
双齿围沙蚕对潮汐更替的行为响应
2022-09-25 09:46:16何为房景辉梁博蒋增杰毛玉泽方建光姜娓娓高亚平
海洋学报 2022年10期
何为,房景辉*,梁博,蒋增杰,毛玉泽,方建光,姜娓娓,高亚平
( 1. 上海海洋大学 水产与生命学院,上海 201306;2. 中国水产科学研究院黄海水产研究所 海洋渔业科学与食物产出过程功能实验室,山东 青岛 266071)
1 引言
水生动物的摄食、生长发育、繁殖等生命活动与水体环境条件紧密相关[1-4]。海洋潮汐作为普遍现象会对潮间带环境产生显著影响。有研究发现,潮汐会改变水下沉积物的层理结构和悬浮泥沙的含量[5-6],进而影响生物的栖息与生存。潮间带生物在潮汐更替过程中会经历退潮干露和涨潮水体淹没等过程。底内动物作为潮间带优势生物类群之一,对潮汐变化有独特的应对策略。Koo等[7]在研究潮滩沉积物-水界面时发现,潮位越高,底内动物建造洞穴的尺寸越大,洞穴越深。在大潮时,双齿围沙蚕(Perinereis aibuhitensisGrube)对沉积颗粒的修复率比小潮阶段更高[8]。在潮汐过程中,除了改变栖息空间和栖息底质,潮间带底内动物可能存在其他的适应性行为。杂色刺沙蚕(Nereis diversicolor)在退潮后的2 h内先滤食后进行表面摄食,但是在潮滩表面暴露的总时间较短[9]。底内动物在沉积物中进行运动和掘洞时会产生泵水作用和水流效应,这一过程对沉积物中的生地化过程和水中的溶解氧交换有着重要作用[10]。潮汐更替导致强烈的水体交换,底内动物的行为在不同的潮汐阶段可能存在差异。底内动物对潮汐变化的响应过程对于我们了解沉积物-水界面的生地化过程有着很大的帮助,但是关于这一过程的研究较为缺乏。
双齿围沙蚕作为河口和沿海潮间带广泛分布的底内动物之一,在水产养殖中有着重要的经济价值并且对生态环境有积极的修复作用[11]。当水中溶解氧含量低于某一数值时,双齿围沙蚕会主动进行泵水来抵御低氧带来的威胁[12],因此其在潮汐更替过程中可能存在明显的行为变化。本研究以双齿围沙蚕为研究对象,研究了其在不同潮汐阶段和温度下的行为规律,以期探究潮汐更替和水温的协同作用对双齿围沙蚕行为的影响。本研究可以明晰双齿围沙蚕等底内动物对潮汐更替的行为适应策略,为底内动物行为生态学研究提供重要参考。
2 材料与方法
2.1 实验动物与暂养
实验所用的沉积物均从双齿围沙蚕自然栖息地取得,将沉积物置于海水循环养殖池中沉降稳定1周。从威海乳山的沙蚕养殖场采集了双齿围沙蚕,规格为(2.5±0.2)g,在海水循环养殖池中暂养1周。暂养期间,不断挑出活力不佳的个体,水体条件维持在水温为20℃,盐度为29~31,溶解氧浓度为7.0~8.5 mg/L,每两天投喂一次对虾饲料,并且每天换水30%。本实验设置了15℃、20℃和25℃ 3个温度梯度,并通过冷水机(深圳,日升CL-650)控温。将暂养池中的双齿围沙蚕分为3份,转入提前控温为20℃的3个循环水系统中,其中提前1周铺设了和暂养池一样的沉积物。为了使水族箱中水体的温度满足实验要求,在20℃的基础上,每天升高或者降低水族箱内水温1℃,一直到25℃或15℃。让箱中的双齿围沙蚕再适应3 d后才开展实验。
2.2 实验过程
实验将潮汐过程分成4个阶段,即T1:退潮前(沉积物上方存在上覆水超过3 h)、T2:退潮后(沉积物无上覆水时)、T3:涨潮30 min内(从开始添加上覆水到之后的30 min内)和T4:涨潮30 min后(添加上覆水30 min后),每个潮汐阶段对应一个时间段。由于双齿围沙蚕在干露状态时,无径向起伏和泵水过程,因此退潮后的径向起伏和泵水过程不用摄像装置录制。
在底内动物行为学观测装置中的小正方体容器中注入沉积物,往装置的3层空间通入不同温度的海水。在沉积物沉降之后,提前1 d,傍晚约17:00,将一条活力较佳的沙蚕放于小隔间中,让其自然掘洞栖息。当双齿围沙蚕建好洞穴并且其运动状态易于观察时,开始录制。录制1.5 h时,放掉装置第一层的海水,待沙蚕干露3 h后,往装置第一层中通入海水,再录制1.5 h,关闭摄像装置。整个录制过程耗时6 h。每个实验温度梯度均设置6个重复。
将人工洞穴行为定量装置置于玻璃水族箱中,控制水体到实验所需温度。将双齿围沙蚕置于装置中的人工洞穴内,待其稳定1 d后,开始录制。录制开始1.5 h后,关闭录像设备,放掉水族箱中的水,将装置放到恒温环境中,恒温环境的温度与实验所需温度一致。3 h后,再将装置缓慢放于玻璃缸中,录制1.5 h后关闭录像设备。每个实验温度梯度均设置6个重复。
本实验测定了双齿围沙蚕的径向起伏频率(F)、径向起伏泵水速率(R)、径向起伏泵水效率(P)、轴向爬行速度(V)和运动时间(T),其定义方法借鉴了房景辉等[12]的研究。除此之外,我们还测定了双齿围沙蚕头尾对调的次数(TF,头部与尾部互换位置的次数)和头尾对调一次的时间(TT,头部与尾部互换一次位置的耗时)。
2.3 实验装置
2.3.1 摄像装置
双齿围沙蚕在洞内的运动过程主要由400万像素的红外摄像头(杭州,海康威视 DS-2CD3345-I)记录,并且摄像头配有显示器和硬盘刻录机。调整摄像头位置(距离观测装置80 cm),以使双齿围沙蚕的运动过程可以被清楚地记录下来。
2.3.2 运动分析软件
将硬盘内双齿围沙蚕的运动视频导出,用ImageJ 2019软件(由National Institutes of Health开发)进行分析。ImageJ是科研中应用十分广泛的图像处理软件,可以手动追踪测量视频中物体的运动轨迹和距离。
2.3.3 观测装置
实验使用了底内动物行为学观测装置和人工洞穴行为学定量装置[12]对双齿围沙蚕在洞穴中的行为进行观察记录。
2.4 数据分析
所有的统计分析均由开源软件R3.6.3来执行。对每个数据集进行正态分布(Shapiro-Wilks)和方差齐性(Barlett)检验,对不符合条件的数据进行对数转换。采用双因素方差分析研究温度和时间段对沙蚕的径向起伏频率、轴向爬行速度、径向起伏泵水速率、径向起伏泵水效率、头尾对调次数和头尾对调时间的影响。使用LSD多重比较法进一步分析显著差异,并用Bonferroni法对结果进行校正。若p<0.05,则认为差异显著。
3 实验结果
3.1 径向起伏
温度和时间段对双齿围沙蚕的径向起伏均有显著影响(p<0.01),然而两者的交互作用无显著影响(p>0.05)(图1)。随着温度的升高,不同时间段内双齿围沙蚕的径向起伏频率均显著升高(p<0.05)。双齿围沙蚕涨潮30 min内的径向起伏频率比退潮前显著增加(p<0.05)。涨潮30 min内,双齿围沙蚕20℃时的径向起伏频率显著高于15℃时的(p<0.05),与25℃时的无显著差异(p>0.05)。20℃时,涨潮30 min内双齿围沙蚕的径向起伏频率相比退潮前增加的幅度最大。涨潮30 min内,双齿围沙蚕的径向起伏频率显著增加(p<0.05),但在涨潮30 min后,其径向起伏频率逐渐减小到退潮前的水平。
3.2 轴向爬行
温度、时间段以及两者的交互作用对双齿围沙蚕轴向爬行速度均有显著影响(p<0.05)。在退潮前、退潮后和涨潮30 min后3个时间段内,随着温度的升高,双齿围沙蚕的轴向爬行速度先升高后降低,并且均在20℃时达到最大值(图2)。退潮前,双齿围沙蚕在20℃和25℃的轴向爬行速度均显著高于15℃时(p<0.05)。涨潮30 min内,随着温度的升高,双齿围沙蚕的轴向爬行速度显著增大(p<0.05)。相同温度条件下,涨潮30 min内双齿围沙蚕的轴向爬行速度均达到最大值,并且在25℃时显著高于其他时间段(p<0.05),相较于退潮后增加的幅度最大。

图2 温度和时间段对双齿围沙蚕轴向爬行速度的影响Fig. 2 The influence of temperature and time periods on the axial crawling velocity of Perinereis aibuhitensisGrube
3.3 泵水速率
温度和时间段均对双齿围沙蚕的径向起伏泵水速率有显著影响(p<0.05),而两者的交互作用对其无显著影响(p>0.05)(图3)。随着温度的升高,双齿围沙蚕的径向起伏泵水速率先急剧增大再缓慢增大,20℃和25℃时的径向起伏泵水速率无显著差异(p>0.05)。同一温度条件下,涨潮后30 min内的双齿围沙蚕径向起伏泵水速率均高于其他时间段,而退潮前和涨潮30 min后的平均径向起伏泵水速率无显著差异(p>0.05)。

图3 温度和时间段对双齿围沙蚕径向起伏泵水速率的影响Fig. 3 The influence of temperature and time periods on the pumping rate of radial undulating motion of Perinereis aibuhitensisGrube
3.4 泵水效率
温度和时间段对双齿围沙蚕的泵水效率有显著影响(p<0.05),而两者的交互作用对泵水效率无显著影响(p>0.05)(表1)。随着温度的升高,双齿围沙蚕整体的径向起伏泵水效率在T3和T4时间段内先升高后降低,泵水效率在20℃时最大。当温度为15℃和20℃时,涨潮30 min内的泵水效率最大,并且涨潮30 min内的泵水效率与涨潮30 min后无显著差异(p>0.05)。

表1 温度和时间段对双齿围沙蚕径向起伏泵水效率(单位:mL/pulse)的影响Table 1 The influence of temperature and time periods on the pumping efficiency (unit: mL/pulse) of radial undulating motion of Perinereis aibuhitensisGrube
3.5 头尾对调
温度、时间段以及两者的交互作用对双齿围沙蚕头尾对调一次的时间均有显著影响(p<0.05)。随着温度的升高,退潮前、涨潮30 min内和涨潮30 min后双齿围沙蚕头尾对调一次的时间均显著降低(p<0.05)。退潮后,双齿围沙蚕20℃时的头尾对调一次的时间均高于15℃和25℃时的(图4)。当温度为15℃和20℃时,退潮后和涨潮30 min内双齿围沙蚕头尾对调一次的时间均低于退潮前,然而当温度为25℃时,双齿围沙蚕退潮后的头尾对调一次的时间显著高于其他时间段(p<0.05)。
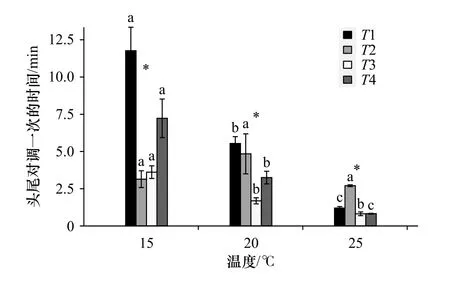
图4 温度和时间段对双齿围沙蚕头尾对调一次时间的影响Fig. 4 The influence of temperature and time periods on the time of head-tail exchange of Perinereis aibuhitensisGrube
温度、时间段和两者的交互作用对双齿围沙蚕的头尾对调次数均有显著影响(p<0.05)(表2)。随着温度的升高,双齿围沙蚕的头尾对调次数总体呈增加的趋势,并且退潮前25℃时的头尾对调次数显著高于15℃时(p<0.05)。当温度为15℃和20℃时,涨潮30 min内双齿围沙蚕的头尾对调次数均高于其他时间段,然而当温度为25℃时,双齿围沙蚕退潮前的头尾对调次数显著高于退潮后(p<0.05),与涨潮30 min内和涨潮30 min后无显著差异(p>0.05)。
3.6 径向起伏与轴向爬行时间
温度对双齿围沙蚕的径向起伏时间影响显著(p<0.05),而时间段和两者的交互作用对其无显著影响(p>0.05)(表3)。同一时间段内,随着温度升高,双齿围沙蚕径向起伏时间表现出先增加后减少的趋势。在退潮前和涨潮30 min内两个时间段内,双齿围沙蚕20℃时的径向起伏时间显著高于15℃时的(p<0.05)。涨潮后,随着时间的推移,双齿围沙蚕径向起伏的时间有减少的趋势。

表2 温度和时间段对双齿围沙蚕头尾对调次数的影响Table 2 The influence of temperature and time periods on the frequency of head-tail exchange of Perinereis aibuhitensisGrube

表3 温度和时间段对双齿围沙蚕运动时间(单位:min)的影响Table 3 The influence of temperature and time periods on motion time (unit: min) of Perinereis aibuhitensisGrube
温度和时间段对双齿围沙蚕的轴向爬行时间影响显著(p<0.05),而两者的交互作用对其无显著影响(p>0.05)(表3)。同一时间段内,随着温度的升高,轴向爬行的时间呈现先减少后增加的趋势。同一温度下,双齿围沙蚕涨潮30 min内的轴向爬行时间均多于其他时间段。
4 讨论
4.1 温度对双齿围沙蚕运动行为的影响
温度作为重要的环境因子之一,与生物的生长、繁殖、环境耐受能力和运动强度密切相关[1,12-14]。当环境温度发生变化时,生物会做出行为上的响应。王玲等[15]研究发现当温度为10~27℃时,温度升高,双齿围沙蚕的耗氧率也随之升高。研究显示,刺参的运动节律并未随着温度的升高而呈现一个连续性的变化,其在20℃时的平均运动速度和路程长度都高于其他实验温度,并且刺参活动的高峰存在温度的限制[16]。除了温度之外,其他环境因子也会影响生物的生命活动过程。陈松波[17]研究发现,在相同温度下,幼年鲤鱼的摄食率高于成年鲤鱼,并且在34℃时,不同规格的鲤鱼摄食率虽然均急剧降低,但是幼鱼摄食率仍然有9.45%。另外,同一温度下,团头鲂在水中的游泳速度将会随流速的增大而加快[18]。房景辉等[12]研究发现,大规格双齿围沙蚕在相同温度下的总泵水量更大。在不同的水体交换条件下,双齿围沙蚕幼体的生长状况在12倍/d流水下最佳,模拟潮汐次之[19]。对于海洋生物而言,每日的潮汐变化对其生活有重要的影响。潮汐变化过程中,生物体表会从充满水的状态向干露状态相互转变,而且双齿围沙蚕在低温保湿干露的条件下,大多能存活10 d[20]。因此,温度和潮汐对生物的共同作用可能会有别于温度的单一影响。
本研究发现,在同一潮汐阶段内,随着温度的升高,双齿围沙蚕径向起伏和头尾对调运动总体均有增强的趋势。但是,涨潮30 min内,双齿围沙蚕径向起伏频率在20℃的增长幅度(相较于退潮前)比15℃和25℃更大;在不同的潮汐阶段内,20℃时双齿围沙蚕的泵水效率均有高于15℃和25℃时的趋势,其径向起伏运动的时间均高于15℃和25℃时的,其轴向爬行运动的时间均有低于15℃和25℃时的趋势。除此之外,在退潮前、退潮后和涨潮30 min后3个潮汐阶段内,双齿围沙蚕的轴向爬行速度均在20℃时最大,但是在涨潮30 min内,双齿围沙蚕轴向爬行的速度在25℃时显著高于15℃和20℃时(p<0.05)。双齿围沙蚕行为随温度的变化表明,其运动强度随温度的升高而增强,但是涨潮后,面临水环境的剧烈变化,双齿围沙蚕加快径向起伏运动改善栖息环境的能力可能有一个极限状态,并在20℃左右能表现出来。双齿围沙蚕能通过轴向爬行来进行觅食和改善栖息水环境条件[12]。在高温条件下,涨潮后的双齿围沙蚕会因为对高温的不适应而加快轴向爬行的速度,希望通过轴向爬行来消除这种不适,这一结果与文献[21]结果一致。从双齿围沙蚕的运动时间可以看出,双齿围沙蚕在沉积物中主要以径向起伏运动为主,并且双齿围沙蚕的径向起伏和轴向爬行运动可能会同时发生。除此之外,王玲等[15]研究发现,温度从20℃升至29℃的过程中,双齿围沙蚕的耗氧率和排氨率随温度的升高而急剧升高。以上结果表明,20℃可能是双齿围沙蚕运动的一个临界温度,温度低于20℃时,双齿围沙蚕的运动强度较低;温度高于20℃时,双齿围沙蚕会表现出对高温的不适。双齿围沙蚕在20℃时的运动状态最佳,但是其最佳运动状态可能处于一个温度范围内,但具体范围还需要进一步研究。
4.2 潮汐更替对双齿围沙蚕行为的影响
海洋潮汐变化,包括潮汐更替、大小潮交替等过程,极大地影响了底栖动物栖息环境的水交换条件。在潮汐更替过程中,双齿围沙蚕的机体在上覆水浸泡与干露两种状态之间相互切换。本研究观察到,在这两种状态相互切换时,由于受到外部的刺激,双齿围沙蚕的行为也会随之发生变化。本研究发现,退潮后,双齿围沙蚕的轴向爬行速度降低;头尾对调一次的时间随温度的升高先增加后降低,25℃时头尾对调一次平均耗时最长;头尾对调次数减少;此阶段的轴向爬行时间较少。结果表明,退潮后,双齿围沙蚕的运动强度减弱。若退潮时环境温度较高,双齿围沙蚕的运动将更为迟缓。杂色刺沙蚕在退潮暴露后的2 h内,会在洞内外不断进行摄食活动,但是在洞外停留的时间不会超过50 s[9]。因此,双齿围沙蚕在洞内进行摄食活动时,为了尽可能获得更多的食物,会更加仔细地对沉积物进行搜寻,这种行为导致了其轴向爬行速度和头尾对调次数的降低,符合动物的最佳觅食理论[22]。双齿围沙蚕对干露有一定的耐受能力,这种能力在低温下表现得更加突出,其在低温保湿干露的情况下能生存10 d[20]。在干露过程中,水生动物体内会逐渐积累代谢废物并且高温环境会增加代谢废物的积累[14]。因此,双齿围沙蚕在低温环境时,退潮对其头尾对调的能力无明显负面影响,反而能促进其进行头尾对调。然而在高温环境中,退潮干露时,由于对高温的不耐受,双齿围沙蚕会通过降低机体的运动强度和运动时间,以此减少代谢废物对自身的负面影响。
双齿围沙蚕在水中溶氧低于某一数值时,会主动进行泵水运动,即存在泵水启动溶解氧阈值[12]。双齿围沙蚕在长久的干露状态下,其泵水启动的溶解氧阈值早已达到。除此之外,随着退潮时间的延长,双齿围沙蚕洞穴内部的有害物质会因沙蚕本身代谢和沉积物释放而逐渐累积,而涨潮能改善这种不利的条件。因此涨潮时双齿围沙蚕必然会更加迅速地启动泵水运动,这同时也是外部刺激导致动物释放行为的表现[22]。在本研究中,涨潮时伴随着大量新鲜海水的涌入,双齿围沙蚕的一系列运动都剧烈加快,表现在径向起伏频率和轴向爬行速度增加、径向起伏泵水速率和效率加快,头尾对调一次的时间更短,头尾对调次数显著增多(p<0.05)。结果表明,涨潮时,除了径向起伏和轴向爬行运动,双齿围沙蚕在洞穴中的头尾对调运动对水环境的改善也有重要的促进作用,并且温度较高时,这种运动能进一步被加强。多毛类的洞穴内部大多分布有黏液[23],黏液由多毛类自主分泌形成,有减少多毛类机体与洞穴壁之间的摩擦、保护体表皮肤和促进多毛类适应水体的作用。涨潮后,双齿围沙蚕的头尾对调运动可能对洞穴壁黏液的更新有着重要的作用。
动物在外部刺激的重复作用下会出现反应疲劳,称之为特定反应疲劳。如当一只枭出现在一只苍头燕雀面前时,苍头燕雀会不停地鸣叫作为对敌害刺激的应对,但是30 min后其鸣叫反应通常逐渐停止,重新恢复到之前的正常状态[22]。本研究发现,涨潮30 min后,双齿围沙蚕径向起伏的频率、泵水速率、泵水效率、轴向爬行的速度、头尾对调一次的时间和运动的时间均与退潮前无显著差异(p>0.05)。涨潮后,双齿围沙蚕在水体的刺激下,其运动会变得剧烈起来,但是随着时间的推移,其运动会逐渐恢复到退潮前的状态,并且所需的时间大约为30 min。除此之外,当温度为25℃时,涨潮30 min后,双齿围沙蚕的径向起伏频率、径向起伏泵水速率和轴向爬行速度均高于退潮前,并且头尾对调的次数仍低于退潮前。因此,在高温环境下,双齿围沙蚕恢复到退潮前的正常状态需要更长的时间(不止30 min),其适应高温环境的能力较差。
猜你喜欢
水道港口(2023年3期)2023-09-06 09:22:44
散文诗世界(2023年3期)2023-04-15 17:12:36
农民致富之友(2020年16期)2020-06-19 08:32:48
环球时报(2019-11-14)2019-11-14 05:14:40
精密制造与自动化(2018年1期)2018-04-12 07:42:47
作文与考试·初中版(2018年6期)2018-03-03 00:20:28
海洋与渔业(2017年7期)2017-12-08 07:13:17
儿童故事画报·自然探秘(2016年6期)2016-09-14 02:08:56
数学小灵通·3-4年级(2015年6期)2015-05-30 10:48:04
制造技术与机床(2015年10期)2015-04-09 07:06:10
