
有氧暴露下全株玉米青贮饲料品质及微生物动态变化
2022-10-11 08:46:38王昌洲叶兴腾韩富荣梁龙飞孙文涛
草业科学 2022年8期
张 莓,王昌洲,叶兴腾,韩富荣,梁龙飞,孙文涛,郝 俊
(贵州大学动物科学学院草业科学系, 贵州 贵阳 550025)
青贮饲料是在厌氧条件下由乳酸菌经一定时间的发酵,将新鲜饲草制成一种颜色黄绿、气味酸香、质地柔软、适口性强、消化率高,能作为反刍动物在冬春季摄食的优质粗饲料[1],已经成为调节饲草料供应不平衡的重要手段之一[2]。青贮饲料原材料的品质对其青贮的品质具重大影响,而研究最多的作物就是青贮玉米(Zea mays)。全株青贮玉米具有适口性好、易消化、营养丰富、耐贮藏、青贮成功率及青贮品质较高等特点而被广泛使用[3]。近些年,为了积极响应“粮改饲”政策,贵州省采取以养带种方式推动种植业结构调整,扩大青贮玉米种植面积来增加收贮量,制成的青贮饲草料由草食家畜就地转化,进而促进草地畜牧业的发展。青贮饲料开窖后会充分接触到氧气,好氧微生物大量繁殖,致使pH 上升,青贮饲料发生有氧腐败,饲喂腐败变质的青贮饲料会给养殖业带来潜在威胁[4]。有氧腐败的青贮饲料会产生大量的霉菌毒素,一旦被反刍动物长期大量摄食,不仅会抑制反刍动物的免疫功能,使其产能降低甚至死亡,严重会影响畜牧业的健康发展[5],而且腐败青贮料中产生的霉菌毒素及其代谢产物还会通过肉类和乳制品传递给人类,最终对人类健康产生威胁[6]。
一直以来,青贮饲料的有氧稳定性都是国内外相关领域的研究热点和重点。大量研究证明,青贮饲料有氧稳定性的高低与青贮品质的好坏密切相关,因此可以通过影响青贮饲料的发酵品质以提高有氧稳定性,进而提高其饲用率和经济效益。经研究发现,青贮添加剂的适当添加可以降低青贮过程中营养成分的流失,改善发酵品质,提高有氧稳定性,从而改善动物的采食量与消化率[7-9]。此外,青贮饲料的刈割期、青贮时长和青贮密度等因素也会影响其青贮品质。王旭哲等[10]研究表明,紧实度的高低会间接影响青贮饲料的有氧稳定性及微生物群落组成,尤其是对青贮开窖后发酵品质的影响较大,且达到青贮品质和有氧稳定性最好的紧实度为600 kg·m-3。微生物是影响青贮玉米饲料青贮发酵品质的关键因素[11]。经研究发现,全株玉米原料表面附着的微生物主要有农杆菌属(Agrobacterium)、克雷伯菌属(Klebsiella)和乳杆菌属(Lactobacillus)等,青贮后则以乳杆菌属、魏斯氏菌属(Weissella)、醋酸杆菌属(Acetobacter)和假单胞菌属(Pseudomonas)等为主[12]。胡宗福等[13]研究发现,全株玉米青贮40 d 开袋暴露空气后,基于门水平的主要优势菌群为厚壁菌门(Firmicutes)和变形菌门(Proteobacteria)。基于属水平的主要优势菌群为乳杆菌属和芽孢乳杆菌属(Sporolactobacillus)。此外,青贮饲料青贮暴露于空气后,其细菌数量变化整体上呈现出乳酸菌数量下降,酵母菌、霉菌和其他好氧腐败菌数量上升的规律[14-16]。因此,为了降低饲喂青贮腐败饲料对反刍动物健康造成的不利影响,部分学者对青贮饲料产生霉菌毒素的原因、毒性作用以及降解方法开展了相关研究[17-19]。调制高品质青贮饲料的关键是具备一个由微生物良好介导的发酵环境,同时还与青贮原料特性及原料附着微生物密切相关,已有大量研究表明种植青贮原料地区的温度、降水和海拔等环境因子与青贮饲料发酵的代谢产物对青贮饲料微生物菌群的结构组成均会产生影响[20-22]。Gharechahi 等[23]利用3 个不同区域的杂交玉米进行青贮,研究发现青贮原料附着微生物的变化与原料生长的环境因子具有相关性,生长在温度、湿度及降水量较少地区的全株玉米原料,其附着细菌的多样性相对较低。Guan 等[24]对四川、重庆和贵州等5 个地区的青贮玉米饲料进行分析发现,影响原料表面附生细菌群落变化的主要因素为湿度和降水,而温度是影响青贮发酵过程中微生物群落变化的主要因素。综上,不同取样地环境因子会影响青贮饲料原料表面和发酵过程中的微生物多样性,但是否对有氧暴露期间的微生物群落结构存在直接影响缺乏深入研究。
本研究以贵州省关岭和威宁两地区的全株玉米为对象,对全株玉米青贮饲料有氧暴露后的营养品质、发酵品质及微生物菌群动态变化进行全面分析与比较,并为了保证其安全饲用,确定两组全株玉米青贮饲料的最佳或安全有氧暴露天数。
1 材料与方法
1.1 试验材料
试验材料来源于贵州关岭县沙营镇养牛村和威宁县五里岗街道骑龙村的全株玉米,其品种均为‘青丰4 号’,分别于蜡熟期刈割,将两地区全株玉米的地上部分(包括茎、叶、果实)均切短至1~2 cm,分别取一部分原料混匀分装于4 个50 mL 的灭菌离心管后,保存于-80 ℃超低温冰箱中,用于检测全株玉米原料附着的细菌,共8 个样品。分别将两地区的另一部分原料调节水分(65%~75%) 后,将其装入双层聚乙烯青贮袋(28 cm × 40 cm),每袋装料1 000 g 左右,每采样点装10 袋,真空密封,室温条件下避光贮藏45 d。本试验采样点的采样时间及采样点气候条件如表1 所列。
1.2 试验设计
发酵45 d 后,将各采样点的10 袋试验材料分别装入2 L 的敞口聚乙烯塑料桶中(共20 桶),用纱布覆盖桶口,保证空气自由进入的同时,防止原料被污染或其水分散失,置于室温条件下(日平均气温19.5 ℃)保存[10]。分别将多通道温度记录仪的探头置于原料中心,并分别放置6 个探头于环境中,用于环境温度的监测,每间隔30 min 记录一次温度,当样品温度高于环境温度2 ℃时,认为发酵饲料有氧不稳定,并记录时间[25]。
各取样点随即选取4 桶已发酵好的全株玉米青贮饲料,分别取有氧暴露0 (开袋时)、2、5 d 的青贮饲料样品约200 g 置于信封袋,于105 ℃鼓风干燥箱中杀青20 min,然后在烘箱中用65 ℃烘干48 h,用于营养成分测定。另分别取0、2 和5 d 的样品约100 g,一部分装入50 mL 灭菌离心管中速冻后置于-80 ℃超低温冰箱保存用于微生物检测;另一部分制成浸提液,测定pH 及发酵品质。其中,微生物检测设4 次重复,营养成分、发酵品质及有氧稳定性测定各设3 次重复。
1.3 测定指标与方法
1.3.1 营养成分
测定指标为干物质(dry matter, DM)、粗蛋白(crude protein, CP)、可溶性碳水化合物(water soluble carbohydrate, WSC)、淀粉(starch, ST)、中性洗涤纤维(neutral detergent fiber, NDF)和酸性洗涤纤维(acid detergent fiber, ADF)。其中DM、CP、NDF 和ADF含量采用张丽英[26]的方法测定,WSC 含量采用蒽酮-硫酸比色法测定[27],ST 含量采用酸水解-蒽酮比色法测定[28]。
1.3.2 发酵品质
浸提液制备:分别取各采样点0、2 和5 d 的待分析样品10 g,分别倒入90 mL 蒸馏水,均匀搅拌,于4 ℃下浸提24 h 后过滤得浸提液。立即测定浸提液的pH 后,置于-20 ℃条件下保存备用,用于乳酸(lactic acid, LA)、乙酸(acetic acid, AA)、丙酸(propionic acid, PA)、丁酸(butyric acid, BA)和氨态氮(ammoniacal nitrogen, AN)的测定。pH 采用上海佑科PH-3C 酸度计测定;AN 含量采用苯酚-次氯酸钠比色法测定[29];LA、AA、PA 和BA 含量采用高效液相色谱法测定[30]。
1.3.3 微生物多样性
按照DNA 提取试剂盒HiPure Stool DNA Kits 说明书提取样品的DNA,采用紫外分光光度计对提取到的样品DNA 进行浓度和纯度检测,并采用0.8%琼脂糖凝胶电泳进行DNA 完整性检测。对细菌16S rDNA 基因V5~V7 区进行序列扩增,引物序列参照梁龙飞[31],引物序列为799F (AACMGGATT AGATACCCKG)及1193R (ACGTCATCCCCACCTT CC)。纯化扩增产物使用AMPure XP Beads,定量使用ABI StepOnePlus Real-Time PCR System (Life Technologies,美国),测序根据Hiseq2500 的PE250模式pooling 上机。
生物信息学和数据分析参照胡宗福等[13]的方法将测序后的原始数据过滤,再进行序列的拼接、过滤和去嵌合体等处理。利用OTU、Chao、ACE、Simpson、Shannon 指数来度量细菌α 多样性,基于OTU 分析结果列出两组全株玉米表面附着微生物及有氧暴露后的微生物菌群的门、属类丰度均值排名top10 的物种堆叠图。
1.4 数据分析
所测数据均用Excel 2019 初步整理后用SPSS 26.0 软件进行双因素方差分析、多重比较(Duncan法)及独立样本t检验,测定结果用“平均值 ± 标准误”表示;采用OMICSMART 微生物多样性分析平台与Adobe Illustrator 2021 制图。“P< 0.05”表示差异显著。利用Chao 丰富度估计量和ACE 计算菌群丰度;利用Shannon-Wiener 多样性指数和Simpson多样性指数计算菌群多样性。计算方式如下:
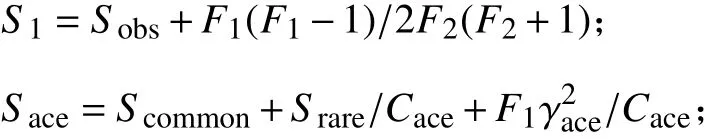

式中:S1为Chao;F1为样本中数量只为1 的物种数;F2为样本中数量只为2 的物种数;Sobs为测序分析得到的物种数;Sace为ACE 指数;Scommon为样本中数量超过10 的物种数;Srare为样本中数量不超过10 的 物 种 数;Cace为Srare中 非singleton 的 比 例;为变异系数;H为Shannon;Pi为第i个物种的个体数占总个体数的比例;Ds为Simpson。
2 结果与分析
2.1 全株玉米原料的营养成分
如表2 所列,两组全株玉米原料的各营养成分间差异较大。其中,关岭组的DM 和ST 含量显著低于威宁组(P< 0.05),分别少了11.44%和7.16%;关岭组的CP、WSC、NDF 和ADF 含量显著高于威宁组(P< 0.05),分别多了7.78%、9.58%、5.61%和2.62%。

表2 全株玉米原料的营养成分Table 2 Nutrient composition of the whole-maize raw material
2.2 有氧暴露期间全株玉米青贮饲料营养物质动态变化
随着有氧暴露天数的延长,两组有氧暴露5 d的DM、CP、WSC、ST、NDF 和ADF 含量较0 d 均显著 降 低(P< 0.05) (表3), 关 岭 组 分 别 降 低了13.86%、4.07%、46.90%、24.46%、14.05% 和24.64%,威宁组分别降低了15.39%、6.33%、39.91%、29.01%、7.16%和7.32%。两组有氧暴露2 d 的DM、WSC、ST、NDF、ADF 和威宁组有氧暴露2 d 的CP 的含量较0 和5 d 均差异显著(P< 0.05),关岭组有氧暴露2 d的CP 含量较0 和5 d 均无显著差异(P> 0.05)。关岭组在有氧暴露期间的DM 和ST 含量均显著低于威宁组(P< 0.05),CP 和NDF 含量均显著高于威宁组。两组的WSC 含量在0 和5 d 无显著差异,而2 d时关岭组的WSC 显著低于威宁组。关岭组的ADF含量在0 d 时显著高于威宁组,在2 和5 d 时显著低于威宁组。

表3 青贮玉米饲料有氧暴露后的营养成分Table 3 Nutrient composition of the silage maize under aerobic exposure
2.3 有氧暴露期间全株玉米青贮饲料有氧稳定性及发酵品质动态变化
经测定,关岭组和威宁组全株玉米有氧稳定性分别为130.50 和61.83 h。随有氧暴露天数的延长,两组有氧暴露5 d 的pH、PA 和AN/TN 的含量较0 d 均显著增加(P< 0.05) (表4),关岭组分别增加了13.99%、22.22% 和20.86%, 威 宁 组 分 别 增 加了24.43%、12.00% 和20.38%;两 组 有 氧 暴 露5 d 的LA 和AA 的含量较0 d 均显著降低(P< 0.05),关岭组分别降低了44.11% 和33.33%,威宁组分别降低了62.15%和36.73%。两组有氧暴露2 d 的LA、AA、PA、AN/TN 和威宁组有氧暴露2 d 的pH 的含量较0 和5 d 均差异显著,关岭组有氧暴露2 d 的pH 较0 d 无显著差异(P> 0.05),显著低于5 d 的pH。关岭组的AA、PA 和AN/TN 含量在有氧暴露期间均显著高于威宁组。关岭组的pH 在0 和2 d 时显著高于威宁组,在5 d 时同威宁组无显著差异。关岭组的LA 含量在0 和2 d 时显著低于威宁组,在5 d 时两组无显著差异。两组均未检测到BA 的存在。

表4 青贮玉米饲料有氧暴露后的发酵品质Table 4 Fermentation quality of the silage maize under aerobic exposure
2.4 有氧暴露期间玉米青贮饲料微生物动态变化
2.4.1 细菌α 多样性
有氧暴露5 d 与0 d 相比,关岭组的Chao、ACE、Shannon-Wiener、Simpson 和两组的OTU 的数量均显著降低(P< 0.05) (表5),而威宁组的Chao、ACE、Shannon-Wiener 和Simpson 数量无显著变化(P> 0.05)。关 岭 组 有 氧 暴 露2 d 的OTU、Chao、ACE、Shannon-Wiener 和Simpson 数量较0 d 无显著差异(P> 0.05),而较5 d 差异显著。威宁组有氧暴露2 d的OTU、Chao、ACE、Shannon-Wiener和Simpson数量较0 和5 d 均差异显著。两组间比较,有氧暴露期间的Chao 和ACE 数量均无显著差异。关岭组在0 d 时的OTU、Shannon-Wiener 和Simpson 数量均显著高于威宁组,2 和5 d 时的OTU、Shannon-Wiener和Simpson 数量同威宁组无显著差异。
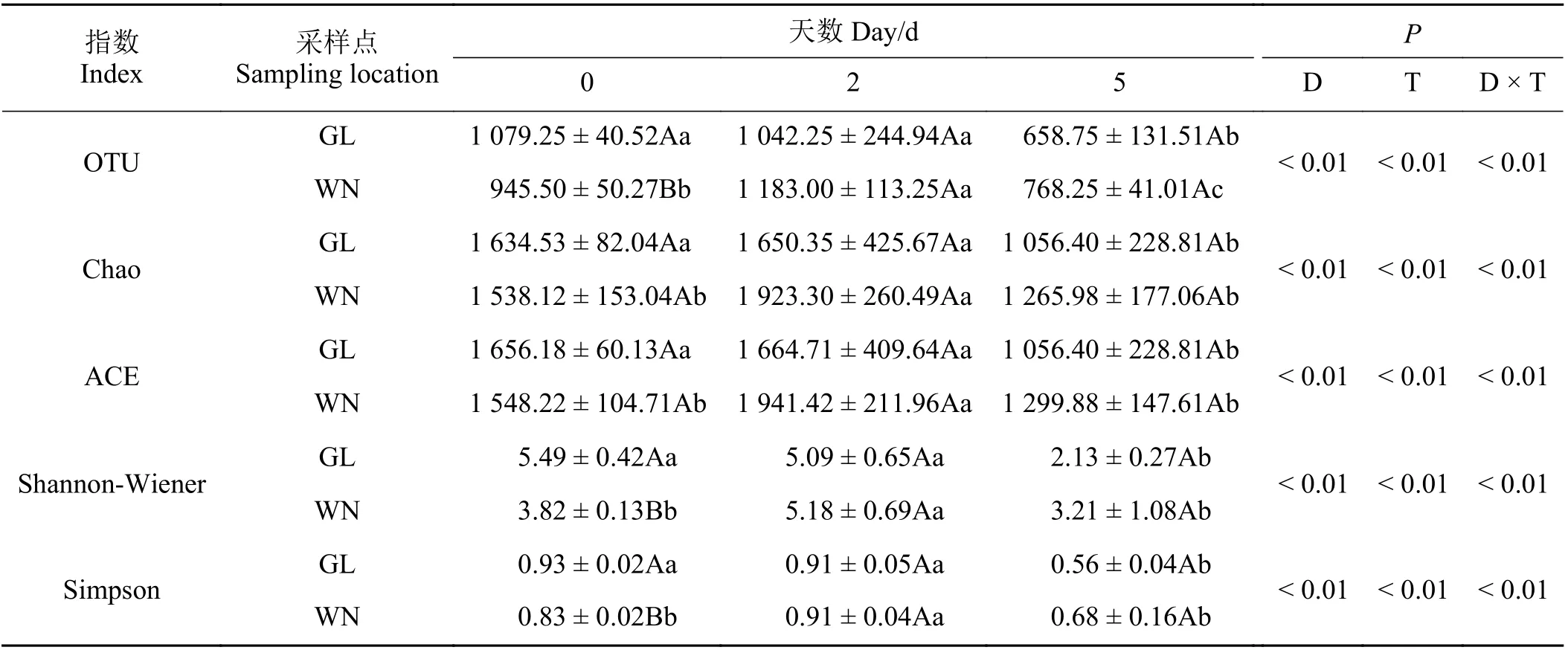
表5 青贮玉米饲料有氧暴露后的细菌α 多样性Table 5 Bacterial alpha diversity of the silage maize feed after aerobic exposure
2.4.2 门分类水平细菌群落组成
随有氧暴露天数延长,两组主要的门水平优势菌群均由厚壁菌门变为变形菌门(图1)。有氧暴露5 d 与0 d 相比,关岭组变形菌门的相对丰度增加了5.69 倍,厚壁菌门的相对丰度减少了93.42%,而威宁组变形菌门的相对丰度增加了32.49 倍,厚壁菌门的相对丰度减少了86.91%,未分类菌门细菌的相对丰度增加了17.00 倍。有氧暴露5 d 与全株玉米原料相比,关岭组变形菌门的相对丰度减少了1.28%,厚壁菌门的相对丰度增加了27.27%,放线菌门和未分类菌门细菌的相对丰度分别增加了1.00倍和2.00 倍,其他门水平菌群均检测不到;威宁组变形菌门的相对丰度增加了2.22 倍,厚壁菌门的相对丰度减少了82.26%,未分类菌门细菌的相对丰度增加了50.00%,其他门水平菌群均下降或检测不到。关岭组在0 和5 d 时变形菌门的相对丰度高于威宁组,厚壁菌门的相对丰度低于威宁组。关岭组全株玉米原料变形菌门的相对丰度高于威宁组,厚壁菌门的相对丰度低于威宁组。
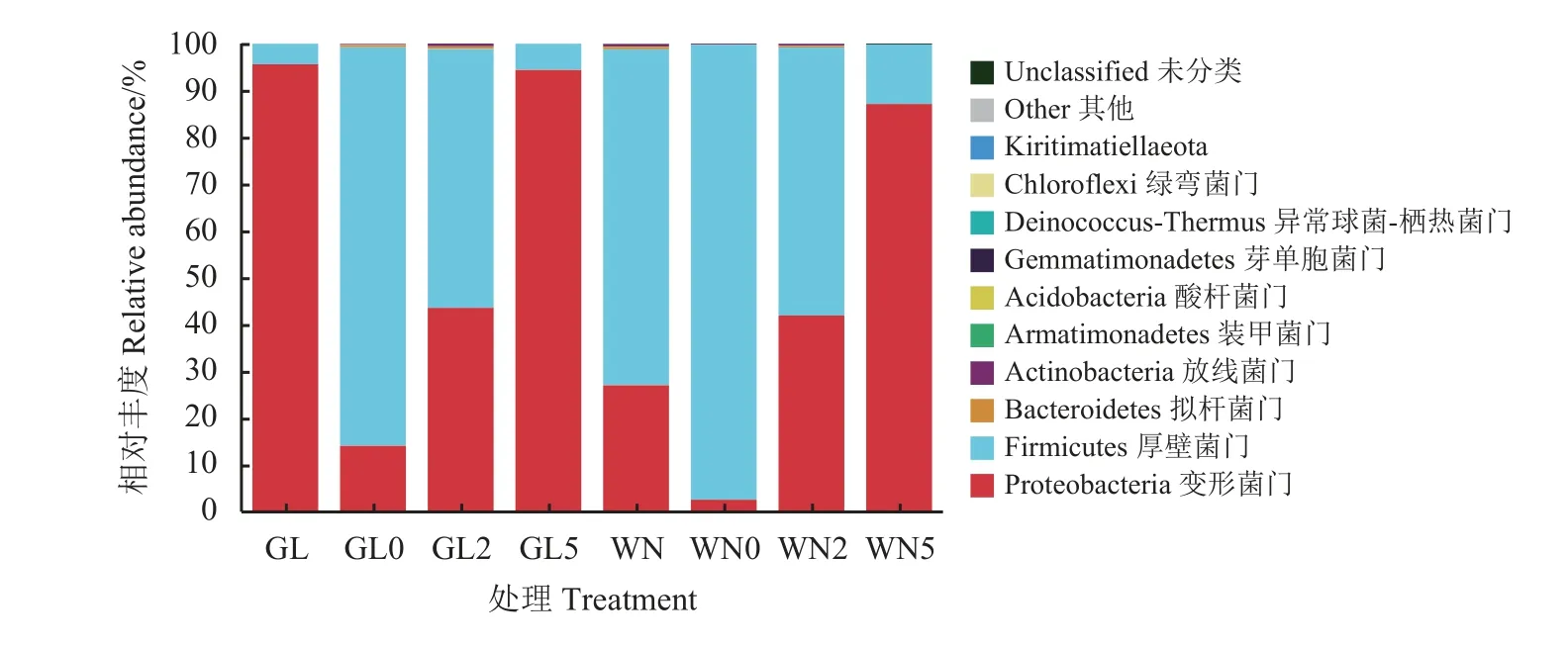
图1 全株玉米青贮饲料有氧暴露后的门水平细菌群落图Figure 1 Phylum-level bacterial community diagram of whole-plant corn silage after aerobic exposure
2.4.3 属分类水平细菌群落组成
随有氧暴露天数延长,两组主要的属水平优势菌群均由乳杆菌属变为醋酸杆菌属(Acetobacter)(图2)。有氧暴露5 d 与0 d 相比,关岭组乳杆菌属的相对丰度减少了97.96%,醋酸杆菌属的相对丰度增加了245.92倍,葡糖杆菌属(Gluconobacter)的相对丰度增加了66.67%;威宁组的乳杆菌属的相对丰度减少了95.99%,醋酸杆菌属的相对丰度增加了211.53 倍,葡糖杆菌属的相对丰度增加了48.00 倍,寡养单胞菌属(Stenotrophomonas)、假单胞菌属(Pseudomonas)、泛菌属(pantoea)、未分类菌属和其他菌属的相对丰度分别增加了1.94倍、96.60倍、3.63倍、7.86倍和5.57倍,两组其他属水平菌群丰度均下降。有氧暴露5 d 与全株玉米原料相比,关岭组和威宁组乳酸菌属的相对丰度分别减少了44.36% 和45.93%,醋酸杆菌属的相对丰度分别增加了43.47 倍和32.92倍,其他菌属的相对丰度分别增加了2.67 倍和81.48%;威宁组寡养单胞菌属、假单胞菌属和葡糖杆菌属的相对丰度分别增加了28.21%、6.75 倍和6.35 倍,两组其他属水平菌群丰度均下降。关岭组在0 和5 d时的乳杆菌属细菌的相对丰度均低于威宁组,醋酸杆菌属细菌的相对丰度均高于威宁组。关岭组全株玉米原料乳杆菌属和醋酸杆菌属细菌的相对丰度均低于威宁组。

图2 全株玉米青贮饲料有氧暴露后的属水平细菌群落图Figure 2 Genus-level bacterial community diagram of whole-plant corn silage after aerobic exposure
3 讨论与结论
3.1 有氧暴露后全株玉米青贮饲料的营养品质动态变化
干物质含量可反映底物营养成分的高低。本研究中干物质含量显著下降,与Queiroz 等[32]研究的玉米青贮后在有氧稳定性期间的干物质含量降低的结果相似。这是由于有氧暴露后的好氧微生物活动性增强,粗蛋白被快速分解,产生大量的氨态氮,同时大量的营养物质被消耗利用,从而导致干物质的含量下降[33]。本研究中,两组有氧暴露后的中性洗涤纤维和酸性洗涤纤维含量均显著降低,这与万学瑞等[8]研究的全株玉米青贮饲料的中性洗涤纤维和酸性洗涤纤维含量在有氧暴露后0~7 d 内缓慢增加,而在有氧暴露15 d 后急剧下降的结果相似。相关研究表明,假单胞菌属和寡养单胞菌属细菌具有分解纤维素的能力,其分析结果表示寡养单胞菌属与酸性洗涤纤维含量呈负相关关系[34]。关岭组在有氧暴露2 和5 d 时的酸性洗涤纤维含量低于威宁组,其原因可能是关岭组在有氧暴露2 d 时的寡养单胞菌属相对丰度高达10.08%,远高于威宁组的1.77%。
3.2 有氧暴露后全株玉米青贮饲料的发酵品质动态变化
氨态氮占总氮比的含量可作为青贮玉米饲料粗蛋白降解程度的重要指标[35]。试验中两组的氨态氮含量均显著增加,和Schmidt 等[36]研究苜蓿青贮饲料的氨态氮含量在发酵开袋后显著增加的结果相似。这是由于氧气暴露后,好氧微生物迅速繁殖,分解氨基酸,生成氨、硫化氢和胺类物质,粗蛋白被大量降解,即氨态氮增加[37]。pH 的变化能直观反映出青贮饲料的变化[38],而过高的pH 会增加微生物的多样性[39]。本研究中两组的pH 均上升,这与Shi 研究不同菌剂对全株玉米好氧稳定期内pH 变大的结果一致[40]。这是由于氧气的供给促进好氧微生物利用可溶性碳水化合物、乳酸和乙酸等营养物质大量繁殖,可溶性碳水化合物、乳酸和乙酸被好氧微生物代谢,致使pH 的上升[30]。而关岭组在有氧暴露2 d时的pH 变化不显著及可溶性碳水化合物含量低于威宁组,其原因是2 d 时的粗蛋白降解速度慢,产生的氨态氮等碱性物质过少,可溶性碳水化合物被好氧微生物过多代谢。同时,有氧暴露后威宁组的pH及氨态氮占总氮比含量均低于关岭组,说明在有氧暴露过程中,较低pH 环境可以抑制微生物对蛋白质的降解[41]。由有氧稳定性的测定结果可知,关岭组于有氧暴露120.00 h (5 d)时较临近有氧不稳定,而威宁组于有氧暴露48.00 h (2 d)后就开始了有氧腐败。
3.3 有氧暴露后全株玉米青贮饲料的微生物多样性动态变化
两组在有氧暴露5 d 后的细菌α 多样性较0 d均下降,但在有氧暴露2 d 时有略微上升。这是由于在有氧暴露的前期,原被抑制的各种好气性微生物在氧气的供给下活动性增强,但随有氧暴露天数的延长,有限的营养物质不足以提供其进行代谢繁殖[42],从而导致微生物菌群丰度及群落多样性降低。Li 等[43]研究表明细菌群落组成和多样性主要由有机酸和氨态氮等发酵品质决定的,威宁组在有氧暴露2 d 的细菌α 多样性较0 d 显著增加是由于其有氧暴露0~2 d 的发酵品质显著降低所致。本研究中,两组有氧暴露后的乳杆菌属相对丰度均降低,醋酸杆菌属相对丰度均增加,与胡宗福等将全株玉米青贮并开袋暴露于空气3 d 后乳杆菌属丰度下降严重的结果一致[13]。这是由于开窖后,在氧气的暴露下乳酸菌被抑制生长,而原被抑制生长的好氧细菌在氧气滋养下开始繁殖[14]。本研究中两组有氧暴露5 d 后的醋酸杆菌属相对丰度较0 d 时的增加幅度均较大,且5 d 时的pH 均超过了常规成功青贮要求的临界值(4.2),这是由于醋酸杆菌属细菌可以氧化乳酸生成二氧化碳和水[15],导致pH 升高,从而直接导致饲料有氧腐败。因此,可推断5 d 时的变形菌门中的醋酸杆菌属相对丰度增加是导致饲料发生腐败的原因之一。本研究中,威宁组有氧暴露2 d时的魏斯氏菌属相对丰度较0 d 增加了7.27%,同时在有氧暴露0~2 d 的细菌α 多样性显著降低。这是由于魏斯氏菌属中有的魏斯氏菌可产生具有抑菌活性的物质,以抑制腐败菌和好氧微生物的生长[44]。
综上,两个区域全株玉米青贮饲料有氧暴露期间,营养品质和发酵品质均随暴露天数的延长而降低,关岭组的有氧稳定性高于威宁组;不同取样地环境因子对全株玉米原料表面附着微生物的群落结构有较大影响,有氧暴露后两个区域青贮饲料的细菌α 多样性均降低,主要门、属水平的优势菌群均为变形菌门和醋酸杆菌属,其整体呈现出一致的变化规律;建议关岭组青贮玉米饲料于开窖后2 d内尽快使用,尽可能地减少营养品质损失和提高饲料安全性;威宁组于开窖后(0 d)应立即使用,饲喂有氧暴露2 d 后的青贮料会影响反刍动物的健康。
猜你喜欢
河南医学研究(2022年19期)2022-10-19 00:44:18
乡村地理(2019年2期)2019-11-16 08:49:48
生态学报(2019年11期)2019-07-08 06:18:58
乡村地理(2018年1期)2018-07-06 10:34:10
乡村地理(2017年4期)2017-09-18 02:54:18
乡村地理(2017年4期)2017-09-18 02:53:52
贵州科学(2016年5期)2016-11-29 01:25:49
中国农资(2016年9期)2016-08-01 07:51:14
食品工业科技(2014年23期)2014-03-11 18:19:08
现代检验医学杂志(2014年6期)2014-02-02 03:01:54
