
不同类型水稻土可溶性有机氮及其组分的剖面分布①
2022-09-27 01:33鲍清涛杨文浩周碧青张黎明邢世和
土壤 2022年4期
杨 静,张 毅,鲍清涛,杨文浩,,周碧青,张黎明,,邢世和,*
不同类型水稻土可溶性有机氮及其组分的剖面分布①
杨 静1,张 毅1,鲍清涛1,杨文浩1,2,周碧青2,张黎明1,2,邢世和1,2*
(1 福建农林大学资源与环境学院,福州 350002;2 土壤生态系统健康与调控福建省高校重点实验室,福州 350002)
为揭示可溶性有机氮(soluble organic nitrogen,SON)在土壤剖面的分布状况,选取中亚热带地区发育于相同母质的黄泥田、灰黄泥田和灰泥田3种不同类型水稻土为对象,研究不同类型水稻土剖面中SON含量、组分及主控因子。结果表明,不同类型水稻土SON、游离氨基酸氮(FAA-N)、酰胺氮(AN-N)和可溶性蛋白氮(SP-N)含量具有明显的剖面分异,均表现为0 ~ 20 cm 土层>20 ~ 40 cm土层>40 ~ 60 cm土层。不同类型水稻土SON及各组分含量差异主要表现在0 ~ 20 cm土层,均表现为灰泥田>灰黄泥田>黄泥田,灰泥田SON、FAA-N、AN-N和SP-N含量分别高于灰黄泥田50.5%、41.7%、44.8% 和2.1%,高于黄泥田196.5%、200.9%、180.4% 和76.5%。0 ~ 20 cm土层3种不同类型水稻土FAA-N和AN-N分别占SON的54.4% ~ 58.7% 和45.5% ~ 48.1%,而底层(40 ~ 60 cm)FAA-N和AN-N分别占SON的33.1% ~ 55.7% 和50.3% ~ 52.8%,说明FAA-N和AN-N具有向下累积的趋势,且在土壤剖面中对SON的贡献大于SP-N。中性氨基酸尤其是肌氨酸是底层FAA-N的主要组分。灰色关联分析表明有机质、蛋白酶、脲酶和总孔隙度与不同类型水稻土SON及组分垂直分布密切相关,对SON、FAA-N、AN-N和SP-N含量变化的总贡献度分别为95.2%、92.1%、98.3% 和90.9%。因此,SON及组分在土壤剖面中的垂直分布与土壤类型和土层深度密切相关,且受土壤有机质、酶活和总孔隙度的影响。
水稻土;可溶性有机氮;组分;垂直分布;影响因素
土壤中90% 以上的氮素以有机形态存在,其中可被水或盐溶液(0.5 mol/L K2SO4、1 mol/L KCl和10 mmol/L CaCl2等)浸提出来的有机氮称为可溶性有机氮(soluble organic nitrogen,SON)[1]。SON在土壤中的移动性强,易受淋溶作用和胶体吸附作用的影响,具有明显的剖面部分特征,可为土壤的供氮能力和存在的生态环境影响提供参考[2]。土壤SON的化学组成十分复杂,不同组分自身性质存在较大差异,可能导致其在土壤剖面中的分布也具有一定的差异。研究表明亲水性小分子SON组分(如游离氨基酸、氨基糖和酰胺等)较大分子SON组分在土壤中更为活跃,不仅易被植物和微生物所吸收利用,还可随重力水的作用向下迁移累积[3]。因此,土壤剖面中SON的组成及存在形态对土壤氮素的有效性具有重要的影响。然而,由于分析技术的限制,目前关于SON在土壤剖面分布的研究主要集中在其含量方面,SON组分在土壤剖面的分异有待进一步研究。
不同土壤有其特定的理化性质,使得土壤微生物活性、有机质分解与释放发生相应的改变,故土壤SON含量和组分的分布也必然有所不同。姬景红等[4]研究表明草甸土和盐碱土20 ~ 40 cm土层SON含量较0 ~ 20 cm土层显著降低,而黑土则无显著差异。党亚爱等[5]研究表明黄土高原地区干湿砂质新成土、黄土正常新成土和土垫旱耕人为土有机氮各组分含量均表现为0 ~ 40 cm土层迅速下降,40 ~ 80 cm土层缓慢下降,80 cm以下土层基本稳定。段鹏鹏等[6]研究结果显示简育湿润淋溶土SON含量随土层深度加深呈逐渐下降趋势,且在20 ~ 40 cm处有明显的累积,而孔祥忠等[7]研究则表明土壤SON在40 ~ 60 cm有积累的趋势。可见,不同类型旱作土壤SON含量的剖面分异具有一定差异。水田土壤因频繁淹水耕作、干湿交替以及水稻根系泌氧特性等特异的环境条件可能导致SON的分布有别于旱地土壤,然而,国内外至今对不同类型水稻土SON组分剖面分布差异仍缺乏了解,且导致差异的原因尚不清晰。因此,本研究以亚热带3种不同类型水稻土为研究对象,分析不同类型水稻土SON及各组分的剖面分布差异,探讨产生差异的影响因素,为阐明水田生态系统SON的化学本质和生态功能提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于福建省闽侯县白沙镇溪头村(119°04'10"E,26°13'31"N),处于中亚热带季风气候区,年平均气温19.5 ℃,年无霜期311 d,夏季高温多雨,年均降水量可达1 350.9 mm,年日照时长1 812.5 h。供试土壤分别为黄泥田、灰黄泥田和灰泥田3种不同类型的水稻土,皆由中性岩红壤发育而来。研究区栽种的水稻品种为Ⅱ优673,属于再生稻,采用单季稻种植。试验中施用的化肥为尿素481.7 kg/hm2、过磷酸钙900 kg/hm2和氯化钾300 kg/hm2,尿素和氯化钾50% 作基肥,50% 作分蘖肥施用,过磷酸钙全部作基肥施用。田间管理和栽培措施均与当地保持一致。
1.2 土壤样品采集
土壤样品于2019年4月水稻种植前按照对角线多点取样法,利用不锈钢采土器分别采集不同类型及其不同深度(0 ~ 20、20 ~ 40、40 ~ 60 cm)的水稻土样品,同时采集各层次土壤容重圈样品。将采集的土壤样品放入聚乙烯自封袋中尽快带回实验室,将土壤样品中的根系、植物残体和侵入体等剔除,充分混匀后一部分新鲜样品置于4 ℃(短期)或–20 ℃(较长期)恒温冰箱存放,用于测定土壤SON、蛋白酶与脲酶等相关参数,另一部分置于室内常温下自然风干并研磨成不同粒径土样,供质地、有机质、pH、全氮和全碳等测定。容重圈样品用于土壤容重与毛管持水量测定,进而计算土壤孔隙度。
1.3 测定方法
土壤理化性质均采用土壤农化常规分析方法[8]。土壤脲酶和蛋白酶活性分别采用靛酚蓝比色法与福林酚比色法测定[9]。土壤SON含量采用安装有氮检测器总有机碳分析仪(Shimadzu,日本,TOC-L CPH)测定的土壤可溶性总氮(TSN)与采用连续流动注射分析仪(Systea,意大利,Flowsys)测定的土壤总无机氮(TIN)的差值获得[10]。土壤游离氨基酸含量和组成采用70 ℃热水浸提后,添加磺基水杨酸混合除蛋白,微调节pH至2.2后吸取1 ml滤液于氨基酸自动分析仪(Biochrom,英国,Biochrom 30+)测定[11];土壤酰胺态氮含量采用对二甲氨基苯甲醛比色法测定[12];土壤可溶性蛋白质采用70 ℃热水浸提,浸提后的溶液加入考马斯亮蓝充分混合后在595 nm 下比色测定[13];土壤可溶性组分采用70 ℃热水浸提,将浸提后的溶液通过冻干机冷冻干燥为粉末,并将1 mg冷冻干燥的样品与200 mg干燥的KBr磨细混匀,通过油压机压成均一透明的薄片,用傅里叶红外光谱仪(Bruker,德国,Vertex 70)测定[14]。
1.4 数据统计分析
数据采用Microsoft Excel 2010软件进行处理,利用SPSS 24软件对数据结果进行差异性分析检验,使用Sigmaplot 12.5和R 3.5.1软件进行作图处理。采用DPS7.05统计软件中的灰色关联分析模型探讨水稻土SON及各组分剖面分布差异的影响因素。
2 结果与分析
2.1 不同类型水稻土理化性质及其剖面差异
从表1可知,除灰泥田表层(0 ~ 20 cm)土壤为黏土外,不同类型不同层次土壤均为壤质黏土。不同类型水稻土的容重表现为黄泥田>灰黄泥田>灰泥田,且随深度的增加而增大。不同类型水稻土表层土壤总孔隙度无显著差异,且均显著高于深层土壤。不同类型水稻土的pH和C/N均随深度的增加而增大,其中以黄泥田表层土壤pH和C/N最大,而灰黄泥田pH和C/N最小。不同类型水稻土有机质、脲酶和蛋白酶活性均随土壤深度的增加而降低,且黄泥田有机质、脲酶和蛋白酶活性均低于灰泥田和灰黄泥田。可见,不同类型水稻土理化性质差异显著,表层土壤理化性质与深层间的差异显著。
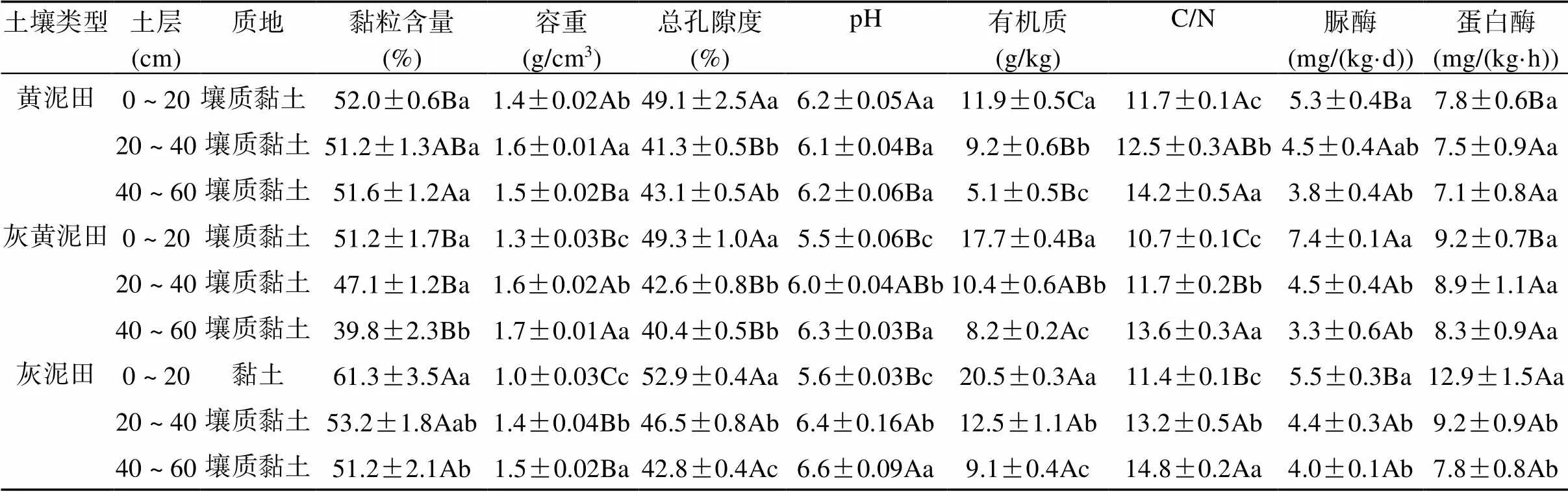
表1 不同类型水稻土不同土层理化性质差异
注:表中数值为平均值±标准误,大写字母不同表示同一土层不同土壤间差异显著(<0.05),小写字母不同表示同一土壤不同土层间差异显著(<0.05),下表同。
2.2 不同类型水稻土SON含量及其剖面差异
水稻土不同土层SON含量具有一定差异,灰黄泥田和灰泥田SON含量均表现为0 ~ 20 cm>20 ~ 40 cm>40 ~ 60 cm,而黄泥田0 ~ 20 cm土层SON含量与20 ~ 40 cm土层间无显著差异(图1)。不同类型水稻土SON含量差异显著,0 ~ 20 cm土层灰泥田水稻土SON含量显著高于灰黄泥田和灰泥田,但深层土壤(20 ~ 40 cm和40 ~ 60 cm)不同类型水稻土SON含量无显著差异。可见,不同类型水稻土SON含量的剖面分异较为明显,灰泥田表层SON含量较深层土壤的差异最大,灰黄泥田次之,黄泥田最小。
2.3 不同类型水稻土SON组成及其剖面差异
2.3.1 不同类型水稻土游离氨基酸氮含量、组成及其剖面差异 不同类型水稻土游离氨基酸氮(FAA-N)含量随土壤深度增加呈递减的变化趋势,但20 ~ 40 cm和40 ~ 60 cm土层间FAA-N含量无显著差异(图2)。不同类型水稻土FAA-N含量具有一定差异,灰黄泥田和灰泥田0 ~ 20 cm土层FAA-N含量显著高于黄泥田,而40 ~ 60 cm土层黄泥田FAA-N含量则显著高于灰黄泥田和灰泥田。不同类型水稻土不同土层FAA-N/SON介于33.8% ~ 58.7%(表2),灰黄泥田和灰泥田FAA-N随土壤深度增加呈递减,而黄泥田3个土层间FAA-N/SON无显著差异。可见,FAA-N是水稻土SON的重要组成成分,且不同类型水稻土FAA-N差异主要表现在0 ~ 20 cm土层;不同类型水稻土FAA-N存在向下累积现象,且黄泥田中FAA-N向下累积的趋势更为明显。

(大写字母不同表示同一土层不同土壤间差异显著(P<0.05),小写字母不同表示同一土壤不同土层间差异显著(P<0.05),下图同)
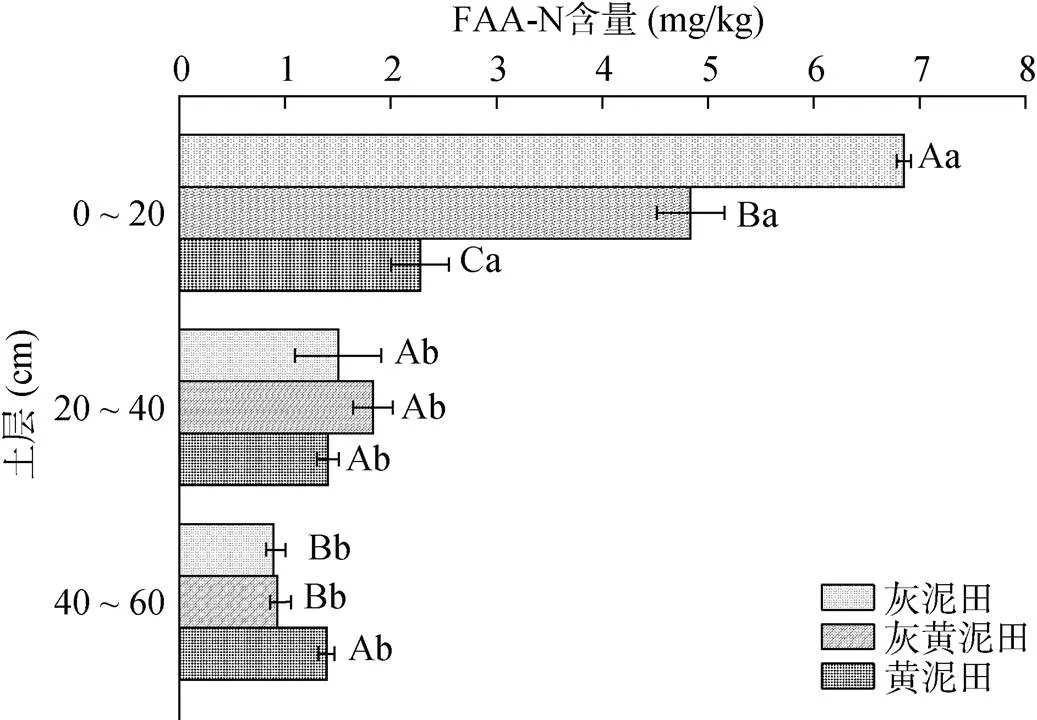
图2 不同类型水稻土不同土层FAA-N含量
不同类型水稻土游离氨基酸(FAA)种类及含量均随土壤深度的增加呈递减(图3),0 ~ 20 cm土层不同类型水稻土FAA均以肌氨酸、丙氨酸、缬氨酸和亮氨酸为主,而20 ~ 40 cm和40 ~ 60 cm土层中FAA均以肌氨酸含量相对较高。黄泥田3个土层间氨基酸的组成差异小于灰黄泥田和灰泥田(图4),0 ~ 20 cm土层肌氨酸含量表现为黄泥田<灰黄泥田、灰泥田,而20 ~ 40 cm和40 ~ 60 cm土层肌氨酸含量均表现为黄泥田>灰黄泥田、灰泥田。可见,不同类型水稻土FAA的组成均以中性氨基酸为主,黄泥田中FAA组成的剖面分异较小,且在土壤剖面分布中肌氨酸的差异最小。

表2 不同类型水稻土不同土层SON组分所占比例(%)
注:SON:可溶性有机氮;FAA-N:游离氨基酸氮;AN-N:酰胺氮;SP-N:可溶性蛋白氮。
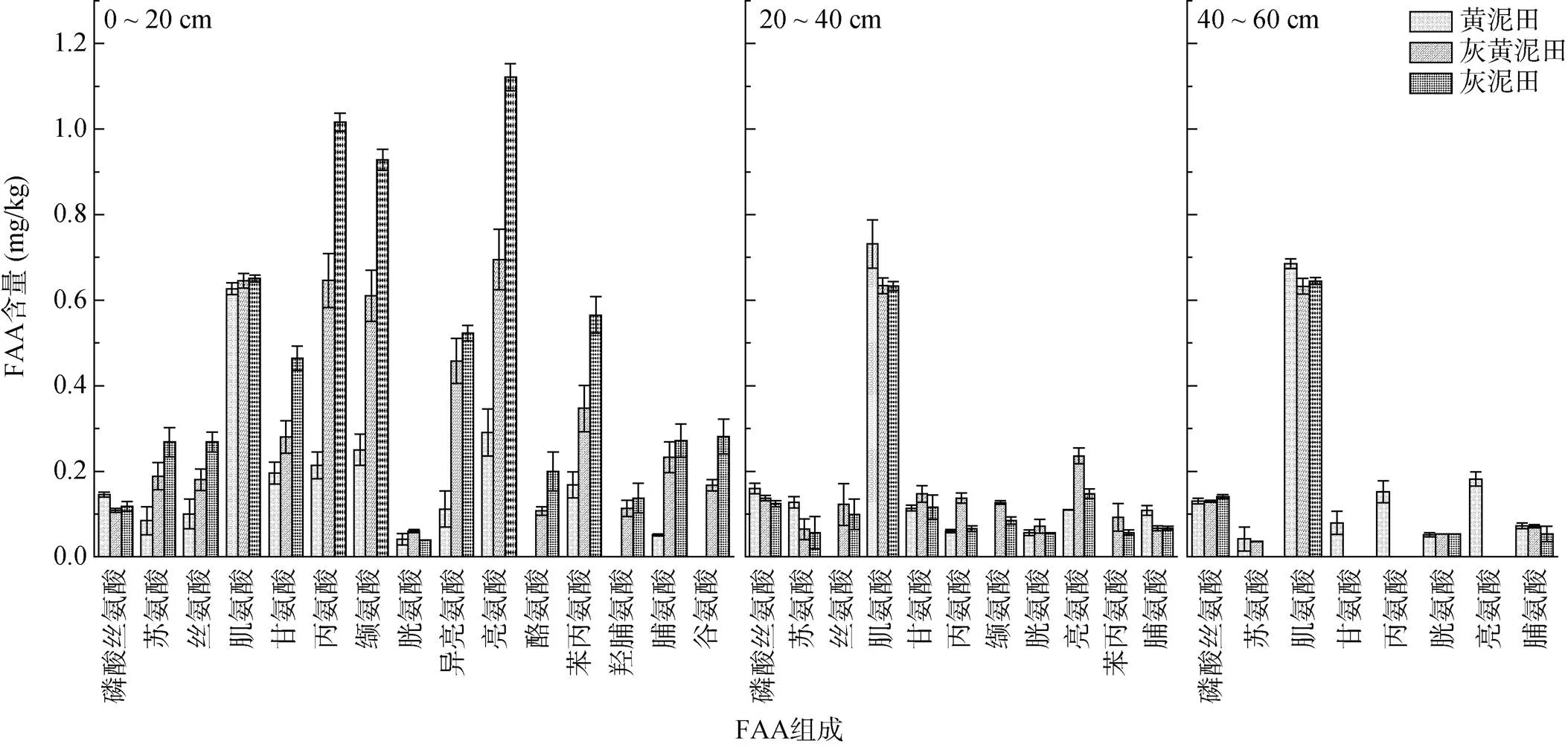
图3 不同类型水稻土不同土层FAA组成

(图中A、B和C分别表示黄泥田、灰黄泥田和灰泥田;20、40和60分别表示0 ~ 20、20 ~ 40和40 ~ 60 cm土层)
2.3.2 不同类型水稻土酰胺氮含量及其剖面差异 灰黄泥田和灰泥田0 ~ 20 cm土层酰胺氮(AN-N)含量显著高于20 ~ 40 cm和40 ~ 60 cm土层,而黄泥田不同土层间AN-N含量无显著差异(图5)。不同类型水稻土AN-N含量也具有一定差异,0 ~ 20 cm土层AN-N含量表现为灰泥田>灰黄泥田>黄泥田,而20 ~ 40 cm和40 ~ 60 cm土层不同类型水稻土AN-N含量无显著差异。不同类型水稻土不同土层AN-N/SON介于45.0% ~ 55.2%,不同类型水稻土间和不同土层间AN-N/SON均无显著差异(表2)。可见,AN-N是SON的重要组成成分,且不同类型水稻土AN-N差异主要表现在0 ~ 20 cm土层;不同类型水稻土AN-N存在向下累积的现象,且黄泥田中AN-N向下累积的趋势更为明显。
2.3.3 不同类型水稻土可溶性蛋白氮含量及其剖面差异 不同类型水稻土0 ~ 20 cm土层可溶性蛋白氮(SP-N)含量显著高于20 ~ 40 cm和40 ~ 60 cm土层,而20 ~ 40 cm和40 ~ 60 cm土层间SP-N含量无显著差异(图6)。不同类型水稻土SP-N含量也具有一定差异,0 ~ 20 cm土层中灰黄泥田和灰泥田SP-N含量显著高于黄泥田,而20 ~ 40 cm和40 ~ 60 cm土层不同类型土壤SP-N含量无显著差异。不同类型水稻土不同土层SP-N/SON介于16.0% ~ 38.7%,3种类型水稻土SP-N/SON均表现为0 ~ 20 cm>20 ~ 40 cm≈40 ~ 60 cm(表2);0 ~ 20 cm土层灰泥田SP-N/SON显著低于黄泥田和灰黄泥田,20 ~ 40 cm和40 ~ 60 cm土层3种类型水稻土SP-N/SON无显著差异。可见,SP-N是SON的重要组成成分,不同类型水稻土SP-N剖面分异较大,且均主要表现在表层土壤。
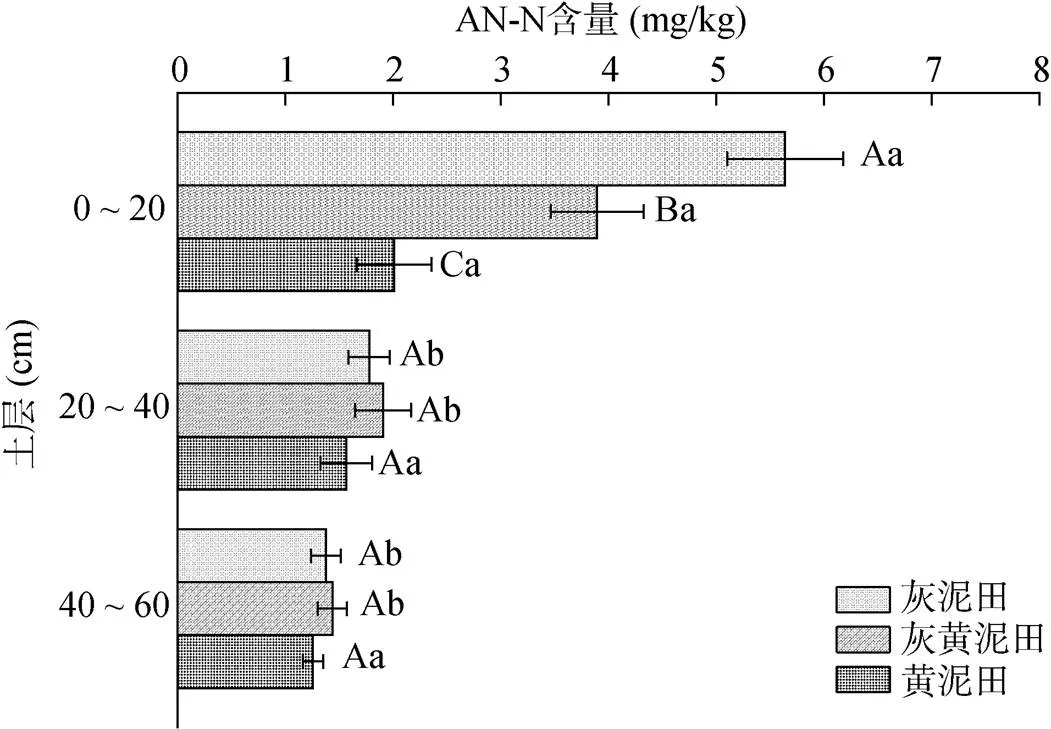
图5 不同类型水稻土不同土层AN-N含量
2.3.4 不同类型水稻土可溶性组分的剖面差异 不同类型水稻土不同土层可溶性组分浸提液的红外光谱均存在7个明显的吸收峰(图7)。就不同类型水稻土而言,0 ~ 20 cm土层各吸收峰的峰面积均表现为灰泥田>灰黄泥田>黄泥田的变化趋势;20 ~ 40 cm土层峰1和峰4的峰面积表现为黄泥田高于灰黄泥田和灰泥田;而40 ~ 60 cm土层各吸收峰的峰面积均表现为黄泥田高于灰黄泥田和灰泥田,表明黄泥田底层土壤中SON累积量大于灰黄泥田和灰泥田。就不同土层而言,黄泥田峰1和峰4的峰面积均表现为20 ~ 40 cm>40 ~ 60 cm>0 ~ 20 cm,而灰黄泥田和灰泥田则均表现为0 ~ 20 cm>20 ~ 40 cm>40 ~ 60 cm;不同类型水稻土峰2的峰面积均表现为0 ~ 20 cm>20 ~ 40 cm>40 ~ 60 cm,而其他吸收峰的峰面积无明显的变化规律。可见,大分子可溶性氮主要分布在水稻表层土壤中,而小分子可溶性氮具有在水稻土底层累积的趋势,且黄泥田的累积量最大。

图6 不同类型水稻土不同土层SP-N含量
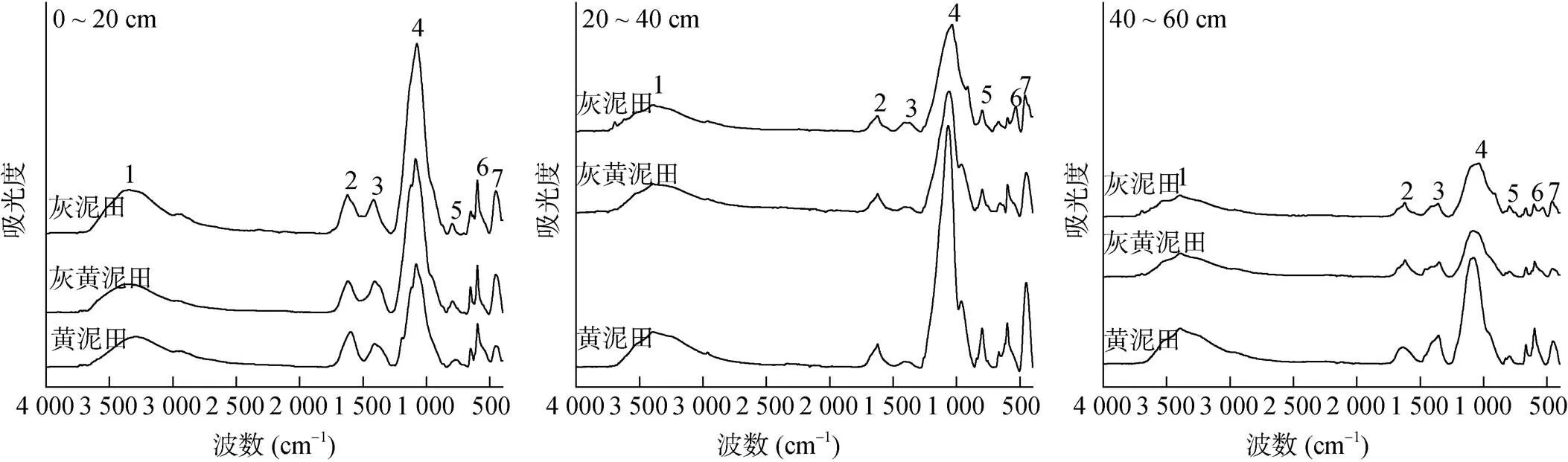
(吸收峰1位于3 400 cm–1左右,属于碳水化合物、羧酸、酚类、醇类等的–OH伸缩振动,以及酰胺类官能团的N-H伸缩振动;吸收峰2位于1 630 cm–1左右,木质素中与芳香环相连的C=O伸缩振动及酰胺类化合物C=O伸缩振动,即酰胺I吸收带;吸收峰3位于1 400 cm–1左右,木质素、脂肪族化合物及C-N伸缩振动,即酰胺III吸收带;吸收峰4位于1 086 cm–1左右,属于碳水化合物(纤维素、半纤维素)和多糖的 C-O 键的伸缩振动吸收;吸收峰5位于795 cm–1左右,属于碳酸根形成的吸收峰;吸收峰6位于585 ~ 655 cm–1左右,羟基的变角振动;吸收峰7位于453 cm–1左右,属于Si-O振动吸收)
2.4 不同类型水稻土SON及其组分差异的主要影响因素和贡献率
为探究不同影响因子对土壤SON及其组分的影响大小,采用灰色关联分析模型计算了水稻土SON及其组分与可能影响因子之间的关联程度(图8)。就水稻土SON含量影响因素而言,土壤有机质、蛋白酶和脲酶与SON的关联系数较大,均高于0.90,其次为总孔隙度,关联系数为0.85。从水稻土SON主要组分的影响因素来看,土壤有机质与FAA-N、AN-N和SP-N的关联度均为最高,关联系数分别为0.89、0.94和0.91,其次为总孔隙度、脲酶和蛋白酶,关联系数介于0.80 ~ 0.88。可见,有机质、总孔隙度、蛋白酶和脲酶活性与水稻土SON含量及组分差异密切相关。

图8 水稻土SON及其组分与影响因素间的灰色关联系数热图
采用方差分解分析进一步探究水稻土SON主要组分的4个主要影响因素对其含量变异的贡献率大小。结果(图9)表明,土壤有机质、总孔隙度和酶活性(脲酶和蛋白酶)对SON含量变异的贡献率分别为0.03%、0.9% 和8.3%;对FAA-N含量变异的贡献率分别为1.8%、1.1% 和8.6%;对AN-N含量变异的贡献率分别为0.2%、0.6% 和5.5%;对SP-N含量变异的贡献率分别为2.7%、0.8% 和2.8%。4个主效因子的交互作用对水稻土SON、FAA-N、AN-N和SP-N含量变异的贡献率分别达68.7%、71.0%、70.7% 和74.4%。各变量及交互作用对SON、FAA-N、AN-N和SP-N含量变异的总贡献率分别达95.2%、92.1%、98.3% 和90.9%。可见,土壤有机质、脲酶活性、蛋白酶活性和总孔隙度的协同作用对土壤SON各组分差异的贡献率较大。

(A代表有机质,B代表总孔隙度,C代表脲酶活性和蛋白酶活性,D代表未解释量;AB、BC和AC分别代表有机质和总孔隙度、总孔隙度和酶活性、有机质和酶活性的协同作用;ABC代表有机质、总孔隙度和酶活性的协同作用)
3 讨论
3.1 不同类型水稻土SON含量的剖面分异
土壤SON具有较强的迁移性,过量施肥会导致其在土壤剖面中垂直迁移累积,造成氮素损失。本研究结果表明不同类型水稻土SON含量剖面分异较为明显,SON随土层深度的增加逐渐下降,且不同类型水稻土SON含量差异主要表现在0 ~ 20 cm土层,这与已有的结论相似[15]。究其原因与耕层土壤中有机质的累积密切相关,施入土壤的肥料和动植物残体均在表层积累[16],致使土壤中的有机质多分布在土壤表层,其含量随土壤深度的增加而减少(表1),且由于长期人为施肥使表层土壤中营养物质富集,有利于提高土壤酶活性,本研究表明3种不同类型水稻土表层土壤中脲酶和蛋白酶活性均显著高于20 ~ 40 cm、40 ~ 60 cm土层,有利于表层土壤有机质矿化释放出SON;此外,由于供试水稻土的质地较为黏重(均为黏土和壤质黏土),孔隙度随土层的增加而降低(表1),加上紧实犁底层的滞水作用,导致水稻土SON不容易随水下渗;第三,水稻根系分泌物也是SON的来源之一,其主要分布于土壤表层,且根系分泌物还可促进微生物繁殖,提高土壤酶活性,进而促进表层土壤有机氮分解和矿化产生SON[17];致使供试水稻土SON含量均随土壤深度增加而减少,且表层土壤SON的浓度显著高于20 ~ 40 cm和40 ~ 60 cm土层。
土壤类型及其颗粒组成决定着土壤的含水量,在一定程度上影响土壤通气性和微生物活性,是影响土壤中氮化合物转化和迁移的内因[18]。不同类型水稻土SON的剖面分布具有一定差异,灰泥田表层SON含量较深层土壤的差异最大,灰黄泥田次之,黄泥田最小,这可能与不同类型土壤的有机质含量、酶活性和孔隙度不同有关。不同土壤类型由于熟化程度不同,导致其耕层有机质含量具有一定差异,灰泥田耕层土壤有机质是灰黄泥田和黄泥田的1.16倍和1.73倍,增加了其耕层SON的来源。此外,SON在土壤中移动性和有效性受土壤吸附过程的影响,而有机质对SON的吸附可降低其在土壤中的迁移[19],且灰泥田黏粒含量高(61.32%),具有较大的比表面积与电荷密度[20],能有效吸附土壤中的SON,减少SON的迁移量,从而导致灰泥田表层SON含量显著高于深层。土壤酶是土壤中有机物分解和氮转化的主要驱动因素,有机物料施入土壤腐解过程中能够释放大分子量的复杂有机聚合物,在酶的解聚作用下可释放大量小分子有机氮(如氨基酸、酰胺、氨基糖等)[21]。本研究中供试的3种不同类型水稻土中灰泥田不同土层间脲酶和蛋白酶活性差异最大,对有机质分解能力的不同也导致灰泥田中SON垂直分布的差异较大。土壤孔隙度大小决定了土壤的通气性,是影响土壤水分运动和溶质迁移的重要因素[22]。李胜龙[23]研究表明影响氮素迁移的因素较多,其中土壤孔隙具有重要的作用。本研究灰色关联分析结果也表明土壤孔隙度与土壤SON垂直分布密切相关(图8)。孔隙度大,保肥能力差,孔隙小,氮素下渗速度慢[24]。本研究供试黄泥田中通气孔隙度是灰黄泥田和灰泥田的2.4倍和1.3倍,通气孔隙虽在一定程度上能增加土壤透气性,但很大程度上造成大量的溶质迁移[22],从而导致黄泥田中SON向下累积,垂直分布差异较小。
3.2 不同类型水稻土SON组分的剖面分异
不同类型水稻土AN-N、FAA-N和SP-N分别占SON的45.0% ~ 55.2%、33.8% ~ 58.7% 和16.0% ~ 38.7%,说明AN-N、FAA-N和SP-N是SON的重要组成成分,与Murphy等[1]研究结果一致。AN-N、FAA-N和SP-N均随土壤剖面深度的增加逐渐下降,且不同类型土壤间的差异主要表现在0 ~ 20 cm土层,与SON含量变化规律一致。不同SON组分在土层间的迁移能力不同,在土壤剖面中的分布也具有一定的差异。本研究结果表明AN-N和FAA-N具有向下累积的趋势,且底层土壤中AN-N和FAA-N对SON的贡献大于SP-N,这主要与SON组分自身性质及土壤对其的吸附和固持作用差异有关。研究表明小分子的亲水性SON不易被土壤吸附,吸附后生物稳定性差,还易出现解吸或被疏水性SON代替吸附的现象,因此易在土壤中发生迁移[24]。Michalzik和Matzner[25]通过核磁共振研究也表明氨基酸和酰胺态氮是森林土壤滤出液SON的重要组成成分。而SP由氨基酸脱水缩合而成的多条肽链所组合而成[26],分子量显著大于AN-N和FAA-N,结构也较为复杂,导致其在土壤中迁移能力较弱。本研究傅里叶红外分析结果也表明大分子复杂SON主要集中于表层土壤,而酰胺等小分子SON在底层中的累积量较大(图7)。
供试3种类型水稻土中表层土壤氨基酸均以中性氨基酸含量最高,酸性氨基酸次之,碱性氨基酸含量最低,这与雷秀美等[15]研究长期不同施肥对土壤游离氨基酸影响的结果类似。本研究的试验地位于亚热带,供试水稻土呈酸性反应(pH为5.5 ~ 6.6),酸性条件下碱性氨基酸的化学稳定性较低,容易发生降解,故供试土壤中未检出碱性氨基酸[27]。本研究中深层土壤中仅有中性氨基酸,说明中性氨基酸在不同类型水稻土中较为稳定,是不同类型水稻土剖面中迁移的主要氨基酸组分,这可能与氨基酸的带电性有关。酸性氨基酸(谷氨酸)等电点为3.2,在土壤溶液中带负电荷,带有负电荷的羧基可通过二价离子作为离子桥与土壤基质相结合而被吸附,从而导致其迁移性较弱;而中性氨基酸等电点与土壤溶液较为接近,基本以偶极的形态存在,带正电荷的基团和带负电荷的基团距离太近,电荷相互抵消[28]。因此土壤对中性氨基酸的吸附能力较低,易迁移至深层。Rothstein等[29]研究也表明酸性土壤可同时吸附带正、负电荷的游离氨基酸,导致土壤对酸性和碱性氨基酸的吸附较中性氨基酸强烈。就单个氨基酸组分而言,3种不同土壤类型水稻土不同土层中肌氨酸的含量均较高,且无显著性差异,这可能是因为肌氨酸作为中性氨基酸不易被土壤胶体吸附,在土壤剖面中移动性强,从而导致不同土层中肌氨酸含量无显著差异。此外,氨基酸在剖面的分布与其性质密切相关,极性氨基酸的矿化程度高于非极性氨基酸[30],因此供试土壤中肌氨酸作为非极性氨基酸矿化程度较低,周转速率较慢,从而成为土壤剖面游离氨基酸含量较高的成分之一。不同类型水稻土游离氨基酸的垂直分布具有一定的差异,黄泥田底层氨基酸的种类较多且含量较高,说明游离氨基酸易在黄泥田底层土壤中累积,可能是SON流失的重要形态,这与施书莲等[30]研究的土壤剖面不同粒级中氨基酸含量的结果一致。土壤颗粒粒径含量组成影响土壤剖面中氮素分布特征、累积强度或淋失强度,本研究供试黄泥田砂粒含量相对较高,保水能力弱,利于游离氨基酸随水分向下迁移。
4 结论
不同类型水稻土SON及各组分含量剖面分异明显,均随土层的加深而降低,且主要受土壤有机质、蛋白酶、脲酶和总孔隙度的交互影响。土壤肥力对SON组分在土壤剖面中的分布有显著影响,FAA-N和AN-N是水稻土底层土壤SON累积的主要形态,且肌氨酸是游离氨基酸累积的主要组分。低肥力的供试黄泥田SON在底层累积的趋势更为明显,易产生损失,田间管理中应特别注意水肥管理,降低其产生的环境风险。
[1] Murphy D V, Macdonald A J, Stockdale E A, et al. Soluble organic nitrogen in agricultural soils[J]. Biology and Fertility of Soils, 2000, 30(5/6): 374–387.
[2] 储成, 程谊, 曹亚澄, 等. 土壤可溶性有机氮测定方法研究进展[J]. 土壤, 2021, 53(3): 449–457.
[3] 张亚亚, 李军, 郭颖, 等. 土壤可溶性有机氮的研究进展[J]. 生态学杂志, 2016, 35(6): 1661–1669.
[4] 姬景红, 刘双全, 李玉影, 等. 黑龙江省不同类型旱田土壤有机氮组分含量及分布[J]. 黑龙江农业科学, 2020(9): 38–41.
[5] 党亚爱, 王国栋, 李世清. 黄土高原典型土壤有机氮组分剖面分布的变化特征[J]. 中国农业科学, 2011, 44(24): 5021–5030.
[6] 段鹏鹏, 丛耀辉, 徐文静, 等. 氮肥与有机肥配施对设施土壤可溶性氮动态变化的影响[J]. 中国农业科学, 2015, 48(23): 4717–4727.
[7] 孔祥忠, 于红梅, 束良佐, 等. 农田利用方式对土壤可溶性氮素含量及迁移的影响[J]. 土壤通报, 2015, 46(6): 1359–1365.
[8] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000.
[9] 关松荫. 土壤酶及其研究法[M]. 北京: 农业出版社, 1986.
[10] Chen C R, Xu Z H, Zhang S L, et al. Soluble organic nitrogen pools in forest soils of subtropical Australia[J]. Plant and Soil, 2005, 277(1/2): 285–297.
[11] 杨静, 聂三安, 杨文浩, 等. 不同施肥水稻土可溶性有机氮组分差异及影响因素[J]. 土壤学报, 2018, 55(4): 955–966.
[12] 周佳, 孙勇, 唐传球, 等. 对二甲氨基苯甲醛比色法测定溶液中的尿素[J]. 化学与生物工程, 2014, 31(2): 75–78.
[13] Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J]. Analytical Biochemistry, 1976, 72(1/2): 248–254.
[14] 宋祥云, 宋春燕, 柳新伟, 等. 小麦玉米轮作条件下不同生物质炭对土壤腐殖物质的影响[J]. 土壤学报, 2021, 58(3): 610–618.
[15] 雷秀美, 王飞, 周碧青, 等. 长期施肥对稻田土壤可溶性有机氮和游离氨基酸剖面分异的影响[J]. 农业环境科学学报, 2019, 38(7): 1550–1559.
[16] 李孝刚, 彭曙光, 靳志丽, 等. 有机物料对植烟土壤氮素矿化及微生物性质的影响[J]. 土壤学报, 2021, 58(1): 225–234.
[17] 颜志雷, 方宇, 陈济琛, 等. 连年翻压紫云英对稻田土壤养分和微生物学特性的影响[J]. 植物营养与肥料学报, 2014, 20(5): 1151–1160.
[18] 李京京, 刘文, 任天宝, 等. 不同土壤质地和含水率对炭基肥料氮素矿化的影响[J]. 土壤, 2020, 52(1): 40–46.
[19] 赵满兴, 周建斌, 陈竹君, 等. 不同类型农田土壤对可溶性有机氮、碳的吸附特性[J]. 应用生态学报, 2008, 19(1): 76–80.
[20] 王红霞. 土壤中溶解性有机碳、氮及其迁移淋溶特性研究[D]. 杨凌: 西北农林科技大学, 2008.
[21] Cenini V L, Fornara D A, McMullan G, et al. Linkages between extracellular enzyme activities and the carbon and nitrogen content of grassland soils[J]. Soil Biology and Biochemistry, 2016, 96: 198–206.
[22] 刘俊廷, 张建军, 孙若修, 等. 晋西黄土区退耕年限对土壤孔隙度等物理性质的影响[J]. 北京林业大学学报, 2020, 42(1): 94–103.
[23] 李胜龙. 稻田—田埂过渡区土壤大孔隙分布与水氮渗漏特征[D]. 武汉: 华中师范大学, 2018.
[24] 黄倩, 吴靖霆, 陈杰, 等. 土壤吸附可溶性有机碳研究进展[J]. 土壤, 2015, 47(3): 446–452.
[25] Michalzik B, Matzner E. Dynamics of dissolved organic nitrogen and carbon in a Central European Norway spruce ecosystem[J]. European Journal of Soil Science, 1999, 50(4): 579–590.
[26] 冯艳芳. 氨基酸保护及肽键合成的研究[D]. 太原: 山西大学, 2011.
[27] 王星, 崔晓阳, 郭亚芬. 寒温带林区不同林型土壤中游离氨基酸的研究[J]. 南京林业大学学报(自然科学版), 2016, 40(4): 42–48.
[28] 张旭东, 汪景宽, 张继宏, 等. 长期施肥对土壤中氨基酸吸附特性的影响迈向. 21世纪的土壤科学—提高土壤质量促进农业持续发展[C]. 南京: 中国土壤学会, 1999: 53–57.
[29] Rothstein D E. Effects of amino-acid chemistry and soil properties on the behavior of free amino acids in acidic forest soils[J]. Soil Biology and Biochemistry, 2010, 42(10): 1743–1750.
[30] 施书莲, 周克瑜, 杨文醒. 土壤剖面不同粒级中氨基酸组成特征[J]. 土壤, 1998, 30(4): 209–213.
Profile Distribution of Soluble Organic Nitrogen and Its Components in Different Paddy Soils
YANG Jing1, ZHANG Yi1, BAO Qingtao1, YANG Wenhao1,2, ZHOU Biqing2, ZHANG Liming1,2, XING Shihe1, 2*
(1 College of Resources and Environment, Fujian Agriculture and Forestry University, Fuzhou 350002, China; 2 Fujian Provincial Key Laboratory of Soil Environmental Health and Regulation, Fuzhou 350002, China)
In order to reveal the distribution of soluble organic nitrogen (SON) in soil profile, three types of paddy soils (yellow, grey and yellow podzolic paddy soils) with the same parent material were selected to study the content, composition and main influencing factors of SON in paddy soil profiles. The results showed that the contents of SON, free amino acid nitrogen (FAA-N), amide nitrogen (AN-N) and soluble protein nitrogen (SP-N) had significant differences in different paddy soil profiles, and all were in an order of 0-20 cm > 20-40 cm > 40-60 cm. The contents of SON and its components in different paddy soils were mainly manifested in the 0-20 cm, in which all was in an order of grey paddy soil > yellow podzolic paddy soil > yellow paddy soil. The contents of SON, FAA-N, AN-N and SP-N in grey paddy soil were 50.5%, 41.7%, 44.8% and 2.1% higher than those of yellow podzolic paddy soil, 196.5%, 200.9%, 180.4% and 76.5% higher than those of yellow paddy soil. The contents of FAA-N and AN-N of the three paddy soils in 0–20 cm accounted for 55.4%–58.7% and 45.5%–48.1% of SON, respectively, while the contents of FAA-N and AN-N in 40–60 cm accounted for 33.1%–55.7% and 50.3%–52.8% of SON, respectively. These results indicated that FAA-N and AN-N had a downward accumulation trend, and contribute more to SON than SP-N in soil profile. Neutral amino acids, especially sarcosine, were the main components of FAA-N in 40–60 cm. Grey correlation analysis showed that organic matter, protease, urease and total porosity were closely related to the vertical distribution of SON and components in different paddy soils, and their contribution to SON, FAA-N, AN-N and SP-N were 95.2%, 92.1%, 98.3% and 90.9%, respectively. Therefore, the vertical distribution of SON and its components in soil profile is closely related to soil type and depth, and is affected by soil organic matter, enzyme activities and total porosity.
Paddy soil; Soluble organic nitrogen; Composition; Vertical distribution; Influencing factors
S154
A
10.13758/j.cnki.tr.2022.04.005
杨静, 张毅, 鲍清涛, 等. 不同类型水稻土可溶性有机氮及其组分的剖面分布. 土壤, 2022, 54(4): 691–699.
国家自然科学基金面上项目(41671490)、福建农林大学科技创新项目(CXZX2017217)和福建农林大学优秀博士学位论文资助基金(324-1122yb080)资助。
(fafuxsh@126.com)
杨静(1992—),女,福建南平人,博士研究生,主要从事土壤可溶性有机氮研究。E-mail: yangjing_0305@163.com
猜你喜欢
中国科技纵横(2022年17期)2022-10-25
建材发展导向(2022年10期)2022-07-28
作文大王·低年级(2022年3期)2022-03-19
土壤学报(2021年1期)2021-04-08
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
电子技术与软件工程(2019年6期)2019-12-02
广东第二课堂·小学(2019年1期)2019-03-06
新作文·小学低年级版(2016年3期)2017-06-01
