
侵蚀区植被恢复过程中土壤有机碳稳定性的研究进展
2022-10-10 14:20肖胜生房焕英徐佳文常梦琦秦晓蕾彭文杰刘洪光沈发兴
水土保持学报 2022年5期
肖胜生, 房焕英, 徐佳文, 常梦琦, 秦晓蕾, 彭文杰, 刘洪光, 沈发兴
(1.江西省土壤侵蚀与防治重点实验室,江西省水利科学院,鄱阳湖流域生态水利技术创新中心,南昌 330029;2.江西省退化生态系统修复与流域生态水文重点实验室,南昌工程学院,南昌 330099;3.江西农业大学林学院,南昌 330045)
土壤是陆地生态系统最大的碳库,土壤碳收支的微小变化会显著影响大气碳浓度。自然植被转变为耕地或遭人为严重破坏时,会导致土壤有机碳(SOC)的损失,而适当的管理和植被恢复可以使退化土壤重新吸存SOC。前期研究结果表明,侵蚀区进行植被恢复可以显著促进SOC积累。
探究SOC的组成、来源和稳定机制是深入认识陆地生态系统碳汇功能的关键。SOC库的相对稳定在维持碳素平衡和应对全球气候变化方面发挥着重要作用。与“增加土壤碳汇”研究成果相比,“稳定现存土壤碳汇”研究相对薄弱。稳汇与增汇同等重要。但是目前总体上侵蚀退化土地植被恢复过程中SOC动态变化、稳定性及其固持有效性等问题尚不清楚,SOC的稳定机制尚未充分揭示。近几年兴起的土壤微生物“碳泵”理论为揭示侵蚀区植被恢复过程中SOC的稳定机制提供了独特视角和方法。已有研究指出,土壤微生物群落是退化土地恢复过程中的游戏规则改变者/变局者,为进一步理解微生物介导的SOC稳定性提供了最新的视角。
为此,本文综述了侵蚀区植被恢复过程中土壤碳素的积累效益及影响有机碳稳定性的关键因素,并在简要介绍微生物“碳泵”理论框架的基础上,重点梳理了微生物介导的SOC稳定性研究进展。最后,从研究对象、研究内容、研究手段和研究土层4个方面指出了该领域的薄弱环节。
1 侵蚀区植被恢复过程中SOC的积累
1.1 侵蚀区植被恢复过程中SOC的积累效益
Lal认为,退化土壤损失碳的60%~75%能通过生态恢复重新固定,并估计全球退化土壤的碳吸存潜力高达3×10~8×10t/a。另外,相对于植被生物量作为碳的临时库,土壤中累积形成的是一种更理想的相对稳定的碳库。从我国水力侵蚀区水土保持历史进程来看,植被恢复变化显著的主要是西北黄土高原区、南方红壤丘陵区和西南石漠化区3个2级区。总体上,与黄土高原水土保持植被恢复过程中SOC数量和质量的研究成果相比,南方红壤丘陵区和西南石漠化区就显得相对薄弱得多。小流域综合治理40年来,这2个侵蚀区的植被覆盖状况发生了翻天覆地的变化,特别是南方红壤丘陵从曾经的“江南红色沙漠”转变到如今的“绿水青山”。
学者们在黄土高原典型侵蚀区进行了诸多研究。如马祥华等指出,随着植被的演替恢复,SOC含量表现为先减小后增大的趋势。SOC含量减小时,地上植被参与了SOC的消耗,从而提高了植被的丰富度;由于植被的凋落、腐解,SOC含量提高,这样就形成植被与土壤的正向互动状态。Feng等研究证实,退耕还林还草是黄土高原地区生态系统固碳增加的主要原因,2000-2008年黄土高原生态系统固碳量增加了96.1 Tg,该区域生态系统已从碳源转变为碳汇。相比于退耕还林等植被人工恢复,退耕还草等植被自然恢复更有利于表层(1 m)土壤SOC的积累。
大量研究表明,随着造林/植被恢复时限的延长,土壤中的碳储量迅速增加,并可以在几十年内恢复到与附近未受干扰森林相当的水平。同时,由于气候条件有利于植物快速生长,这种特点在热带地区表现得更为明显。Yu等研究表明,亚热带森林生态系统具有很高的CO吸收强度,其净生态系统生产力NEP(以C计)达到(362±39)g/(cm·a),存在一个被忽略的东亚季风区(20°—40°N)碳汇功能区。江西省赣州市是我国南方红壤侵蚀区的典型代表,水平竹节沟+乔灌草混交补植是区域内严重侵蚀地最常见的植被恢复模式。有研究表明,经过近40年的综合治理,植被得到了有效恢复,土壤碳素积累明显。与未治理对照样地相比,治理4,14,24,34年后样地1 m深度内SOC吸存速率分别达到0.74,0.14,0.54,0.58 t/(hm·a)。福建师范大学相关团队以长汀为研究区进行了长期研究,何圣嘉等研究表明,侵蚀退化红壤地建植马尾松林后,林地表层(0—20 cm)SOC吸存速率在15~25年内达到最大,马尾松恢复30年后SOC吸存速率约为0.39 t/(hm·a),自马尾松建植起至演替为当地顶级群落(次生林)全过程中SOC平均吸存速率约0.16 t/(hm·a)。
另外,关于侵蚀区植被恢复背景下不同层次SOC积累的差异(表层土壤碳积累对植被恢复的响应大于深层土壤)、SOC吸存速率的时间非线性表现(恢复后期SOC增加速率低于恢复早期)、强度侵蚀退化(或者说依然存在较强坡面侵蚀)导致马尾松林地碳素积累效益不明显等方面也做了深入研究。总体上,侵蚀区植被恢复后土壤碳素积累效益与多种因素有关,如退化程度、土壤矿物学特征与养分状况(特别是N、P养分含量)和造林时间与恢复年限等。
1.2 侵蚀区植被恢复过程中土壤碳素积累的影响因素
侵蚀区植被恢复过程中主要通过3种途径对土壤碳素积累产生积极影响:植被恢复畅通了凋落物和细根归还的途径、土壤结构改善与团聚体保护机制、碳素坡面侵蚀损失得到抑制。
侵蚀区植被恢复过程中土壤碳素积累效益不明显,或者说与周边未受扰动的次生林相比还存在较大的SOC吸存潜力,这主要是因为侵蚀区自然生态环境具有潜在的脆弱性,随坡面径流损失的碳素占有较大比例。Jackson等基于对全球2 700多个土壤剖面的分析,得到在较为湿润的地区,乔、灌木入侵引起的地上生物量增加有可能被SOC的流失所抵消。侵蚀逆境下,SOC含量主要是2种正向输入过程(凋落物分解和细根分解释放)与2种负向输出过程(土壤呼吸和侵蚀损失)共同作用的结果。侵蚀劣地植被恢复后,增加了向土壤中输入的碳素,相应增大了土壤呼吸速率以及碳素侵蚀损失的可能性,特别是降雨充沛、山高坡陡的西南石漠化区和南方红壤丘陵山地。以南方红壤区严重侵蚀退化山地为例,其植被恢复措施主要是等腰环山开挖水平竹节沟,辅以乔灌草补植。在这个过程中,工程措施对坡面碳素侵蚀的抑制有可能大于植被重建对土壤碳素的归还作用。下一步要加强这4个关键过程综合实证研究,明确侵蚀逆境下SOC主要来源的相对贡献以及不同去向的相对比例问题,摸清侵蚀区植被恢复中土壤有机碳的吸存机理。
另外,Dou等研究认为,南方红壤侵蚀区植被恢复后非保护性土壤有机碳能够逐渐向保护性有机碳转化,以及土壤老碳分解较慢也是土壤有机碳提高的重要原因。另外,植被恢复过程中由于养分限制、树种选择不当以及对植物-土壤-微生物相互作用缺乏了解等也会限制SOC积累。
2 SOC稳定性和固持有效性的关键影响因素
增加陆地生态系统碳汇功能非常重要,但是稳定现存的土壤碳汇同等重要。从缓和大气CO浓度上升角度考虑,能在土壤中长时间保存的SOC其固碳意义才更大。植被恢复后积累的碳素能否长期储存在土壤中,很大程度上取决于其稳定性,尤其是在人为扰动增加与气候变暖的情况下。
2.1 植被恢复对SOC稳定性的影响
已有研究表明,随着植被恢复,植物种类组成结构的变化可显著改变凋落物的质量和数量、根构型及其分泌物,显著提高SOC的含量、组分和稳定性;红壤侵蚀区的研究指出,植被恢复初期(7~11年)主要以非保护性有机碳的形式积累,而随着恢复时间的延长,土壤非保护性有机碳逐渐向保护性有机碳转化,这有利于土壤有机碳的长期保持。但有研究认为,得到恢复的森林土壤碳素周转效率仍然较高,从而降低了SOC稳定性,如Poeplau等发现,造林有利于稳定碳库转变为活性碳库。
在侵蚀地植被恢复过程中,土壤总有机碳积累的短期变化并不能指示生态恢复对土壤碳动态的影响。因此,识别更敏感的活性有机碳组分有助于阐明土壤有机碳的动态。植被恢复重新建立了凋落物和细根的物质循环途径,特别是逐渐增多的有机物质输入量为微生物提供了可利用的C源、N源,促进了微生物活性及微生物量的升高,从而促进了土壤活性有机碳组分含量。邓翠等研究表明,马尾松恢复过程中土壤活性碳库在恢复7~10年后逐渐向缓效性碳库(物理保护态的团聚体与粉粒和黏粒组合成)转化;惰性碳库储量随恢复年限不断增加,但其分配比例保持较稳定水平。土壤有机碳以可溶性有机碳(DOC)与颗粒态有机碳(POC)积累逐渐转化为缓效性碳库积累为主,进而影响惰性碳库的积累,有利于SOC的长期保持。Xiao等同在亚热带红壤侵蚀区马尾松林地的研究表明,在恢复35年期间,DOC和微生物量碳(MBC)等活性碳组分一直呈线性增加状态。Liu等通过微生物降解培养试验,首次发现随着黄土高原植被演替恢复,可溶性有机质(DOM)的生物降解能力逐渐降低,并对温度变化的承受能力不断增加,进而促进了DOM含量的积累,为黄土高原土壤有机质的稳定维持途径提供了理论依据。
2.2 SOC的保护与稳定机制
土壤碳稳定性取决于SOC不同组分的构成及其与环境的相互作用。不同的SOC组分有着不同的来源,且对影响因素有着不同的响应机制。总体上,SOC的保护与稳定受到土壤团聚体、土壤矿物和微生物等环境条件的作用与影响。土壤团聚体能够将SOC包裹起来,通过空间上的物理隔离保护,使其免受微生物的分解利用。SOC的稳定性通常与稳定性团聚体的数量相联系,而土壤团聚体的稳定性受土地利用方式的影响,大团聚体对SOC的保护作用较弱,微团聚体对SOC的保护相对较强。除去物理隔离,微团聚体中微生物来源的有机碳还可以与矿物结合形成复合体从而相对稳定地留存在土壤中,即矿物学保护机制在有机碳稳定化过程中也发挥了重要作用。研究者运用原位光谱学观测技术研究表明,有机碳在土壤中的长期固持不是由其分子结构决定的,而主要是由其所处生态系统决定。SOC与无机矿物,特别是纳米尺度的铁铝氧化物通过表面相互作用形成有机无机复合体,在更长时间尺度上控制着SOC固持。
随着研究手段与分析技术的飞速发展,目前对于SOC形成和稳定机制的认知发生了巨大的变革,逐渐从早期的腐殖质理论、土壤团聚体保护理论、土壤矿物保护理论等向微生物转化和调控理论转化,或者说,SOC的保护与稳定机制存在多样性及差异性,是团聚体的物理保护-土壤矿物的结合-微生物代谢过程相互作用、相互依存的结果。SOC能否顺利接触到微生物(即空间不可接近性)以及周围环境条件是否适宜于微生物的分解则成为SOC稳定的重要限制因素,积累在更细粒级部分的微生物代谢产物可能具有更长的驻留时间。
与天然林或未扰动森林相比,侵蚀区人工恢复的森林更应受到关注。生物量碳的积累与SOC的积累并不总是同步或等效的,较高的生物量或凋落物产量并不一定意味着较高的SOC储量。凋落物质量通过调节凋落物分解过程中的产物去向来控制SOC的积累,演替后期的成熟林其高质量的凋落物分解产物向土壤转移的比例更高,土壤碳库拥有更为丰富的有机碳源,且SOC的存储环境更加稳定,这可能是原始森林能够在土壤中持续积累SOC的原因。我国南方红壤侵蚀区的研究表明,森林转换过程中的扰动加速了深层土壤稳定性有机碳的分解,导致人工恢复森林具有更高的微生物可利用碳比例和矿化速率。最近有学者研究表明,通过汇总分析全球53个国家地区、264个野外研究的近2.6万条数据发现,与结构简单的人工林相比,天然林具备更好的碳存储能力。
不同土壤层次SOC稳定性也有所差异。Chen等在福建首次发现,与表层土壤相比,在严重侵蚀退化地上恢复起来的人工林底层土壤均拥有更高的极易分解有机碳含量和快得多的相对碳矿化速率,其原因可能与小分子量、简单可溶性有机物向底层土壤的选择性迁移、底层土壤中底物与微生物空间分离以及底层土壤中微生物的r型生长策略有关。
3 基于微生物“碳泵”理论的SOC稳定研究
3.1 微生物“碳泵”理论框架
土壤微生物存在双重身份,既可以作为分解者调控非微生物来源碳的周转,也可以作为贡献者调控微生物来源碳的形成。在厦门大学焦念志“海洋微生物碳泵”(microbial carbon pump, MCP)理念的基础上,中国科学院沈阳应用生态研究所Liang等2017年首次提出“土壤微生物碳泵”,强调了土壤微生物同化合成产物是土壤稳定有机碳库的重要贡献者,其概念体系可以较好地阐释SOC的来源、形成与截获过程,为揭开微生物介导的固碳过程的真实面纱提供了可能。
MCP理论强调的是土壤微生物在有机碳固存中的正效应,其将活性有机碳转化为较为稳定的合成代谢形式,并通过微生物残留物的续埋效应不断输入到稳定土壤有机碳库。相比于土壤MBC周转更快、含量较低,微生物残体碳(MRC)在土壤中周转时间更长,对稳定性碳库的形成具有重要贡献,并可表征微生物对SOC的长期贡献。氨基糖是目前应用最为广泛的土壤微生物死亡残体的生物标识物,由于氨基糖具有异源性,因此常用氨基葡萄糖/胞壁酸比值指示真菌和细菌对土壤有机碳的相对贡献。
3.2 微生物残体碳对SOC的贡献
在许多生态系统的研究中MRC都呈现出与SOC极强的正相关关系,植物残体碳(木质素)与SOC负相关,微生物残体是SOC积累的稳定组分和重要贡献。Liang等利用吸收马尔科夫链(absorbing markov chain)首次模拟并估算出土壤中MRC是MBC的40倍,并整合估算出温带区农田、草地和森林生态系统表层土壤中MRC占SOC比例分别为55.6%,61.8%,32.6%。与之类似的是,Wang等也得到全球尺度上农田、草地和森林0-20 cm表层土壤中MRC对SOC的平均贡献分别为51%,47%,35%,表明SOC形成的微生物途径(即微生物残体的续埋效应)在耕地和草地土壤占主要地位,而植物途径(即难分解植物残体的物理迁移)在森林中占主导地位,并且继续得到这种贡献在农田土壤中随土层深度(0—150 cm)的增加而降低,而草地和森林土壤中随土层深度的增加而增加。MRC不仅是发育良好土壤SOC的主要来源,Wang等证明了MRC也是初始土壤(生物结皮覆盖下黄土高原沙质土壤)SOC的重要来源。
在土壤微生物中,根系相关的菌根真菌占微生物生物量的30%以上,菌根真菌使用植物衍生碳来进行生物量生产和呼吸代谢,将C从活菌丝(有机酸和胞外酶等)和死亡菌丝生物量释放到土壤中,直接贡献POC和矿质结合态有机碳(MAOC)组分。Saskia等在美国印第安纳州的温带落叶林中,通过C和N自然丰度分析与氨基糖标志物测定相结合的方法揭示了腐生真菌对POC的贡献为4%~53%,对MAOC的贡献为23%~42%,而外生菌根的贡献则相反。该结果表明,相比于植物残体,真菌对稳定态SOC的贡献更大,改变真菌群落可以加强土壤SOC长期固存的贡献。
但是由于微生物“碳泵”研究视角起步不久,结论还存在不一致性,如Matthew等对美国温带森林的培养试验表明,微生物的生长、效率和周转与矿物相关的有机碳呈负相关,微生物残体的来源、有机碳形成的替代途径、激发效应和土壤的非生物特性等因素可以强烈地将微生物的生长、效率和周转与矿物相关的SOC分离,微生物残体的大量产生不是温带森林SOC持久性的主要驱动因素。
3.3 微生物介导的SOC稳定研究
目前,基于微生物“碳泵”理论,研究者对不同生态系统的固碳过程和稳碳机制进行了研究,并对土壤微生物“碳泵”的影响因素进行了深入分析。在全球尺度上,降水、温度和土壤pH是控制微生物残体的重要气候和环境因素,较低的温度和土壤pH(如在温带和寒带生态系统中)促进真菌和细菌残体积累。土壤微生物群落作为土壤MCP的核心和载体,微生物残体的产生主要受微生物生长速率(MGR)、微生物碳利用效率(CUE)和微生物生物量周转率(MTR)3个主要指标控制。MGR、CUE、MTR与微生物介导的有机碳截获过程并非简单的线性关联,主要与土壤微生物的化学计量特征密切相关。土壤微生物的化学计量特征可以指示微生物养分需求、代谢过程以及酶学特征等与群落内秉特征相关的过程,有助于判断微生物群落驱动的土壤MCP功能对外界干扰的应对策略。尽管这些特征可能是SOC形成和分解的核心,但对微生物生理特征的控制及其在环境梯度上对SOC的影响还没有得到很好地阐明,阻碍了对SOC库如何响应环境变化的预测,这是目前研究的不足之处。Wang等研究结果表明,黄土高原沙质土壤裸沙、藻结皮、藻-苔藓结皮、苔藓-藻结皮和苔藓结皮5个阶段组成的生物土壤结皮形成序列中,细菌残体主要贡献MAOC库,而真菌残体在POC中更为丰富,<2%黏粒含量和微生物N是这种黄土高原沙质土壤中稳定碳积累的重要影响因素。依据室内培养试验,有学者对微生物介导的土壤有机碳形成来源、稳定性进行了一些研究,其研究模式见图1。
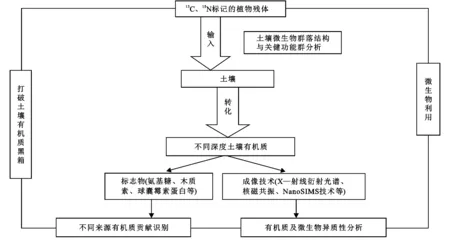
图1 基于培养试验的土壤有机碳研究模式
培养试验过程中,涉及到的微生物残体碳的积累等关键观测指标计算公式为:





基于图1中的研究模式,Zhu等在草地和森林生态系统中都进行了相关研究,结果表明,高寒草甸的增温和干旱控制试验通过增加底层土壤氮限制,降低了CUE,从而降低了微生物“碳泵”效率;亚热带森林样地中,植物途径(植物多样性提高)主导了0—10 cm表层土壤SOC的积累,而植物物种丰富度的提高促进了30—40 cm底层土壤中DOM和N素的有效性,进一步提高了氨基糖、矿物结合有机质等微生物组分有机碳含量,微生物途径主导了底层土壤SOC的积累。质地不同的土壤中,影响氨基糖积累的环境因子明显不同。井艳丽等研究表明,在落叶松纯林改造中引入赤杨能显著提升根际及非根际土壤氨基糖葡萄糖含量,并揭示全氮是驱动土壤中氨基糖发生变化的主要因子。
4 研究不足与展望
从前面的分析可以看出,国内外学者针对典型侵蚀区植被恢复过程中SOC的积累效益、机制、关键因素等方面做了大量的研究,取得了丰富的研究成果。但同时也清醒地认识到,由于土壤的黑箱特性、有机碳稳定影响因素的复杂性、微生物在有机碳存储作用上的两面性,侵蚀区植被恢复过程中SOC的稳定性和固持有效性研究还存在不少薄弱环节,建议重点围绕4个方面展开下一步的研究工作。
4.1 研究对象
前期围绕植被恢复对包括SOC含量在内的土壤质量的研究主要侧重于黄土高原等侵蚀区,对国内南方红壤丘陵区(花岗岩发育的红壤本身碳密度低)、西南石漠化区(水土流失严重)等其他侵蚀区的研究相对薄弱。这些侵蚀区开展了长时期的水土保持生态恢复工作,植被恢复进程强烈影响了土壤碳素积累机制、过程和效益。特别是南方红壤丘陵是我国仅次于黄土高原的第二大水土流失区。历史上,花岗岩发育的红壤大面积严重退化,碳密度低。20世纪80年代以来,每平均5年1个周期,以水平竹节沟+乔灌草混交补植为主要手段的生态恢复与以坡改梯经果林开发为代表的小流域水土保持综合治理在该区域广泛推行,植被覆盖度显著增加,水土流失有效抑制,生态环境明显改善,是非常理想的研究区。
4.2 研究内容
总体上,目前侵蚀区植被恢复过程中研究较多的还是SOC及其不同组分的变化,而不同恢复阶段积累的土壤碳素活性如何,是否能够长久保存尚不清楚,特别是微生物介导的SOC及其稳定性对植被恢复的响应机制尚未充分挖掘。微生物群落驱动的土壤MCP功能的具体评估需要考虑其时间尺度效应,探究不同生态与时间尺度的微生物特征对土壤MCP的影响更有助于阐明土壤MCP储碳机制与环境响应策略。因此,以长时间序列水土保持综合治理背景下土壤微生物介导的SOC稳定状态和机制为切入点开展研究有助于科学评估侵蚀劣地植被恢复后土壤固持有机碳的质量、效率和时间以及水土保持碳汇功能,也可以加深水土保持与碳循环相互关系的认识。
4.3 研究手段
SOC的形成与稳定是一个古老且复杂的科学问题。微生物是SOC循环过程最关键的驱动者。在复杂的土壤环境中,通过野外调查,借用MCP的理念与方法,比较侵蚀区不同植被恢复阶段或群落下微生物量与群落结构、SOC及其关键组分、氨基糖等指标的差异,可将植被恢复、微生物过程与SOC循环过程三者联系起来;通过盆栽试验,进行C脉冲标记,模仿侵蚀区不同植被恢复措施或阶段设置不同凋落物输入梯度或过程,进行土壤培养,测定微生物群落结构、氨基糖和碳矿化等指标的变化,监测MGR、CUE和MTR等指标,可以量化凋落物输入的激发效应和续埋效应,揭示MCP机理。野外样地的调查与室内培养试验相结合,应该对验证和解析侵蚀区植被恢复过程中MCP调控的SOC稳定机制起到很好的作用。
4.4 研究土层
深层土壤(20—100 cm)所储存的SOC远超过表层土壤(0—20 cm),但目前关于SOC稳定性,包括MRC含量及其对SOC贡献的研究多数集中在土壤表层,土壤剖面和母质中的情况尚未得到充分挖掘。深层土壤中大部分微生物因受到能量限制而处于“碳饥饿”状态或休眠状态,从而使深层SOC对环境变化比表层SOC更敏感,目前零星的报道证实了这一点,但总体上这方面研究较为鲜见。有研究表明,亚热带地区充沛的降雨和强烈的生物地球化学循环过程导致淋溶作用强烈,DOM的淋溶迁移给深层土壤微生物带来大量的碳源和养分,可能会增加微生物生长和活性,促进MCP机制,因此加强南方红壤丘陵和西南石漠化等侵蚀区的相关研究显得更有可行性和必要性。
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
红领巾·探索(2021年2期)2021-08-26
红蜻蜓(2021年8期)2021-08-23
课外生活·趣知识(2021年2期)2021-05-24
学校教育研究(2021年24期)2021-03-28
农业与技术(2020年12期)2020-07-04
军事文摘·科学少年(2020年2期)2020-03-19
儿童故事画报·自然探秘(2019年12期)2019-02-02
环球时报(2019-01-29)2019-01-29
