
我国粳不籼恢亚种间杂交稻的研究进展与展望
2022-10-11 03:30:48施勇烽刘鑫华宇峰宋昕蔚鄂志国吴明国林建荣
中国稻米 2022年5期
施勇烽刘鑫华宇峰宋昕蔚鄂志国吴明国林建荣*
(1中国水稻研究所/水稻生物学国家重点实验室,杭州310006;2浙江省种子管理总站,杭州310018;#共同第一作者:shiyongfeng@caas.cn;*通讯作者:linjianrong@caas.cn)
粳不籼恢亚种间杂交稻是指利用粳稻不育系与偏籼型或籼粳中间型广亲和恢复系配制的一类籼粳亚种间杂交稻。籼稻和粳稻是亚洲栽培稻的两个亚种,一般籼稻茎秆偏软、分蘖力较强、谷粒细长、较耐热不耐寒,而粳稻一般株型紧凑、叶片短直、谷粒短圆、较耐肥耐寒。育种实践表明,籼粳亚种间杂种优势比传统的籼籼交、粳粳交更为明显[1]。但由于两者间遗传差异较大、亲缘关系较远,籼粳杂种一代结实率往往较低。20世纪80年代,IKEHASHI等[2]研究揭示了籼粳不亲和性的机理,提出籼粳亲和性是由一组复等位基因控制,为解决籼粳杂种F1结实率低的问题指明了方向。20世纪末,国内育种机构陆续利用“籼粳架桥”的策略培育了协优9308、两优培九等一批具有部分籼粳亚种间杂种优势的杂交稻组合[3-4]。2005年,我国第1个粳不籼恢亚种间杂交稻甬优6号在浙江省通过品种审定,2006年被农业农村部认定为超级稻品种,并在生产上推广应用。21世纪以来,浙江省的一些育种单位利用粳型不育系与籼粳中间型广亲和恢复系配组,相继选育出了一批在全国有影响力的粳不籼恢亚种间杂交稻。
本文对我国籼粳亚种间杂交稻的发展历史、粳不籼恢亚种间杂种优势利用研究现状进行了回顾与展望,以期为今后我国籼粳亚种间杂种优势利用研究和籼粳亚种间杂交稻培育提供有益的借鉴与参考。
1 籼粳亚种间杂种优势利用研究的历史
1926年,美国学者JONES首次提出水稻具有杂种优势,1930年中国科学家丁颖用早银黏与印度野生稻杂交,培育出每穗达1 400粒的巨型水稻[5]。而后,印度、日本、美国等国家的科学家也开展了水稻杂种优势利用研究,但均未能应用于生产。袁隆平先生在1964年开始开展水稻雄性不育系选育,提出通过不育系、保持系和恢复系三系配套法利用水稻杂种优势,并于1973年真正实现三系配套。
1987年,袁隆平根据水稻杂种优势的利用水平,战略性的提出杂交水稻育种分为三系法为主的品种间杂种优势利用、两系法为主的亚种间杂种优势利用和一系法远缘杂种优势利用三个发展阶段[6]。在此之前,亚种间杂种优势尤其是籼粳杂种的巨大产量潜力已经引起育种家的关注。籼粳杂交种表现较强的生长优势,具有植株高大、穗大粒多、苗期生长快、分蘖力强、茎粗抗倒、根系发达和抗逆力强等特点[7]。20世纪50年代,杨守仁系统地阐述籼粳杂交后代结实率低、性状不易稳定、籼粳稻优点能否结合及杂种优势能否保持利用等籼粳杂交育种的核心问题[8]。20世纪70年代,杨守仁将籼稻中的有利基因导入粳稻,提出了部分籼粳有利性状的稳定优势利用[9]。杨振玉等[10]通过“籼粳架桥”制恢技术培育出了具有籼稻血缘的粳型强恢复系C57,并提出了“籼粳架桥”亲和、有利基因交换、差异适度的协调发展观点[11]。
20世纪80年代,IKEHASHI等[12]研究发现,爪哇稻及籼粳中间类型品种中存在一些与籼粳杂交F1育性均正常的品种,IKEHASHI称之为广亲和品种。同时期,石明松[13]从农垦58天然雄性败育株中选育了光温敏雄性不育系农垦58S。随着水稻广亲和性的利用和两系不育系材料的创制,我国籼粳亚种间杂种优势利用开始摆脱恢保关系及籼粳杂种F1不育的制约,籼粳杂种优势利用开始从间接利用走向直接利用。经过多年努力,湖南杂交水稻研究中心采用广亲和基因与光温敏不育基因相结合的方法,成功培育带有1/4爪哇稻血缘的广亲和两系不育系培矮64S[14]。1996年我国提出了实施水稻超高产育种的设想,即采用理想株型塑造与籼粳杂种优势利用相结合的技术路线大幅提高水稻单产。江苏省农业科学院用培矮64S与9311配组培育出了两系杂交稻标志性品种两优培九,两优培九成功利用了部分亚种间杂种优势,通过理想株型与强生理功能相结合,协调了籼粳交的诸多矛盾,将优质、超高产、多抗性状聚集于一体[4]。中国水稻研究所利用籼中掺粳恢复系中恢9308配制出具有较强高产优势的组合协优9308[3]。
李振宇等[15]曾指出“粳不/籼恢”配组方式在水稻亚种间杂种优势利用中的重要性,但其存在花时不遇制种难等问题。21世纪初,浙江省部分科研机构在三系法粳不籼恢亚种间杂种优势利用研究方面取得重要突破。如宁波市农业科学研究院以特殊配合力晚粳型不育系甬粳2号A和多个强恢中籼型恢复系配组,育成了甬优6号、甬优12等多个籼粳亚种间杂交稻组合。中国水稻研究所利用早花时晚粳不育系春江12A、春江16A与多个籼粳中间型广亲和恢复系配组育成了春优58、春优84等强优势籼粳亚种间杂交稻组合。甬优系列等籼粳亚种间杂交稻组合具有长势旺盛、茎秆粗壮、丰产性好等特点,它们的大面积推广标志着我国籼粳亚种间杂种优势利用的育种实践已经进入到一个新的阶段。
2 粳不籼恢亚种间杂种优势利用研究的现状
2.1 籼粳亚种间杂种优势利用理论研究进展
1986年,IKEHASHI和ARAKI[2]通过不同生态型水稻品种间的杂交研究,发现了位于第6染色体上控制籼粳亚种间杂种不育的S5位点。该位点在籼稻、粳稻和广亲和品种中的等位基因分别是S5i、S5j和S5n,籼粳交杂种F1由于携带S5j的雌配子败育而产生半不育小穗,而广亲和品种与籼粳稻杂交均产生可育小穗[16]。通过图位克隆和功能研究发现S5位点包含ORF3、ORF4和ORF5等3个开放阅读框,分别编码热激蛋白Hsp70、膜蛋白和天冬氨酰蛋白酶,形成killer-protector系统[17]。S5i等位基因有ORF3+/ORF4+/ORF5+和ORF3+/ORF4-/ORF5+两种类型,S5j等位基因为ORF3-/ORF4+/ORF5-,而S5n等位基因为ORF3+/ORF4+/ORF5n,ORF5+和ORF5-仅在第2外显子上有2个SNP的差异,ORF5n则由于缺失了N端信号肽不能定位到细胞壁上而丧失了功能。在S5i/S5j的杂合子中,ORF5+与ORF4+结合杀死雌配子,而带有保护基因ORF3+的雌配子则免于败育[17-18]。
除了S5位点外,还有一些控制籼粳亚种杂交F1代育性的基因位点报道,如Sa[19]、Sb[20]、Sc[21]、Sd[22]和Se[23],均为控制雄配子育性的基因座位。目前粳不籼恢杂种优势利用过程,尤其是籼粳中间型恢复系选育中主要利用了S5n基因型的广亲和性。杨杰等[24]根据S5位点的序列差异,设计InDel标记S5136,筛选到13份具有广亲和基因S5n的材料,该InDel标记可用于广亲和基因资源筛选和分子标记辅助选择育种。林建荣等[25]通过S5n位点筛选获得了带有广亲和基因的籼粳中间型恢复系CH58、C84和C927,这些恢复系与粳型和籼型广亲和测验种测交F1的平均结实率均超过80.0%,而对照粳稻恢复系与籼型测验种的平均结实率仅为16.9%。
在利用广亲和材料的基础上,袁隆平[26]曾提出“矮中求高、远中求近、显超兼顾、穗求中大、高粒叶比、以饱攻饱、爪中求质和生态适应”等8条亚种间杂交组合选育策略。sd-1半矮秆基因的广泛利用基本解决了亚种间杂交品种植株过高的问题,而发掘新矮秆基因并应用于亚种间杂交育种,可为解决籼粳杂种F1株高偏高寻到新的解决途径。如显性半矮秆基因Dwarf53,可使杂种F1植株的株高基本介于双亲之间,该基因参与独脚金内酯的生理生化途径[27]。目前生产中利用的粳型不育系多为半矮秆株型,如甬粳2号A、春江16A。籼粳中间型恢复系与粳稻保持系相比,在株高、剑叶长、每穗粒数等主要农艺性状方面存在较明显的差异,这也有利于通过性状互补配置出强优势的粳不籼恢亚种间杂交稻组合[25]。袁隆平[26]认为,在不倒伏的前提下,适当增加杂交稻株高,使源库关系更加平衡,可提高亚种杂交组合产量。
籼粳亚种间杂种F1抽穗期具有明显的超亲优势,成为籼粳亚种间杂种优势利用的主要障碍。近年来,浙江省籼粳亚种间杂交稻育种实践表明,“粳不籼恢”这一配组方式的杂种优势非常明显,但也存在生育期偏长的问题。研究表明,籼粳杂交F1的抽穗期主要取决于亲本的一般配合力大小,与亲本自身熟期基本一致[28]。双亲是否带有互补的显性感光基因是决定籼粳交F1抽穗期是否超亲晚熟的主要因素[29],选用无感光性,或感光性基因相同,或感光性基因非互补的籼粳品种作为亲本,可能是获得籼粳交杂种熟期适中的关键[27]。利用感温型中粳不育系春江99A、春江23A与籼粳中间型广亲和恢复系配组,其F1的播始历期明显缩短[30],表明通过筛选均带有隐性感光抑制基因的不育系和恢复系,可避免籼粳交F1生育期超亲晚熟。
2.2 粳不籼恢亚种间杂交稻品种选育情况
2005年,以粳型不育系甬粳2号A为母本,籼型恢复系K4806为父本杂交育成的籼粳杂交稻组合甬优6号通过浙江省审定,粳不籼恢亚种间杂交稻组合开始在生产上大面积推广和应用。根据国家水稻数据中心(http://www.ricedata.cn)的数据,对符合粳不籼恢亚种间类型的杂交组合进行分类统计,2005—2021年我国通过省级以上审定的粳不籼恢亚种间杂交水稻品种(组合)共217个次,其中有37个品种至少通过2个省级以上审定。217个次审定品种区试平均产量为664.8 kg/667 m2,平均比对照品种增产9.6%,增产极其显著。2005—2012年,我国粳不籼恢亚种间杂交组合年审定数均未超过5个次,2017—2021年我国粳不籼恢亚种间杂交组合审定呈爆发式发展,年均审定数约为31个,其中2021年审定数达到59个次(图1)。

图1 2005—2021年通过省级以上审定的粳不籼恢亚种间杂交组合数
2005—2017年浙江省审定的粳不籼恢杂交水稻品种达69个次,国家审定的为47个次,江苏省审定的为21个次,审定数超过10个次以上的还有湖北、广西、江西、安徽和福建。2017—2021年国家和浙江省审定的粳不籼恢杂交稻数量仍为最多,但广西、江苏等省份审定的粳不籼恢杂交稻品种数量呈逐年增多趋势(图2)。2019年浙江省没有籼粳杂交稻获得审定,这可能与2018年浙江省提高了水稻新品种审定标准,且将单季籼粳杂交稻组的对照品种改为综合性状突出的甬优1540有关。此外,从配组方式来看,我国2005—2021年通过省级以上品种审定的217个次粳不籼恢亚种间杂交稻中,两系法组合只有2个次,占比仅0.92%,绝大多数为三系法籼粳亚种间杂交稻组合,说明在两系法籼粳亚种间杂交稻组合选育方面还有较大的努力和发展空间。
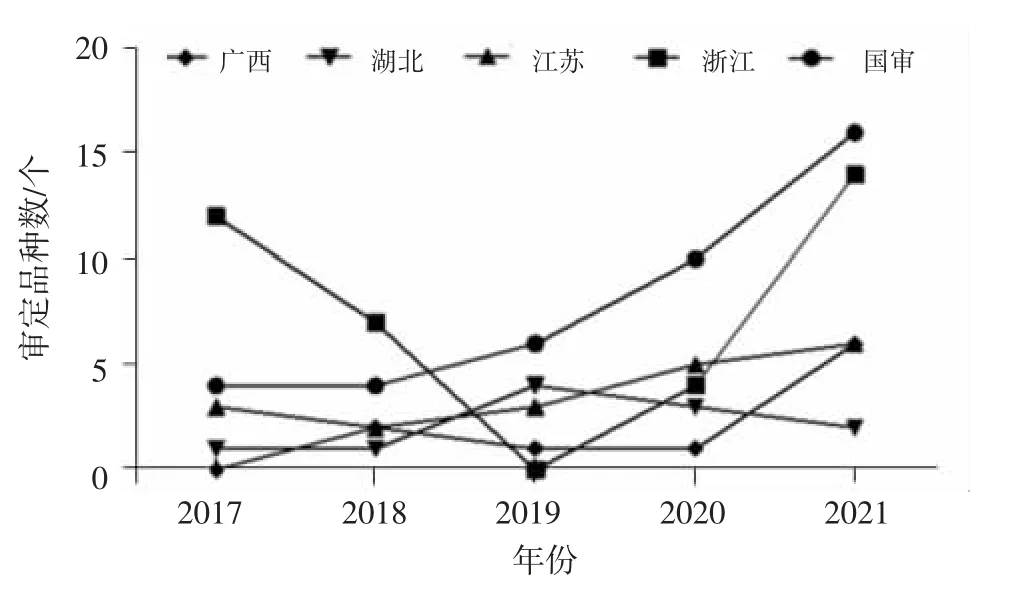
图2 2017—2021年主要区试通道审定粳不籼恢亚种间杂交稻组合情况
根据审定品种米质数据统计,2014—2021年达到NY/T593-2013《食用稻品种品质》优质稻标准的粳不籼恢杂交稻品种在增多,2020年和2021年分别有14和37个次达到优质稻标准,分别占审定粳不籼恢杂交稻品种数的48.3%和62.7%。按此标准,2014—2021通过审定的粳不籼恢杂交稻品种整精米率优质达标率为86.6%,垩白度优质达标率为72.4%,碱消值优质达标率为78.0%。
对浙江省2014—2021年通过审定的粳不籼恢杂交稻品种中整精米率、垩白度和碱消值3个指标变化(2019年由于没有品种通过审定,无法统计)进行分析,整精米率指标均值2014年为70.1%,2021年为65.8%,呈下降趋势(图3A),这可能与近年来长粒型籼粳杂交稻品种审定逐年增多有关;品种间垩白度值差异大,变异系数也较大,2014年、2015年和2020年变异系数均超过60.0%,但垩白度均值呈下降趋势,说明垩白度这一单项外观品质明显改善(图4 B);碱消值年均值在6.1~6.9之间波动,其中2018年的均值最大达6.9(图4C)。在米质单项指标中,整精米率、垩白度和碱消值被认为是制约浙江省籼粳杂交稻品质的主要因素[31],目前这3项指标总体趋于稳定向好,优质率稳中有升,是这两年浙江省优质粳不籼恢杂交稻品种数量增多的主要原因。

图3 2014—2021年浙江省审定粳不籼恢亚种间杂交稻品种整精米率、垩白度和碱消值变化趋势
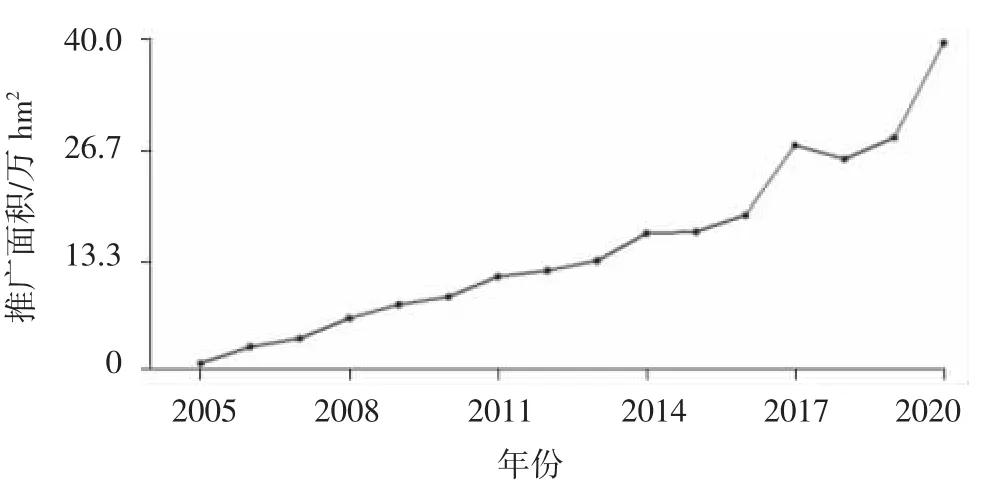
图4 2005—2020年粳不籼恢亚种间杂交稻推广面积
2.3 粳不籼恢亚种间杂交稻品种推广情况
对我国2005—2020年年推广面积在0.67万hm2(10万亩)以上的粳不籼恢亚种间杂交稻品种推广面积进行统计,粳不籼恢亚种间杂交稻累计推广约239.9万hm2,并呈逐年扩大趋势;2009年粳不籼恢亚种间杂交稻组合推广面积已超6.7万hm2,2014年推广面积超过13.4万hm2,2017年推广面积超过26.7万hm2,2020年推广面积达到39.7万hm2(图4)。
对2005—2020各省粳不籼恢亚种间杂交稻品种累计推广面积进行统计,累计推广面积超过3.4万hm2的仅浙江、福建、江西、江苏和广西等5个省份。其中,浙江省累计推广面积最大,达147.6万hm2,推广面积明显呈逐年快速增长的趋势;福建省累计推广44.2万hm2,呈逐年缓慢增长态势;江西省累计推广35.6万hm2,2017年 其 推 广 面 积 超 过6.7万hm2,2018年 和2019年则出现明显下滑,2020年重拾升势,推广面积达8.3万hm2(图5)。

图5 2005—2020年粳不籼恢亚种间杂交稻在浙江、福建和江西的推广情况
对2005—2020年全国粳不籼恢亚种间杂交稻品种推广情况进行统计,累计推广面积超过6.7万hm2的品种有甬优9号、甬优1540、甬优6号、甬优538、甬优1538和春优59。其中,甬优9号累计推广88.7万hm2,甬优1540累计推广36.5万hm2,甬优6号累计推广28.9万hm2,甬优538累计推广22.2万hm2,甬优1538累计推广15.8万hm2,春优59累计推广10.7万hm2。推广面积较大的品种还有甬优2640、春优84、甬优15、浙优18和甬优4949等。
截至2021年,经农业农村部确认,冠名超级稻的水稻品种135个,其中粳不籼恢亚种间杂交品种9个,占比约为6.7%。根据这些品种在区试中的表现,发现粳不籼恢亚种间杂交稻品种具有株型较高大、穗型特大、全生育期较长、结实率较高和超高产特性明显等特征(表1)。甬优系列和春优系列粳不籼恢亚种间杂交稻在浙江省水稻生产上屡创高产记录,超级稻品种甬优12于2012年在浙江省宁波市鄞州区的百亩示范方平均单产达到963.7 kg/667 m2,最高田块单产1 014.3 kg/667 m2,分别创下当年我国水稻百亩示范方和我国水稻主产区高产记录。2015年春优927在浙江省宁波市宁海县的百亩示范方取得平均单产1 015.5 kg/667 m2成绩,创造了浙江省水稻百亩示范片产量纪录。
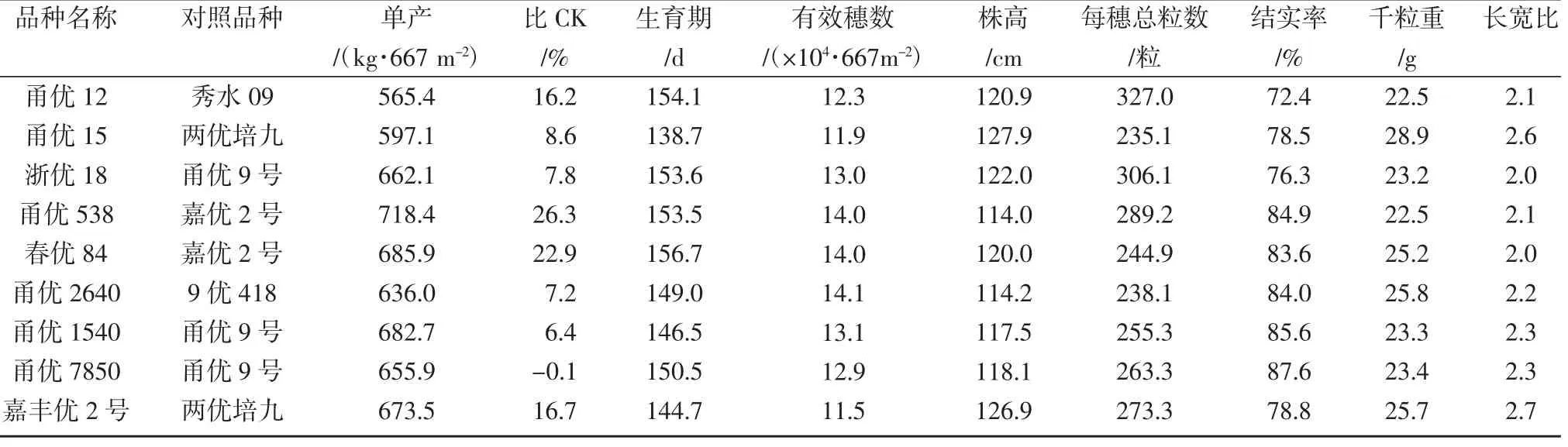
表1 粳不籼恢超级稻品种的区试表现
2.4 粳不籼恢亚种间杂交稻配套栽培技术研究
粳不籼恢亚种间杂交稻作为籼粳杂交稻的最主要类型,其配套栽培技术研究主要围绕超高产栽培展开。张洪程等[32]提出,水稻超高产栽培的根本在于“强支撑、扩库容、促充实”,具体栽培模式为“精苗稳前、控蘖优中、大穗强后”,要求移栽时基本保持叶蘖同伸,有效分蘖临界叶龄期准时够苗,及早控制无效分蘖、育壮秆、攻大穗,增强后期物质生产力。粳不籼恢杂交稻品种穗型大,穗粒数多,非常适合走强壮茎秆支撑大穗的扩库途径。韦还和等[33]通过对甬优系列籼粳杂交稻的生理形态特征进行分析后进一步提出了“稀植化栽插、合理施用氮肥”等配套高产栽培关键措施。
此外,粳不籼恢亚种间杂交稻稻米品质的形成与播期、氮肥用量及其运筹等措施密切相关。徐栋等[34]研究发现,灌浆结实期温度与籼粳杂交稻的加工品质、外观品质和蒸煮食味品质负相关,而通过合理调整播期或栽培管理,使灌浆结实期处于相对较低的温度有利于提高籼粳杂交稻的外观品质和蒸煮食味品质。在轻简化、机械化栽培模式方面,籼粳杂交稻机插技术研究表明,精准条播机插能提高籼粳杂交稻植株均匀度,促进干物质积累和氮素吸收,有效减少氮肥减施下的产量损失[35]。
3 粳不籼恢亚种间杂种优势利用研究的建议与展望
3.1 提高育种效率,挖掘粳不籼恢亚种间杂交稻高产潜力
我国人口众多,解决好“吃饭”问题,始终是治国理政的头等大事。而在耕地资源有限的情况下,提高水稻单产水平就显得尤为重要。粳不籼恢亚种间杂交稻与品种间杂交稻相比,其营养生长旺盛、生物学产量高、抗倒性强、丰产性更好。2005—2017年浙江省通过品种审定的44个次粳不籼恢亚种间杂交稻,区试产量平均比对照品种增产13.9%。而2018—2021年浙江省以超级稻甬优1540作为对照品种后,通过审定的粳不籼恢杂交稻为13个次,平均产量比对照品种减产0.2%,表明目前粳不籼恢亚种间杂交稻品种的产量可能处在了高位瓶颈期。WANG等[36]利用甬优4953、甬优4949和甬优4149构建3个F2群体,通过杂种测序和基因型解析发现,带有株型、产量相关的优良等位基因(sd1、NAL1、Ghd7和Ghd8)的约25%的杂种基因组是纯合的,而其他一些与产量相关的基因(Gn1a、IPA1、DTH2、Hd1和Tb1)则为杂合态,且大部分表现为正显性效应,正是这些累加正显性效应和杂种可育共同解释了甬优粳不籼恢杂交稻组合的高产表现。这也表明,利用现有粳不籼恢杂交稻亲本资源,通过与产量相关基因的单倍性分析,结合S5n、Sd、Sc等育性基因检测,预测杂种优势特性,或可快速筛选到具有高产潜力的杂交稻组合,从而进一步发挥粳不籼恢亚种间杂交稻产量潜力,提高育种效率。
3.2 优化亲本特性,提高粳不籼恢杂交稻制种产量
受到粳稻不育系柱头不外露或外露率较低、粳稻不育系与籼粳中间型广亲和恢复系花时差明显等多方面影响,与杂交籼稻相比,目前生产上利用的粳不籼恢亚种间杂交稻组合制种产量总体仍偏低,种子成本较高,影响了其在生产上大面积推广应用。显然,改造粳稻不育系柱头外露率特性,提早粳稻不育系花时或延后籼粳中间型广亲和恢复系花时,缩短两者开花时差可明显提高制种产量。育种实践中,将早花时高柱头外露率籼稻资源作为供体,通过回交将目标性状转入粳稻保持系,可转育成具有高柱头外露率的早花时粳型不育系。宋昕蔚等[37]将培矮64S的高柱头外露率、早花时特性定向导入普通粳稻,选育出农艺性状好、柱头外露率高、花时早的粳稻保持系材料春江95B,进而转育出高柱头外露率的早花时粳稻不育系春江95A。在恢复系选育中则要选择籼稻成分适当、花时相对较迟、花药大、花粉量足、花粉抗风性好的籼粳中间型广亲和恢复系,以缩小父母本花时差、提高异交结实率[30]。
两系法粳不籼恢亚种间杂交稻摆脱了恢保关系限制,具有配组自由、种子繁育程序更简单、成本更低等优势,但目前通过品种审定的两系法粳不籼恢亚种间杂交稻组合还很少,主要是缺乏育性转换起点温度低、柱头外露率高、花时集中的粳型两系不育系。通过聚合光温敏核不育基因pms1、pms3和tms5获得育性稳定的低起点温度的粳型不育系,并进一步改良不育系柱头外露率、花时集中度等特性,通过优良型两系不育系与籼粳中间型广亲和恢复系配组,选育杂种优势强、制种产量高的两系法籼粳亚种间杂交稻,可以解决目前粳不籼恢亚种间杂交稻制种产量偏低的问题。
除粳不籼恢亚种间杂交稻以外,利用高柱头外露率的籼型、掺粳偏籼型三系或两系不育系与现有的籼粳中间型广亲和恢复系配组,选育制种产量高的三系法或两系法“籼不粳恢”亚种间杂交稻,可能会比三系法、两系法“粳不籼恢”亚种间杂交稻取得更好的效果[30]。如中国水稻研究所利用超级稻春优84的父本C84与籼型三系不育系五丰A配组,选育出五优84通过广东省品种审定;利用籼型三系不育系华浙2A与籼粳中间型广亲和恢复系钰禾配组,选育出华优钰禾通过广西、福建品种审定;利用籼型两系不育系广湘24S与籼粳中间型广亲和恢复系R998配组,选育出湘两优998通过国家品种审定等。
2019年,WANG等[38]利用基因编辑技术在粳不籼恢杂交稻春优84中同时敲除4个基因(OsPAIR1、Os-REC8、OsOSD1和OsMTL),将无融合生殖特性引入到杂交稻中,获得了杂种优势固定的种子。尽管该技术单倍体诱导率仍较低,种子育性也明显下降,但相信经过技术的不断改进,有望突破杂交制种限制而在生产上直接应用。
3.3 协调综合性状,扩大粳不籼恢亚种间杂交稻适应区域
随着我国经济的发展和人们生活水平的提高,稻米消费从以往追求数量向追求质量转变。稻米品质除了垩白、直链淀粉含量、碱消值等性状以外,受各地稻农品种种植习惯和居民稻米消费习惯的影响,稻米长宽比改良将是今后我国籼粳亚种间杂交稻发展必须考虑的一个重要问题。稻米长宽比是一个直接影响稻米外观品质的主要性状,也间接影响到稻米的整精米率。从近年来浙江省籼粳杂交稻区试及审定的发展趋势来看,粳不籼恢亚种间杂交稻的长宽比性状两极化改良值得关注。从2021年开始,浙江省单季籼粳杂交稻区试以稻米粒型为标准,分设团粒组和长粒组,明确要求品种粒长≥6.0 mm或长宽比≥2.5的品种可参加长粒组,其余参加团粒组。长宽比2.0以下的团粒型籼粳杂交稻和长宽比2.5甚至3.0以上的长粒型籼粳杂交稻将是今后籼粳杂交稻改良的发展方向。籼粳杂交稻粒型以及株叶形态的定向改良,即团粒型籼粳杂交稻面向传统团粒粳稻区,长粒型籼粳杂交稻面向传统长粒籼稻区,有利于籼粳亚种间杂交稻不断扩大适应种植区域,分类满足我国水稻主产区居民对稻米品质的更高要求。
由稻绿核菌引起的稻曲病是水稻生产面临的主要病害之一,往往造成水稻减产,并影响稻米品质,威胁食用安全。稻曲病的发生与流行与品种特性、气候条件及栽培措施密切相关,粳不籼恢杂交稻由于其大穗多粒的穗部特征而更易感稻曲病。目前尚未发现具有抗稻曲病的粳不籼恢杂交稻品种,化学防治仍是当前防治稻曲病最有效的方法,但从稻田绿色生产和节本增效等角度考虑,提高品种抗性是最经济、安全的方法。稻曲病侵染机制及与水稻互作机理的研究表明,稻曲菌分泌枯草杆菌蛋白酶UvPr1a靶向降解水稻免疫调控因子OsSGT1从而抑制寄主抗性,而过表达OsSGT1可显著提高稻曲病的抗性[39],这为进一步遗传改良稻曲病抗性提供了方向。
2021年国家农作物品种审定委员会对国家级稻品种审定标准进行了修订,修订后的审定标准不但提高了品种产量指标,与已知品种DNA指纹差异位点及抗性等指标也明显提高。因此,未来粳不籼恢亚种间杂交稻品种要集合高产、优质、多抗、广适等优良特性,才能在稻米生产和消费领域占据主导优势。中国水稻研究所2022年有2个“T27”系列优质强优势籼粳杂交稻春优167和春优83通过浙江省品种审定,春优83区试比对照甬优1540增产6.8%、部标2级优质米,春优167区试对照甬优1540增产3.3%、部标1级优质米,这2个品种株叶形态优良,后期转色好,综合性状突出。未来要实现综合性状优异的粳不籼恢杂交稻品种的突破,仍需加大粳不籼恢杂交稻杂种优势机理、有利基因挖掘利用、亲本资源创制等基础领域的研究工作。此外,传统育种技术对籼粳杂交稻香味、粒型、碱消值、抗病性等性状进行遗传改良往往耗时费力、效率低下,随着基因编辑技术的兴起和安全评价管理体系的进一步规范,基因编辑技术或将带动籼粳亚种间杂交稻育种进入精准调控时代。
猜你喜欢
Natural Products and Bioprospecting(2023年6期)2023-12-29 13:42:30
华中农业大学学报(自然科学版)(2022年1期)2022-11-23 08:48:55
广东农业科学(2022年9期)2022-11-21 07:39:30
作物学报(2022年2期)2022-11-06 12:00:52
热带亚热带植物学报(2022年4期)2022-08-18 00:36:20
蔬菜(2021年7期)2021-11-27 03:45:25
——和田盘羊
野生动物学报(2021年4期)2021-11-14 06:13:24
山西农经(2016年5期)2016-02-28 14:24:37
现代农业科技(2015年13期)2015-09-06 06:17:34
水生生物学报(2015年1期)2015-02-28 16:01:28
