响骆驼刺生存的重要原因,其中以最冷季节平均温影响最为显著。温度决定了植物的物候和生活史类型,同时直接影响植物生理生化过程。我国西北干旱区位于高纬度地区,此处植被类型主要为落叶乔灌木、多年生草本和一年生草本植物为主,生长季主要在4—10月[30]。在寒冷的冬季到来之前,骆驼刺的地上部分全部死亡,靠地下芽越冬[20,31]。因此最冷季度平均温并未直接影响骆驼刺的生长,而可能决定骆驼刺的物候期,从而对其分布产生重要的影响。温度日较差大是荒漠和戈壁地区的显著特点,在极端干旱区,部分植物的叶片能够直接吸收微量降水或昼夜温差导致的微量雾水或露水,作为水分补给[32]。有研究表明,在空气水分较为充足、辐射较弱的清晨,骆驼刺的气孔全部张开,表现出高光合、高蒸腾的特征,而在炎热的午后,骆驼刺气孔导度明显下降,光合作用速率和蒸腾作用降到最低。骆驼刺在夜间能够获得相对充足的水分补给,从而在整个生育期内,没有表现出显著的水分亏缺[33]。因此虽然降水对骆驼刺分布起最重要的作用,但是温度范围及温度日较差对骆驼刺的生存依然起到不可忽视的作用。
3.2 CMIP6情景下骆驼刺未来分布区的变化特点
骆驼刺是一种典型的荒漠植物,具有耐高温、耐旱、耐盐的特性,其生态适应性强,因而可以形成荒漠生态系统的优势种[12—13,31—34]。在未来气候变化情景下,骆驼刺显著的扩张,其扩张方向主要是高海拔的青海省北部地区和高纬度的新疆北部及内蒙古西北部地区。据预测,荒漠植物蒙古沙拐枣与骆驼刺在气候变化情景下具有类似的规律,它们会逐渐向高纬度和高海拔地区扩张但可以保留当代栖息地。然而张华等对胡杨潜在分布区的研究结果表明,在未来气候变化的条件下,胡杨将逐渐丧失中、低纬度的栖息地,而向高纬度栖息地转移[9]。对干旱区裸果木的研究发现,在气候变化情景下,裸果木也会逐渐丧失现有栖息地,而向水分相对充裕的地区转移[35]。众多研究指出,在未来气候情景下,大多植被将向高纬度和高海拔地区转移或扩散,然而地处高纬度地区的干旱区由于降水匮乏并不能成为众多物种的适宜生境[36]。因此耐高温、耐干旱、耐贫瘠的骆驼刺、沙拐枣等植被将是荒漠生态系统最重要的捍卫者。
本研究主要研究生物气候因素、海拔和人类活动强度对骆驼刺适生区的影响,并未考虑地下水、土壤因素、植物与环境的相互作用等方面的因素。这些因素对植物的适生区范围也具有重要的影响作用,因此今后的研究应将更多因素纳入模型,以期为骆驼刺资源的保护和开发提供更有价值的理论支持。且未来气候仅使用了BCC-CSM2-MR模式下的数据,而在CMIP6情景下,具有多种不同的气候模式,且各有偏重。今后的研究应关注不同气候模式下,物种适生区的变迁,以获得更有价值的预测结果。
4 结论
不同的环境因素对最大熵模型的预测结果具有影响。在仅考虑生物气候因素和海拔因素的情况下,骆驼刺的适生区覆盖我国西北干旱区的大部分面积。而在加入人类活动强度因素之后,骆驼刺适生区面积显著下降至71.31万km2,且呈现出破碎化状态。这种下降一方面是环境因子对模型预测结果的影响,另一方面是人为因素干扰了骆驼刺的生存,使其适生区面积显著下降。无论是否包含人为因素的影响,年降水量始终是影响骆驼刺分布的最重要因素,骆驼刺适生区的年降水量范围较窄,为18—145 mm。在未来气候情景下,骆驼刺的适生区显著扩张,主要向高海拔的青海北部、高纬度的新疆北部以及内蒙古西北部发展。骆驼刺具有耐高温、耐干旱、耐贫瘠的特点,其在气候变化条件下依然能够在西北干旱区生存,因而可将其作为防沙治沙的优良品种进行保护与开发。
猜你喜欢
汽车实用技术(2022年14期)2022-07-30
疯狂英语·初中天地(2022年2期)2022-07-07
北京航空航天大学学报(2021年4期)2021-11-24
学苑创造·A版(2021年5期)2021-06-28
电子技术与软件工程(2019年8期)2019-07-16
中学生数理化·教与学(2019年5期)2019-06-06
科学大众(中学)(2019年3期)2019-05-17
劳动保护(2019年3期)2019-05-16
小天使·一年级语数英综合(2017年3期)2017-04-25
科技知识动漫(2017年1期)2017-02-06
