
不同激素组合对番茄离体再生和相关基因表达的影响
2022-10-07 12:27:02金宝霞王伟杰朱晓林魏小红
浙江农业学报 2022年9期
金宝霞,王伟杰,朱晓林,王 贤,魏小红
(甘肃农业大学 生命科学技术学院,甘肃省作物遗传改良与种质创新重点实验室,甘肃省干旱生境作物学重点实验室,甘肃 兰州, 730070)
番茄(Mill)为一年生草本植物,属茄科(Solanacea)番茄属,是一种重要的经济作物和转基因模式植物。近年来,利用转基因技术进行分子育种成为番茄种质资源创新的重要途径,而番茄高效离体再生体系的建立是进行遗传转化的重要前提。植物离体培养主要经过脱分化和再分化2个关键步骤,生长素和细胞分裂素是诱导外植体分化的关键因素,两者配合使用能够提高植株的再生率和转化率。番茄离体再生试验中关于外植体脱分化、再分化的研究大多集中于筛选外植体、优化培养基激素类型和比例等应用方面。
近几年来,在现代分子生物学研究的推动下,外植体脱分化、再分化的内在调控机制相继在拟南芥、大豆、茶树、苹果等植物中被报道。拟南芥中已鉴定出多个与激素调控脱分化、再分化相关的基因,如LBD转录因子(16、17、18和29)、AP2/ERF转录因子(1、2、3和4)等调控脱分化过程发生。ART1转录因子、AP2/ERF转录因子(1、2、26和)和1基因等调控拟南芥再分化过程。LBD转录因子在生长素存在条件下诱导脱分化,生长素通过增强拟南芥bZIP家族成员AtbZIP59蛋白与LBD蛋白的互作,直接调控下游靶基因-的表达促进愈伤组织的形成。AP2/ERF是创伤响应通路的中心调控因子,1在创伤部位通过B类细胞分裂素响应子ARR1和ARR12信号通路促进细胞脱分化。1编码AP2/EREBP结构域,在细胞分裂素存在条件下过表达1可大幅提高拟南芥根外植体的芽再生效率。26表达量在芽再生初期增加,将该基因敲除后芽的再生率降低,说明其可以促进拟南芥芽再生。1基因部分功能的缺失对拟南芥不定芽形成具有抑制作用,但对细胞分裂素和生长素没有显示相关作用。大豆芽诱导试验中,RAV2-like转录因子同系物的表达也受到细胞分裂素的诱导,因此或许能够通过细胞分裂素促进不定芽的再生。上述研究通过转录组分析发现大量差异表达基因与植物激素信号转导通路密切相关,推测植物激素参与调控植物组织再生。
拟南芥作为保守模式植物,其脱分化与再分化关键调控基因对其他植物离体再生的分子机制研究具有重大指导意义。到目前为止,番茄中关于不同激素类型与浓度配比调控外植体脱分化、再分化分子机制的研究还未见报道。因此,本试验以品质优良的樱桃番茄J6为材料,筛选番茄中与拟南芥同源的脱分化、再分化基因序列,使用实时荧光定量PCR(qRT-PCR)技术分析不同激素组合处理下子叶和下胚轴2种外植体在脱分化、再分化过程中的基因表达量,同时结合脱分化率、再分化率对离体再生机制进行关联分析,为樱桃番茄离体再生体系的完善探索新思路。
1 材料与方法
1.1 材料
樱桃番茄材料J6由甘肃张掖益新泉蔬菜育种公司馈赠。
1.2 试剂与仪器
吲哚-3-乙酸(IAA,货号B21810)、玉米素(ZT,货号B25447)、6-苄氨基腺嘌呤(6-BA,货号B24213),所有激素纯度均为分析标准品,HPLC≥98%,均购自上海源叶生物科技有限公司。
1.3 方法
1.3.1 无菌苗获得与外植体切取
J6种子清水浸泡12 h后在超净工作台用75%乙醇浸泡30 s,然后用10% NaClO溶液消毒10 min,用无菌水冲洗3次后吸干水分,接种于MS基本培养基。取9 d苗龄无菌苗的子叶和下胚轴为外植体。子叶切取方法:剪去子叶尖端和基部,将子叶横切为0.5 cm×0.5 cm的方块,叶片正面向上接种于脱分化诱导培养基。下胚轴切取方法:将下胚轴剪成约0.5 cm长的茎段,接种于脱分化诱导培养基。培养条件:光周期为16 h光照/8 h黑暗,相对空气湿度为60%左右,温度为25.0 ℃,光照强度为2 000 lx,暗培养光照为0 lx。
1.3.2 脱分化与再分化基因的筛选与引物设计
查阅文献确定拟南芥中的关键脱分化基因和再分化基因,然后分别在番茄基因组中进行同源比对找到相关转录因子;根据转录因子名称在https://www.arabidopsis.org/(拟南芥官网)中查询得到该转录因子蛋白序列;根据转录因子蛋白序列在https://www.string-db.org/(蛋白相互作用数据库)中进行比对得到番茄中对应的同源基因名称,并在番茄CDS数据库进行比对,分别得到脱分化基因序列和再分化基因序列;将基因序列输入NCBI—primer BLAST进行引物设计。基因注册号与引物序列见表1。

表1 番茄脱分化与再分化基因引物信息
1.3.3 不同激素组合对外植体的培养
剪取子叶和下胚轴,分别在以下3种激素组合的MS培养基中进行培养。A组:2 mg·L6-BA+0/0.1/0.2/0.3 mg·LIAA(依次编号为A1、A2、A3、A4);B组:2 mg·LZT+0/0.1/0.2/0.3 mg·LIAA(依次编号为B1、B2、B3、B4);CK:不加任何激素。激素类型与浓度选择参照文献[2-6]。每个培养皿中接20个外植体,每个处理设置3个重复。以10 d为周期,观察外植体诱导情况,计算第10天(脱分化前期)和第20天(脱分化后期)脱分化率,第60天的再分化率。每隔15 d进行继代培养。
脱分化率(%)=产生愈伤组织外植体总数/接种的外植体数×100;
再分化率(%)=产生不定芽的外植体数/接种的外植体数×100。
1.3.4 RNA提取
采用AG RNAex Pro RNA提取试剂盒(艾科瑞生物公司)提取RNA。
1.3.5 cDNA合成
采用Evo M-MLV反转录试剂盒Ⅱ(艾科瑞生物公司)的操作步骤进行cDNA合成。
1.3.6 qRT-PCR
采用2×SYBR Green qPCR Master Mix(None ROX)(武汉赛维尔生物科技有限公司)试剂盒进行qRT-PCR实验。
1.4 数据分析
采用Excel 2013、SPSS 22软件进行数据分析,基因相对表达量采用2-ΔΔC法计算。
2 结果与分析
2.1 不同激素组合对子叶、下胚轴脱分化的影响
J6外植体在脱分化第10天形成初期愈伤组织,第10天到第20天完成愈伤组织生长并为不定芽的发生做准备。由图1可知,接种第10天,CK处理的子叶和下胚轴外植体没有明显变化(图1-A、图1-B),其余处理的子叶外植体叶片颜色变淡,呈浅绿色,叶片边缘切口轻微卷起并出现现白色愈伤组织团(图1-C);下胚轴呈透明淡绿色胚状体,切口的一端或两端出现膨大(图1-D)。培养至第20天,CK处理的子叶和下胚轴依然无明显变化,未分化出愈伤组织;B1处理的子叶愈伤组织呈黄绿色并零星出现绿色芽点(图1-E),其他处理的子叶愈伤组织膨大增厚,叶片卷曲更加明显;下胚轴愈伤组织变粗,质地更为疏松,边缘有白色小突起(图1-F)。
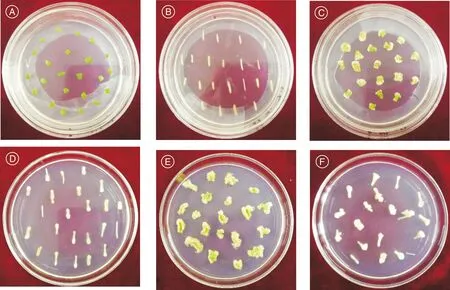
A,CK处理10 d的子叶;B,CK处理10 d的下胚轴;C,A2处理10 d子叶;D,B1处理10 d下胚轴;E,B1处理20 d子叶;F,B1处理20 d下胚轴。A, The cotyledons of 10 days under CK treatment; B, The hypocotyls of 10 days under CK treatment; C, The cotyledons of 10 days under A2 treatment; D, The hypocotyls of 10 days under B1 treatment; E, The cotyledons of 20 days under B1 treatment; F, The hypocotyls of 20 days under B1 treatment.图1 不同激素组合下子叶和下胚轴的脱分化情况Fig.1 Dedifferentiation of cotyledons and hypocotyls under different hormone combinations
采用不同激素组合对子叶和下胚轴进行脱分化诱导,结果(表2)表明,激素类型不影响子叶的脱分化率,培养至第10天,所有处理的子叶脱分化率已达到100%;在第10天,A组的下胚轴脱分化率优于B组。A组合中,下胚轴在A2处理下脱分化率最高,为88.33%;B组合中,脱分化率在B2、B4处理下最高,均为65.00%。培养至第20天,下胚轴脱分化率增加不超过3.34百分点。

表2 不同激素组合对J6子叶、下胚轴脱分化率的影响
2.2 不同激素组合对子叶、下胚轴再分化的影响
由图2可知,外植体培养至第60天,CK的部分下胚轴未通过愈伤组织的分化而直接长出嫩芽(并非不定芽),并伴随大量根的生长(图2-A);适宜激素组合处理的子叶愈伤组织生长为绿色球状,表面组织较为松软,长出的独立不定芽数量更多更健壮,畸形芽数量少(图2-B);子叶和下胚轴未分化芽的外植体表面僵硬,底部与培养基接触部分发生褐化,诱导再分化的效果较差(图2-C、2-D)。通过比较再分化率和不定芽生长状态,发现子叶比下胚轴更适合诱导再分化。

A,CK处理60 d的下胚轴;B,B1处理60 d的子叶;C,B4处理60 d的子叶;D,A3处理60 d的下胚轴。A, The hypocotyls of 60 days under CK treatment; B, The cotyledons of 60 days under B1 treatment; C, The cotyledons of 60 days under B4 treatment; D, The hypocotyls of 60 days under A3 treatment.图2 不同激素处理下子叶和下胚轴的再分化情况Fig.2 Redifferentiation of cotyledons and hypocotyls treated with different hormone combinations
采用不同激素组合对子叶和下胚轴进行再分化诱导,由表3可知,子叶在B组处理下的再分化率明显优于A组,并且2种组合处理下的再分化率随IAA浓度升高而逐渐降低。A组中,子叶在A1处理下再分化率最高为22%;B组中,其再分化率在B1和B2处理下达到最高,分别为67.00%和58.00%,无显著性差异。下胚轴在不同激素组合的再分化率显著低于子叶,其再分化率在A3组合处理下最高,仅为15%。因此,选择子叶作为外植体材料,在激素组合2.0 mg·LZT+0/0.1 mg·LIAA (B1和B2处理)处理的离体再生效果最好。

表3 不同激素组合对J6子叶、下胚轴再分化率的影响
2.3 子叶、下胚轴脱分化基因表达量
2.3.1 子叶脱分化基因表达量
将子叶作为外植体,第10天,脱分化基因Solyc09g066260.3.1、Solyc06g083930.2.1、Solyc01g091420.2.1、Solyc04g072900.1.1、Solyc12g056980.1.1和Solyc04g054910.3.1在A组的表达量较B组更高(图3)。A组中,所有脱分化基因的表达量均显著上调,表达量为CK的5.56~64.60倍;B组中,仅基因Solyc09g066260.3.1、Solyc06g083930.2.1和Solyc04g072900.1.1在B4处理下表达量上调,为CK的5.03~10.20倍。第20天(图4),B组处理的基因表达量较A组明显上调。A组中,所有基因表达量仅在A4处理上调,为CK的7.48~35.20倍;B组中,基因Solyc06g083930.2.1、Solyc01g091420.2.1和Solyc04g072900.1.1在B2处理下高表达,分别为CK的12.12~44.05倍;基因Solyc09g066260.3.1、Solyc12g056980.1.1和Solyc04g054910.3.1在B3处理下表达量大幅上调,为CK的17.40~29.21倍。

编号1-6分别代表Solyc09g066260.3.1、Solyc06g083930.2.1、Solyc01g091420.2.1、Solyc04g072900.1.1、Solyc12g056980.1.1和Solyc04g054910.3.1。柱上无相同小写字母表示差异显著(P<0.05)。下同。1-6 represented Solyc09g066260.3.1, Solyc06g083930.2.1, Solyc01g091420.2.1, Solyc04g072900.1.1, Solyc12g056980.1.1 and Solyc04g054910.3.1, respectively. Data marked without the same lowercase letter indicated significant differences at P<0.05. The same as below.图3 不同激素组合第10天子叶愈伤组织中脱分化基因的相对表达量Fig.3 Relative expression level of dedifferentiated genes in callus of cotyledons treated with different hormone combinations at the 10th day
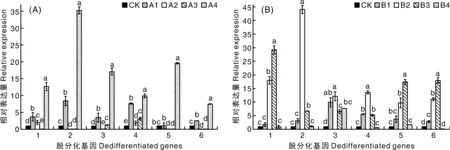
图4 不同激素处理第20天子叶愈伤组织中脱分化基因的相对表达量Fig.4 Relative expression level of dedifferentiated genes in callus of cotyledons treated with different hormone combinations at the 20th day
将脱分化情况与相关基因表达量结合进行分析,脱分化第10天,子叶脱分化率在不同激素组合下无差异,均达到100%,脱分化基因在A组处理下的表达量均显著上调;而在B组处理下,仅Solyc09g066260.3.1、Solyc06g083930.2.和Solyc04g072900.1.1基因在B4处理下表达上调。推测脱分化率与基因表达无明显联系,说明子叶在脱分化前期(第10天)可能不受到脱分化基因的调控。脱分化第20天,B组处理的子叶愈伤组织形态更利于不定芽分化与生长,脱分化基因Solyc01g091420.2.1、Solyc04g072900.1.1和Solyc04g054910.3.1的表达量在B1和B2组合下显著上调,表明这3个脱分化基因对脱分化后期愈伤生长可能产生重要影响。
2.3.2 下胚轴脱分化基因表达量
以下胚轴为外植体,第10天,A组处理的脱分化基因Solyc09g066260.3.1、Solyc06g083930.2.1、Solyc01g091420.2.1、Solyc04g072900.1.1、Solyc-12g056980.1.1和Solyc04g054910.3.1表达量显著上调(图5)。Solyc09g066260.3.1、Solyc06g0-83930.2.1、Solyc01g091420.2.1、Solyc04g072900.1.1和Solyc12g056980.1.1随IAA浓度升高呈先升高后下降的趋势,其中,Solyc04g072900.1.1在A2处理下表达量最高,为CK的14.37倍;Solyc09g066260.3.1、Solyc06g083930.2.1、Solyc-01g091420.2.1和Solyc12g056980.1.1在A3处理下表达量达到最高,为CK的17.19~43.35倍;Solyc04g054910.3.1表达量在A4处理下最高,为CK组的25.23倍。B组处理下,仅Solyc09g066260.3.1、Solyc06g083930.2.1、Solyc-04g072900.1.1和Solyc04g054910.3.1在B2、B3处理下表达量上调,为CK的1.36~6.32倍。第20天,A组中Solyc09g066260.3.1、Solyc06g0839-30.2.1、Solyc01g091420.2.1、Solyc12g056980.1.1和Solyc04g054910.3.1分别在不同处理下表达量上调,为CK的1.06~5.29倍;B组中,6个脱分化基因表达量均在B2处理中上调,为CK的4.5~23.35倍;Solyc09g066260.3.1、Solyc06g08-3930.2.1和Solyc04g072900.1.1在B3处理中的表达量上调,为CK的2.03~6.55倍(图6)。

图5 不同激素处理第10天下胚轴愈伤组织中脱分化基因的相对表达量Fig.5 Relative expression level of dedifferentiated genes in callus of hypocotyls treated with different hormone combinations at the 10th day
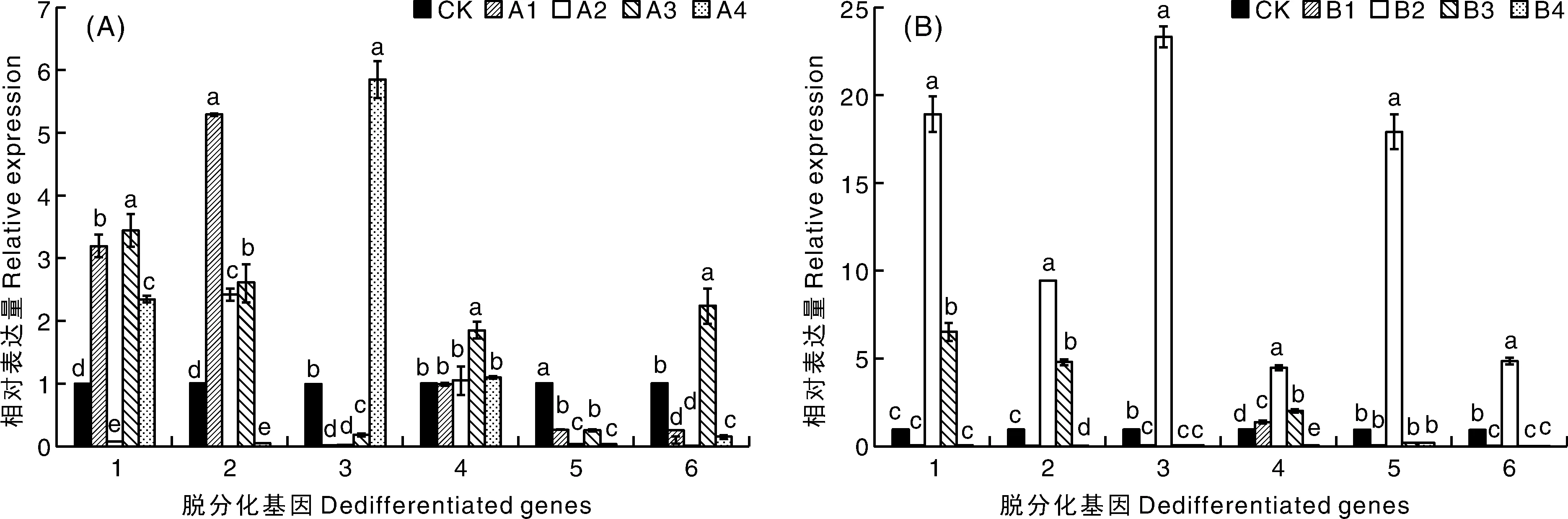
图6 不同激素处理第20天下胚轴愈伤组织中脱分化基因的相对表达量Fig.6 Relative expression level of dedifferentiated genes in callus of hypocotyls treated with different hormone combinations at the 20th day
结合脱分化情况与相关基因表达量分析可知,脱分化第10天,下胚轴在A组的脱分化率和脱分化基因表达量显著高于B组,表明其在脱分化前期可能受到脱分化基因的正调控。脱分化第20天,下胚轴在不同激素组合下愈伤组织形态均无明显差异,脱分化基因仅在B2处理下显著上调。表明下胚轴愈伤组织形态与脱分化基因表达没有关联,激素对脱分化后期生长和脱分化基因表达没有产生影响。
2.4 再分化基因表达量
2.4.1 子叶再分化基因表达量
子叶再分化第60天,A组处理下,再分化基因Solyc05g013540.1.1、Solyc03g082980.3.1、Solyc12g042210.2.1、Solyc02g022850.1.1、Solyc-08g066840.3.1和Solyc10g052510.2.1仅在A3处理中表达量上调,为CK的1.82~14.71倍,其余处理变化均不显著;B组处理下,基因表达量均显著上调,其中Solyc05g013540.1.1、Solyc03g082980.3.1、Solyc12g042210.2.1、Solyc08g066840.3.1和Solycv1-0g052510.2.1表达量随IAA浓度升高呈先升高后下降的趋势,在B2处理中达到最高,分别为CK的29.60、6.56、13.37、18.83和34.41倍;Solyc02g022850.1.1在B1处理中表达量最高,为CK的46.59倍(图7)。

编号7-12分别代表Solyc05g013540.1.1、Solyc03g082980.3.1、Solyc12g042210.2.1、Solyc02g022850.1.1、Solyc08g066840.3.1和Solyc10g052510.2.1。下同。7-12 represented Solyc05g013540.1.1, Solyc03g082980.3.1, Solyc12g042210.2.1, Solyc02g022850.1.1, Solyc08g066840.3.1 and Solyc10g052510.2.1, respectively. The same as below.图7 不同激素处理第60天子叶愈伤组织中再分化基因的相对表达量Fig.7 Relative expression level of redifferentiation genes in callus of cotyledons treated with different hormone combinations at the 60th day
将再分化率与再分化基因表达量结合分析发现,子叶在B组合中的再分化率和再分化基因表达量均显著高于A组。其中,子叶在B1、B2处理中不定芽生长情况更佳,且基因Solyc05g013540.1.1、Solyc02g022850.1.1和Solyc10g052510.2.1表达量显著上调,表明上述3个基因对子叶再分化可能具有重要作用。
2.4.2 下胚轴再分化基因表达量
由图8可以看出,下胚轴再分化第60天,A组中再分化基因表达量较低;B组中基因表达量在B1、B2处理中显著上调,其中B2处理的表达量最高,为CK的6.05~23.67倍。

图8 不同激素处理第60天下胚轴愈伤组织再分化基因的相对表达量Fig.8 Relative expression level of redifferentiation genes in callus of hypocotyls treated with different hormone combinations at the 60th day
结合下胚轴再分化率与再分化基因表达量可知,再分化率在所有处理中均较低,再分化基因表达量只在B1、B2处理中上调,表明激素对下胚轴再分化过程和再分化基因的表达没有影响。
3 结论与讨论
细胞分裂素和生长素配合使用诱导分化的效果好于单个激素诱导,因此筛选外源激素的种类和比例是提高外植体分化效率的关键。本研究中,以番茄材料J6子叶为外植体,在ZT+IAA组合下诱导脱分化、再分化的效果显著好于6-BA+IAA组合,其中,在低浓度IAA条件下,即含2.0 mg·LZT+0/0.1 mg·LIAA的MS培养基更适合番茄离体再生。刘炜炜等在对加工番茄子叶进行离体培养时发现,不同激素组合对诱导形成愈伤组织形态和不定芽存在差异,而对其出愈率没有影响,本研究结果与此一致。王全华等、陈丽萍等在番茄离体再生试验中发现,ZT诱导产生不定芽的效果明显优于6-BA,在MS+2.0 mg·LZT+0.5 mg·LIAA和MS+2.0 mg·LZT+0.2 mg·LIAA处理下不定芽诱导率最高,本研究结果与此相似。谢雯琦等选用不同浓度的细胞分裂素6-BA和ZT分别与生长素IAA组合,对黄樱桃-2号子叶进行愈伤组织和再生芽的诱导,发现MS+2.5 mg·L6-BA+0.2 mg·LIAA培养基效果最佳。说明不同番茄品种诱导外植体分化的最佳激素组合不尽一致。
在外植体离体再生过程中,大多数研究仅关注激素在脱分化、再分化阶段发挥的功能,但很少有人研究相关基因表达的差异。近些年来,陆续有研究在拟南芥、大豆、茶树等植物中分离鉴定了与脱分化、再分化相关的转录因子,并进行了功能验证。对拟南芥脱分化、再分化阶段差异表达基因进行功能分析发现,生长素和细胞分裂素的合成、代谢途径的差异表达基因最为丰富。大豆基因同系物在不同植物之间具有功能保守性,并且在细胞分裂素存在下可以促进植物不定芽的再生。Fan等研究表明,16、17、18、29可以诱导不同器官、组织产生愈伤组织,这些基因的异位表达可以使外植体在没有添加外源激素的情况下起始愈伤组织的形成。茶树组织脱分化与再分化过程中5、18、29和18等与生长素、细胞分裂素相关基因的表迖量产生了显著变化。不同浓度的生长素和细胞分裂素对组织培养中的激素调控通路是否有影响还尚不清楚。本研究对最适激素组合下离体再生过程与相关基因表达量结合分析发现,子叶脱分化后期愈伤组织形态生长可能受到脱分化基因Solyc01g091420.2.1、Solyc04g072900.1.1和Solyc04g054910.3.1的正向调控,推测这3个基因分别是番茄LBD转录因子家族和AP2/ERF转录因子家族中调控脱分化过程发生的重要成员;再分化过程可能受到再分化基因Solyc05g013540.1.1、Solyc02g022850.1.1和Solyc10g052510.2.1的正调控,上述3个基因分别与AP2/ERF转录因子、1基因和ART1转录因子为同源基因,推测它们是番茄中促进不定芽形成的重要基因。此外,下胚轴脱分化前期受到脱分化基因(Solyc09g066260.3.1、Solyc06g083930.2.1、Solyc01g091420.2.1、Solyc-04g072900.1.1、Solyc12g056980.1.1和Solyc04-g054910.3.1)的正调控;而脱分化后期生长情况和再分化效果较差,相关基因表达量也较低,推测激素对下胚轴离体再生过程和相关基因表达也有影响。有研究表明,不同植物之间、同种植物不同品种间,以及不同外植体间的脱分化、再分化能力差别较大,这些差异主要与植物基因型有关。因此,推测J6番茄下胚轴再分化效果差可能主要由内源基因决定,外源激素类型与浓度对调控再分化基因表达的影响较小。
本研究结果表明,番茄材料J6的子叶比下胚轴更适合离体再生培养,且诱导分化的最佳激素组合为2 mg·LZT+0/0.1 mg·LIAA。试验所选的脱分化和再分化基因在不同外植体的脱分化时期、再分化时期均有不同程度的表达,推测脱分化和再分化同源基因在不同植物间具有功能保守性,为深入研究激素调控番茄脱分化、再分化发生的分子机制,以及建立更加高效的离体再生体系提供了理论依据。除此之外,本研究还发现了与番茄离体再生可能密切相关的几个脱分化和再分化基因,可在后续研究中通过这些基因的沉默或过表达试验进一步证实它们对番茄离体再生的影响,同时也为离体再生的分子机制研究提供了新的思路。
猜你喜欢
热带海洋学报(2022年6期)2022-12-05 12:53:52
东北师大学报(自然科学版)(2021年1期)2021-03-27 01:18:50
畜牧兽医科技信息(2021年9期)2021-03-05 20:07:56
中国稻米(2019年6期)2019-12-05 03:34:22
山东林业科技(2019年2期)2019-06-03 10:10:54
现代园艺(2017年11期)2017-06-28 11:32:46
上海农业学报(2017年4期)2017-04-10 12:40:22
江西农业学报(2017年12期)2017-02-03 21:49:41
奥秘(2016年10期)2016-12-17 13:13:11
故事作文·高年级(2015年4期)2015-09-07 11:18:13
