
植物花青素代谢途径相关基因的研究进展
2022-10-03 10:19罗天俊
南方农业 2022年16期
罗天俊
(西南林业大学,云南昆明 650051)
植物的花色主要受外部因素和内在因素的影响,外部因素包括光照、温度等一系列环境因素,内在因素是遗传因素,主要指植物细胞内的环境和花瓣的构造等对花色的重要影响。但是,外部因素和内在因素最终都需要作用于花青素(属黄酮类),进而决定花的颜色,花青素是重要的植物色素之一。
近年来,花青素在生物代谢方面的研究已经取得了充分进展,其中有关花青素的代谢途径和相关基因的研究报告相对较多。一些学者发现,圆叶牵牛(Ipomoea purpureaLam.)等观赏植物的开花过程与花青素有着明确的关系,这种关系存在于花青素合成途径遗传因子发育模式与花青素积累之间。花青素的代谢受到多种结构基因、酶、控制基因及外界刺激的调控,如micro RNA、LBD 遗传因子家族、光照、植物激素等。
进入21 世纪以来,在一系列先进技术的作用下,生物技术蓬勃发展,越来越多的相关研究成果逐渐阐明了不同的植物花青素代谢途径的分子控制机制差异。但还有很多经济价值高的植物,其与代谢途径有关的基础机理尚待探索。本文主要总结了与植物花青素代谢途径有关的遗传因子,并总结了利用生物工程技术对植物的表观遗传进行优化改良等的相关理论基础。
1 花青素的结构
花青素以3,5,7-三羟基-2-苯基苯(并)吡喃羊盐(三羟基羊盐、trihydroxyflavylium)为基本碳骨架结构(见图1),根据在苯环B 上不同的取代位置、数量、羟基类型,形成不同的花青素[1]。A、C 环的结构相似,但需修饰增加稳定性,其中修饰作用、溶液环境和花青素二维堆积也导致花青素颜色发生改变[2]。花青素大多具有良好的水溶性,使得许多植物含水量高的组织中含有大量花青素,如花和果实。较高的花青素含量在组织中表现出深红、橙色、蓝色、紫色等性状。

图1 花青素的基本化学结构
目前已发现的天然花青素有500 多种,其中最常见的是芍药色素(Peonidin,Pn)、锦葵色素(Malvidin,Mv)、飞燕草色素(Delphindin,Dp)、矮牵牛色素(Petunidin,Pt)、矢车菊色素(Cyanidin,Cy)、天竺葵色素(Pelargonidin,Pg)6 种。
2 花青素的代谢途径
20 世纪90 年代以来,花青素的代谢途径研究日臻成熟,其基本代谢途径如图2 所示。代谢过程至少涉及12种结构遗传因子(ANS、PAL、AT、C4H、4CL、CHS、DFR、CHI、F3H、F3’H、F3’H、UD3GT)和3 种转录因子(bHLH、MYB、WD40)。结构基因主要用于编码对应类别的酶,转录因子主要用于调控结构基因的活动。
2.1 花青素的合成
如图2 所示,花青素的合成主要分3 个阶段。
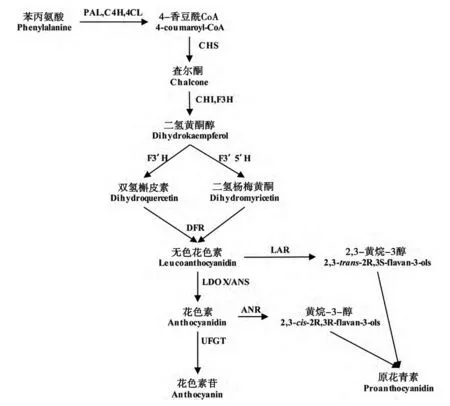
图2 花青素的生物代谢途径图
1)苯丙氨酸按顺序经苯丙氨酸解氨酶(Phenylalanineammonialyase,PAL)、肉桂酸-4-羟化酶(Cinnamate 4-Hydroxylase,C4H)和4-香豆酸辅酶A 连接酶(4-Coumarate Coenzyme A Ligase,4CL)的催化,转变为4-香豆酰CoA。PAL 广泛分布在植物中,是植物代谢途径中重要的调节因子,催化苯基丙氨酸脱氨基,是本阶段反应的限速酶[3]。
2)4-香豆酰CoA 在查尔酮合酶(Chalcone Synthase,CHS)的催化下生成查尔酮(Chalcone),接着在查尔酮异构酶(Chalcone Isomerase,CHI)和黄酮-3-羟化酶(Flavanone 3-Hydroxylase,F3H)的作用下形成二氢黄酮醇(Dihydrokaempferol,DHK),经类黄酮3’-羟化酶(Flavonoid 3’-Hydroxylase,F3’H)和类黄酮3’,5’-羟化 酶(Flavonoid 3’,5’-Hydroxylase,F3’5’H)催化后可分别形成双氢槲皮素(Dihydroquercetin,DHQ)和二氢杨梅黄酮(Dihydromyricetin,DHM)。
3)DHQ 和DHM 在二氢黄酮醇4-还原酶(Dihydroflavonl 4-Reductase,DFR)的作用下生成无色花色素,分别经无色花青素还原酶(Leucoanthocyanidin Reductase,LAR)和无色花青素双加氧酶/花青素合成酶(Leucoanthocyanidin Dioxygenase/Anthocyanidin Synthase,LDOX/ANS)的催化分别形成有色花青素。
2.2 花青素的运输
花青素一般在细胞内合成,在合成相关酶的过程中,通过蛋白质的相互作用,形成可以锚定在内质网膜上的大分子复合体。另外,通过类黄酮糖基转移 酶(UDP-Glycose Flavonoid Glycosyltransferase,UFGT)和尿苷二磷酸葡萄糖醛酸转移酶(UDPGlucuronosyhransferase,UGT)的相互作用和修饰作用,转运到植物液泡中。在当前的研究中,关于花青素的运输机制有如下4 种。1)与液泡膜上的多药耐药相关蛋白(MultildrugResistance-Associated Protein,MRP)及毒物外排转运蛋白(Multidrug And Toxic Compound Extrusion,MATE)有关。液泡上的MATE 根据膜内外的H+/Na+浓度差使H+从液泡内排出,从而将花青素输送到液泡内[4]。如拟南芥功能研究较多的TT12 遗传因子(属MATE 遗传因子家族),在含原花青素的植物组织中特异性表达,定位在液泡膜上,就是这种转运方式。但有学者表示,酰基化之后的花青素苷才能够被MATE 基因家族成员转运[6]。2)依赖谷胱甘肽巯基转移酶(Glutathione S-Transferase,GST)与液泡膜上的多药耐药相关蛋白介导的花青素苷转运的形式。3)与胆红素异位酶同族体(Bilitranslocase-homologue,BTL-homologue)相关。目前关于该项研究还较少,但有研究表明,BTLhomologue 可以转运花青素苷和胆红素(为血红素降解产物)。4)由囊泡介导的花青素的运输。例如,在拟南芥中,液泡产生的囊泡可以直接将花青素输送到相应的空间而不依赖于GST 和ATP。
3 花青素代谢途径相关基因
3.1 结构基因对花青素的影响
1)PAL 是植物代谢途径中的重要限制酶,可以通过催化苯基丙氨酸的脱氨生成反式肉桂酸。PAL 在植物中广泛分布,主要存在于线粒体、叶绿体、过氧化酶体、白血球等细胞器中。PAL 由多基因家族编码,植物的发育情况和环境等因素对PAL 基因的表达有很大的影响。CHS 是植物合成黄酮化合物过程中最重要的酶,它可以通过催化4-香豆酰CoA 和丙二酰CoA缩合而形成查尔酮,为类黄酮类化合物的合成提供了基础碳框架结构[6]。
2)DFR 在植物花瓣的显色中起着重要的作用,其可以将DHQ、DHM 转化为无色花青素。根据研究,蝴蝶草中加入反义DFR 基因,其花色变为蓝色,植物体内的花色素含量降低,但花青素和黄酮醇含量却明显提高。DFR 遗传因子也经常应用于矮牵牛的花色研究。F3’H 可催化黄烷酮生成二氢黄酮醇,研究结果显示,反义抑制草莓(Fragaria×ananassa)中的F3’H 表达之后,阻断了花青素的合成代谢,进而对果实中的花青素含量产生影响,进一步表明了F3’H是植物花青素代谢途径中的一个关键酶[7]。并且DFR和F3’H 的过剩表现,增加了花瓣中红色色素的含量,使花瓣变得更红。根据相关文献,DFR、F3’H和F3’5’H 因为利用共同的底物,所以存在竞争关系,如果DFR 和F3’H 遗传因子含量过高,则会抑制F3’5’H 遗传因子的表达,植物更趋向于合成天竺葵色素苷;在月季中,DFR 基因过量表达,但抑制了F3’5’H 和F3’H 基因的表达,导致月季开粉色花。
3)ANS 遗传因子可以催化无色类黄酮转化为有色类黄酮,因此在植物呈色中起着重要的作用,如洋葱(Allium cepa)的ANS 遗传因子变异后,洋葱的鳞片由红变黄。在缺少ANS 遗传因子的金钟连翘(Forsythia×intermedia)中,花青素的积累受到显著抑制,但是引进DFR 和ANS 遗传因子后,植物体内花青素的积蓄显著增加,胡萝卜素含量明显低于天冬酰胺,花瓣颜色变为橙色[8]。根据相关文献,在植物花色形成过程中,FLS 基因和ANS 存在底物竞争关系,且在花瓣某些部位,FLS 基因会优先表达[9]。因此,可以通过生物技术,将某些基因的表达沉默,进而培育出不同花色的植物品种。
4)通过UFGT 类黄酮代谢途径控制类黄酮合成。该基因在植物中的表达与植物的果实颜色相关,在葡萄、苹果、荔枝中都有相应研究;在葡萄中,通过杂交技术发现,在白色葡萄株和相应的红色突变株中都发现了UFGT 基因,但在红色突变株中,UFGT 基因发生了突变,因此果实表现为红色[10];有研究发现,荔枝果皮中UFGT 的表达与花青素和类黄酮的含量呈现高度正相关[11]。总之,UFGT 基因在植物花青素代谢途径中起着重要的作用。
3.2 转录因子对花青素的影响
植物花青素代谢途径受结构基因影响,转录因子也可以对其产生一定影响,从现有研究来看,转录因子主要有3 种(WD40、bHLH 和R2R3-MYB),这3 种蛋白质通过一系列相互作用形成三元复合体,该三元复合体与结构基因的特定位点结合,用于调节结构基因的活性,并进一步调节花青素的代谢。
1)WD40 参与包括玉米、拟南芥、紫苏等众多植物花青素的代谢途径。例如,在矮牵牛an11 基因突变体中,结构基因DFR 的表达量降低,色素积累受到影响,但是DFR 的表达活性可以逐渐由组成型基因AN2 恢复,说明在花青素合成的控制途径中AN11 基因位于AN2 的上游。
2)根据相关文献,R2R3-MYB 转录因子可直接调控花青素的代谢,如MYB 转录因子PAP1 的过量表达,使花青素代谢途径的下游基因的表达量增加,进而增加花青素积累[12-13]。针对矮牵牛的研究表明,RsMYB1 遗传因子的表达水平与花青素代谢所涉及的结构遗传因子(PAL、CHS、DFF、ANS)的表达水平正相关[14]。MYB 家族除了上述因子外还包含了一类具有转录抑制或负调控效应的因子,如编码R3-MYB蛋白AtMYBL2 的AtMYBL2 基因,缺失该基因的植株,与花青素代谢途径相关基因的表达水平上升,体内花青素累积量也明显增加;而含有mybl2 突变的植株CHI 基因的表达水平较低,但原花青素和黄酮醇含量不变;但MYBL2 表达过高的植株中,CHI 的表达正常。因此,在花青素代谢途径中,R2R3-MYB 蛋白具有关键作用,该基因调控着多个结构基因的表达,进而影响植物体内花青素的积累。
3)bHLH 转录因子是植物中第二大转录因子家族。bHLH 的调节在花青素的代谢途径中起着非常积极的作用。bHLH 转录因子家族中有抑制花青素积累的成员。例如,在拟南芥中,BHLH32 是负调节因子,在BHLH32 缺乏的植物中,DFR 的表达水平增加,花青素的积累增加。
4 小结
花青素是一种分布广、种类多、安全性高、无毒性的天然色素,在许多行业中都有着广泛的应用前景。近年来,关于花青素的代谢途径相关的基因研究取得了较大的突破,目前已发现与花青素生物合成途径相关的代谢基因主要有12 种,结构基因主要有3 种,这些基因在不同物种和组织中有表达差异性,而且目前转录因子与生物代谢基因的相互作用对花青素的遗传表观研究甚少。探究植物花青素的代谢途径相关基因及遗传机制,并利用基因工程开展植物的遗传改良具有重要意义。
猜你喜欢
少儿科技(2022年2期)2022-03-05
数学大王·趣味逻辑(2021年6期)2021-09-27
保健与生活(2021年13期)2021-07-14
发明与创新·小学生(2020年12期)2020-12-21
恋爱婚姻家庭(2019年30期)2019-03-23
热带作物学报(2018年12期)2018-05-14
食品界(2017年12期)2018-01-20
现代家庭·生活版(2017年10期)2017-10-20
课堂内外(小学版)(2017年1期)2017-03-08
