
高粱单宁Tan1 基因分子标记开发及应用
2022-09-29 07:41张丽霞陆萍王春语丛玲王平陈冰嬬陆晓春
山西农业大学学报(自然科学版) 2022年3期
张丽霞,陆萍,王春语,丛玲,王平,陈冰嬬,陆晓春*
(1.辽宁省农业科学院 高粱研究所,辽宁 沈阳 110161;2.黑龙江八一农垦大学 生命科学技术学院,黑龙江 大庆 163002;3.吉林省农业科学院 作物资源研究所,吉林 公主岭 136100)
高粱(Sorghum bicolor(L.)Moench)是世界5 亿多人口的口粮,同时也是动物饲料、酿酒和制造工业乙醇的重要来源[1-3]。单宁(tannins)是高粱重要的次生代谢物之一,通常也称为缩合单宁(condensed tannins,CT)或原花青素(proanthocyanidins,PAs),广泛存在于植物界中,如苹果、蓝莓、草莓、葡萄、蔓越莓、桃子和李子等水果中以及高粱、龙爪稷、蚕豆和大麦等谷物籽粒中[4],但是主要谷类作物如水稻、小麦和玉米籽粒中并不存在单宁[5]。单宁具有多种生物学功能,如抑菌抗病毒、抗过敏、抗氧化、消炎、抗肿瘤,促进免疫、紫外线保护以及免受昆虫、鸟类对谷物籽粒的噬食[6]。此外,单宁可以提高酒的品质[7-8]。在我国,约80%的高粱籽粒用于酿酒,如驰名中外的茅台酒、汾酒、泸州老窖及其他高档酒均以高粱为原料酿成[9]。单宁含量的高低直接影响高粱的用途。鉴于单宁的营养价值和对消化率的影响,食物和动物饲料中含有少量的单宁对人体健康和动物都是有益的,如幼畜和家禽的饲料中添加含有少量单宁的高粱可以抵御痢疾以及反刍动物的气胀病[10-11]。但是当食物和动物饲料中单宁含量较高时,会降低蛋白质、纤维素、淀粉、脂肪、维生素以及矿物质的消化吸收,降低食物的营养价值,甚至导致病变[12]。
不同高粱材料籽粒的单宁含量差异较大,这主要取决于调控高粱单宁含量的基因不同。在拟南芥中,单宁生物合成的转录调控途径已经进行了深入地研究,成功克隆了大量关键基因,包括DFR(dihydroflavonol-4-reductase)、ANS(anthocyanidin synthase)、ANR(anthocyanidin reductase)和LAR(leucoanthocyanidin reductase)等[13]。利用遗传学方法通过分析种皮透明突变体(transparent testa,tt),获得了3 种调控单宁合成的必需蛋白,即AtTT2/AtMYB123(R2R3-MYB)[14],AtTT8/AtbHLH042(R/b-like basic helix-loop-helix)[15]和AtTTG1(transparent testa glabra,WD-40 domain protein)[16]。这3 种蛋白形成MBW 复合体调控单宁的合成[17]。单宁合成调控途径在高粱中研究较少,2012 年调控高粱单宁含量Tannin1(Tan1)基因被成功克隆,是一个WD-40 domain转录因子,与AtTTG1高度同源[18]。截止目前,Tan1共有4 种等位基因型,即Tan1、tan1-a、tan1-b和tan1-c[19]。3 种 等 位 变 异 由 于 碱 基 的 插 入、缺失和突变造成蛋白序列改变或者提前终止,使Tan1 蛋白功能丧失导致籽粒无单宁或低单宁。Tan1基因对调控高粱籽粒单宁含量发挥着举足轻重的作用。
本研究通过比较野生型Tan1和3 种不同等位变异的编码区序列以及含有不同等位基因型的高粱籽粒单宁含量,开发分子标记为培育无单宁或低单宁专用型高粱品种提供快速、有效的分子检测手段。
1 材料与方法
1.1 试验材料
583 份自然群体材料和290 份栽培种来自印度、美国、中国和非洲国家,现由辽宁省农业科学院高粱研究所收集、保存。供试材料种植于辽宁省农业科学院沈阳试验基地,待抽穗开花后,取新鲜叶片用于基因组的提取。每份材料选取3~5 株套袋自交后,取均匀、饱满的籽粒用于单宁含量测定。
1.2 试验方法
1.2.1 基因组DNA 提取
取大约100 mg 新鲜叶片,利用天根公司“植物基因组DNA 提取试剂盒”(目录号:DP305)提取总DNA,详细操作步骤参考试剂盒说明书,DNA于-80℃冰箱保存备用。
1.2.2Tan1等位基因型检测
利用DNAMAN10.0 软件设计引物Tan1-F(5’-TCTCTCGAGTCCCAGCAG-3’)和Tan1-R(5’-GGTAACACTGACCGAGTC-3’),PCR扩增片段包括编码区全长及部分5’UTR、3’UTR序列,经测序和比对后检测Tan1基因在不同群体材料中的基因型变异。
1.2.3 单宁含量测定
(1)将4 g 高粱籽粒全部放入粉碎机中,充分粉碎,粉碎后的样品全部通过1.0 mm 孔径的筛子,第一次没有通过筛子的样品继续粉碎,确保所有样品全部过筛,然后充分混匀。
(2)称取样品约0.1 g,精确至0.001 g,加入1 mL 灭菌的超纯水,80 ℃水浴30 min。
(3)水浴后8000×g,4 ℃离心10 min 后将上清液转移到新的离心管中,参考单宁含量测定试剂盒的步骤测定单宁含量(Cohesion Biosciences,CAK1060)。
1.2.4 分子标记开发
依据Tan1不同等位基因型编码区序列的碱基突变、插入缺失,根据PCR 产物大小、有无酶切位点等开发分子标记。tan1-b由于10 bp(GCAGCGGCGG)的插入,利用引物Tan1-BZF1/Tan1-BZR1 扩 增,Tan1扩 增 片 段 大 小 为257 bp,tan1-b扩增片段大小为267 bp。tan1-a由于编码区580 位G 缺失产生EcoN I 新的酶切位点,利用引物Tan1-BZF2/ Tan1-BZR2 扩增、酶切后,Tan1为723 bp,tan1-a为319 bp 和404 bp 两 条带。tan1-c由于编码区1054 位A 突变为T 导致DdeI 酶 切 位 点 缺 失,Tan1-BZF3/ Tan1-BZR3 扩增、酶切后,Tan1为109 bp 和56 bp 两条带,tan1-c为165 bp。引物序列及详细信息见表1。
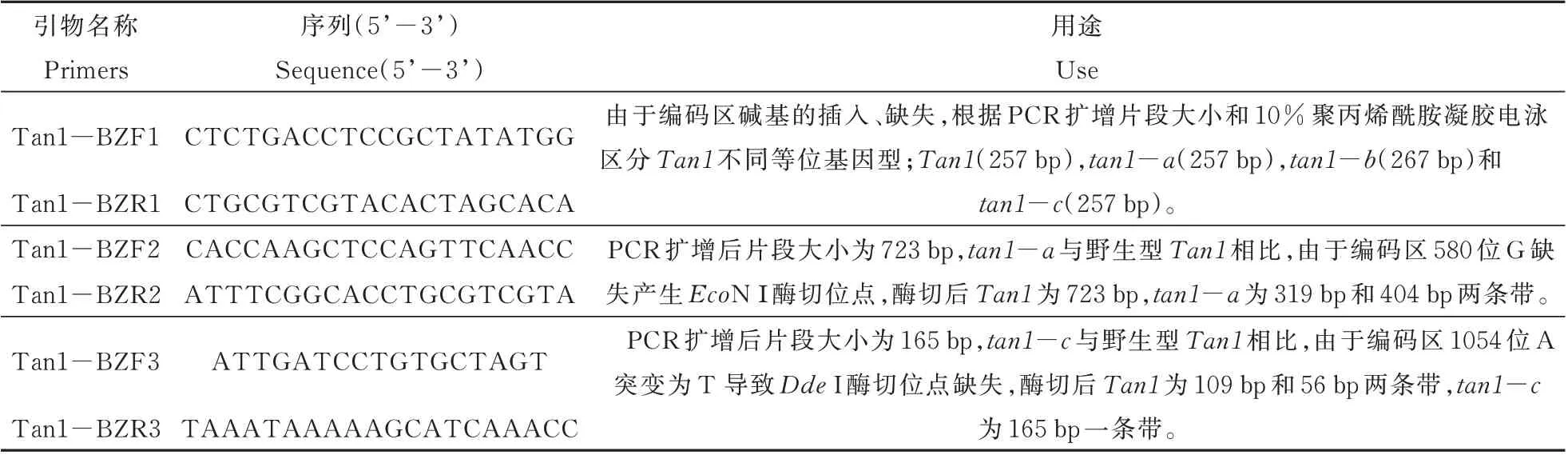
表1 引物序列及用途Table 1 Primer sequences and uses
1.2.5 聚丙烯酰胺凝胶电泳
玻璃板用酒精擦洗干净,并用密封胶条、夹子和垫片将玻璃板固定好。将8 mL 30%丙烯酰胺预制胶(上海迪申生物技术有限公司,货号:B1610156)、4 mL 5×TBE 溶液、27.5 mL 蒸馏水、450 μL 10% APS 和50 μL TEMED 充分混匀迅速灌入装好的玻璃板中,插上梳子,室温放置15 min以上,配置成6%聚丙烯酰胺凝胶。将制备好的胶板放置于已经加入1×TBE 缓冲液的垂直电泳槽中,每个PCR 扩增样品(酶切样品)上样1.5 μL,130 V、电泳40~60 min 后关闭电源。取胶,放入硝酸银染色液(0.5 g AgNO3充分溶于500 mL 蒸馏水)染色15 min,染色后的胶用蒸馏水冲洗3 次,然后放入显影液(10 g NaOH、0.2 g NaHCO₃充分溶于500 mL 蒸馏水中,再加入5 mL 甲醛混匀,现用现配),轻摇至胶显出清晰的条带为止。照相,存储,记录、分析数据。
2 结果与分析
2.1 Tan1 不同等位基因型序列分析
不同等位基因型的成功鉴定是分子标记开发的前提。根据现有的研究结果,归纳总结出调控高粱单宁含量Tan1基因的4 种等位基因型编码区序列和蛋白序列变异情况(表2、图1、图2)。野生型Tan1编码区序列全长1062 bp,蛋白序列全长353 aa。tan1-a与Tan1相比共有2 处发生变化,即编码区580 位G 缺失、798 位G 突变为T,造成蛋白序列移码和提前终止,蛋白序列全长仅有298 aa。tan1-b编码区925 位和926 位之间插入10 bp(GCAGCGGCGG),由于片段的插入导致蛋白序列提前终止,蛋白全长为318 aa。tan1-c共发生3处变异,即1054 位A 突变为T、1057 位G 缺失 和1059 位C 突变为T,由于1054 位碱基突变导致三联密码子AGG 突变为TGG、1057 位单碱基缺失发生移码导致终止密码子TGA 丢失,使得蛋白序列延长至388 aa。由此可见,3 种等位变异均造成Tan1 蛋白C 端序列发生了较大变化。

表2 Tan1 不同等位基因型编码区变异位点Table 2 Variant sites in coding region of different Tan1 alleles

图1 Tan1 不同等位基因型编码区序列分析Fig.1 Analysis of coding sequences of different Tan1 alleles

图2 Tan1 不同等位基因型蛋白序列分析Fig.2 Analysis of protein sequences of different Tan1 alleles
2.2 Tan1 不同等位基因型单宁含量
tan1-a、tan1-b和tan1-c3 种 等 位 变 异 均 对Tan1 蛋白C 端序列产生了或大或小的影响,是否影响Tan1 蛋白功能正常发挥而导致单宁含量发生变化?对来自世界各地的583 份自然群体材料进行Tan1等位基因型鉴定和单宁含量测定。首先,利用引物Tan1-F/Tan1-R 对自然群体材料Tan1基因编码区序列进行扩增、测序、比对发现分别含有488 份Tan1、54 份tan1-a、27 份tan1-b和14份tan1-c(图3A)。其次,利用“单宁含量测定试剂盒”测定了583 份材料籽粒的单宁含量,其中95 份含有3 种不同等位变异基因型的籽粒单宁含量均低于0.5%,而488 份含有Tan1野生型的材料籽粒单宁含量从低到高都有分布(图3B)。由此可见,与AtTTG1一样,C 端蛋白序列对Tan1 蛋白功能发挥非常重要,C 端蛋白序列变异导致单宁含量显著降低或者无单宁。

图3 583 份自然群体材料Tan1 等位基因型和单宁含量分析Fig.3 Analysis of Tan1 alleles and tannin contents in 583 germplasms
2.3 栽培种Tan1 不同等位基因型和单宁含量
为了初步探究4 种Tan1等位基因型在育种中的应用情况,我们对290 份来自世界各地栽培种的Tan1等位基因型和单宁含量进行测定,结果发现其中190 份含有Tan1野生型的高粱籽粒单宁含量仍然从低到高都有分布,而100 份含有tan1-a、tan1-b和tan1-c突变基因型的高粱籽粒单宁含量较 低 或 者 无 单 宁,含 有Tan1、tan1-a、tan1-b和tan1-c基因型高粱材料的比例分别为65.52%、7.59%、26.21% 和0.69%,由此可见,tan1-a和tan1-b广泛应用于低单宁或者无单宁品种培育中,tan1-c应用相对较少(图4)。
图4 290 份栽培种Tan1 等位基因型和单宁含量分析Fig.4 Analysis of Tan1 alleles and tannin contents in 290 cultivars
2.4 Tan1 不同等位基因型分子标记开发
为了快速鉴定Tan1不同等位变异类型,根据编码区序列开发实用的分子标记。tan1-b与野生型Tan1相 比,在925~926 位 之 间 插 入10 bp(GCAGCGGCGG)(图1A)。利用插入片段设计Tan1-BZF1/R1 引物,当检测片段大小为267 bp时,该材料基因型为tan1-b,含有其他等位基因型的材料扩增片段大小为257 bp(表1,图5A)。tan1-a与野生型Tan1相比,由于编码区580 位G缺失产生EcoN I 新酶切位点,经Tan1-BZF2/R2引物扩增和EcoN I 酶切后,Tan1片段大小为723 bp,tan1-a则产生319 bp 和404 bp 两条带(表1,图5B)。tan1-c与野生型Tan1相比,由于编码区1054 位A 突变为T 使酶切位点DdeI 缺失,Tan1-BZF3/R3 引 物PCR 扩增和DdeI 酶切 后,tan1-c片段大小为165 bp,Tan1则有109 bp 和56 bp 两条带(表1,图5C)。
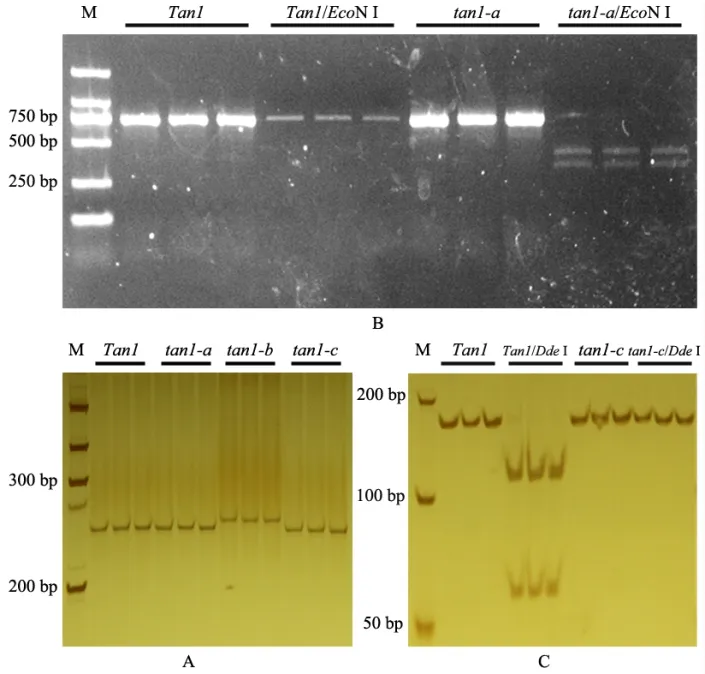
图5 Tan1 不同等位基因型分子标记开发Fig.5 Developing molecular markers for Tan1 different alleles
3 讨论
Tan1基因在不同物种中都存在高度同源的基因,但是生物学功能差异较大。与Tan1基因高度同源的AtTTG1基因在拟南芥中共获得了78 个不同的等位变异突变体(ttg1),通过对启动子、3'UTR、5'UTR、内含子和外显子区T-DNA 插入、SNP 突变以及大片段置换改变基因序列(https://www. arabidopsis. org)。通过鉴定和应用这些突变体对AtTTG1基因功能和参与的信号途径进行了深入研究,如ttg1-1突变体由于单碱基突变导致转录提前终止,造成蛋白功能缺失。ttg1-1影响表皮细胞的分化、花青素和单宁的积累、植物昼夜节律变化、种皮粘液合成分泌和种皮分化等[20-22]。第一外显子由于T-DNA 插入缺失造成编码区序列改变的突变体ttg1-23以及AtTTG1基因的完全缺失突变体ttg1-24,两者均表现出种皮透明、种子呈现黄色、无毛状体分化以及不能合成花青素和单宁[23]。Wu 等[18]也利用拟南芥中7 个ttg1突变体证明了WD-40 domain 和C 端对AtTTG1 蛋白功能正常发挥的重要作用。ttg1-1突变体相比其他突变体蛋白缺失最少且只发生在C 末端,但是仍然导致AtTTG1 蛋白功能缺失。利用35S-SbTan1转化拟南芥ttg1-1突变体,大部分恢复了ttg1突变表型[18]。
蚕豆籽粒富含蛋白质(247~372 g·kg-1DM)和能量(15.6 MJ·kg-1DM),是一种高营养的豆类作物,传统上用于动物饲养和人类食用。尽管在农艺、营养和潜在的健康方面都有益处,但是由于种皮中富含单宁限制了蚕豆在饲料中的应用潜力。蚕豆籽粒无单宁表型受zt-1和zt-2两个互补隐性基因的控制,ZT-1(VfTTG1)基因编码位于2号染色体上的WD-40 domain 转录因子,与拟南芥中AtTTG1高度同源,参与单宁的合成调控途径。鉴定获得2 个VfTTG1基因的等位变异类型ttg1-a和ttg1-b。ttg1-a突变位点发生在启动子区,影响基因的表达以及转录的稳定性;ttg1-b蛋白序列N 端缺失。ttg1-a和ttg1-b蛋白功能丧失导致籽粒无单宁。设计了一套特异等位基因诊断标记,可以将zt-1与野生型和zt-2基因型区分开来,并将其应用于培育无单宁蚕豆的育种中[24]。玉米中与AtTTG1高度同源的基因ZmPAC1已被成功克隆,且发现了3 个不同的等位变异类型,分别为pac1-ref,pac1-2和pac1-3。ZmPAC1基因在花青素合成途径中发挥重要作用,过表达ZmPAC1基因转化拟南芥ttg1突变体可以部分恢复突变表型。ZmPAC1基因突变后影响幼苗根中花青素的积累,但是对根和毛状体的发育、分化没有影响,这与拟南芥ttg1突变体的表型差异很大[25]。高粱与玉米是亲缘关系最近的两个物种,且ZmPAC1与Tan1高度同源,但是生物学功能相差很大,Tan1是调控高粱单宁合成的必须基因,而玉米籽粒中无单宁,ZmPAC1不参与调控单宁的合成而参与花青素的积累。
本研究分析自然群体中单宁含量调控基因Tan1的不同等位基因型以及单宁含量发现,含有tan1-a、tan1-b和tan1-c突变等位基因型的高粱材料籽粒单宁含量均低于0.5%,但是含有野生型Tan1的高粱籽粒单宁含量从低到高都有分布。研究报道表明,调控高粱籽粒单宁含量的基因共有2个,即Tan1和Tan2,两个基因分别含有3 种等位变异基因型,其中任何一个基因发生变异都会导致高粱籽粒无单宁或者低单宁[19]。本研究中含有Tan1野生型且籽粒无单宁或者低单宁的高粱材料是否由于Tan2基因发生了变异还是存在其他的调控基因还需要进一步验证。
调控高粱籽粒单宁含量的基因可能远不止Tan1和Tan2,培育高单宁含量的品种时,需要同时检测多个单宁调控基因的基因型。然而对于培育低单宁含量的品种时,一是建议选择双亲都含有任意Tan1和Tan2突变等位基因型的材料进行杂交组配,后代个体均为低单宁或无单宁表型。二是当两亲本杂交时其中一个亲本含有Tan1和Tan2任何一种突变等位基因型时,在后代材料中选择含有突变等位基因型的个体。
4 结论
调控高粱籽粒单宁含量的Tan1基因共有4 种等位基因型,即Tan1、tan1-a、tan1-b和tan1-c,由于编码区序列的插入、缺失和突变使蛋白C 端序列发生改变,导致Tan1 蛋白功能丧失造成含有Tan1突变基因型的高粱籽粒无单宁或低单宁,而含有Tan1基因型的高粱材料单宁含量从低到高都有分布。通过对290 份栽培种高粱材料Tan1基因型和单宁含量的检测发现,在低单宁或无单宁品种培育中tan1-a和tan1-b应用广泛,而tan1-c应用较少。依据Tan1不同等位基因型的编码区序列差异,开发了一套实用的分子标记,为无单宁或低单宁杂交种培育提供快捷的分子检测手段。
猜你喜欢
作物学报(2022年2期)2022-11-06
中外葡萄与葡萄酒(2022年4期)2022-07-26
动物营养学报(2022年5期)2022-06-10
作物学报(2022年8期)2022-05-29
酿酒科技(2022年5期)2022-05-24
蔬菜(2021年7期)2021-11-27
新课程·下旬(2018年9期)2018-11-14
中国医药导报(2018年14期)2018-08-30
中老年健康(2016年1期)2016-03-07
青少年科技博览(中学版)(2015年10期)2015-01-11
