
玉树州境内草原毛虫种群分布及其对生境草场植被的影响
2022-09-22 04:05:20王海贞
环境昆虫学报 2022年4期
王海贞,刘 昕
(1.吕梁学院生命科学系,山西吕梁 033000;2.中山大学生命科学学院,广州 510275)
草原毛虫Gynaephoraqinghaiensis别名红头黑毛虫,隶属鳞翅目Lepidoptera毒蛾科Lymantriidae草原毛虫属Gynaephora,全世界共有15个种,其中,亚洲13个种,欧洲3个种,北美1个种,北极2个种,主要分布在北半球的高山以及北极的冻土地带,尤以高原高海拔(3 000 m以上)地区居多(Levinetal.,2003;张棋麟和袁明龙,2013;Yuanetal.,2016;Zhangetal.,2017)。在我国分布的草原毛虫共有8个种,全部为青藏高原特有种。
草原毛虫属完全变态发育类昆虫,一个完整的世代由卵、幼虫、蛹和成虫4个阶段组成,且一年发生一代(张棋麟和袁明龙,2013)。虫龄划分为雄虫6龄,雌虫7龄。1~2龄幼虫越冬后,翌年4月下旬至5月上旬开始活动。5月下旬至6月上旬为3龄幼虫盛期。幼虫第2个龄期长达6~7个月,其余龄期一般为15~20 d。7月上旬雄性幼虫开始结茧化蛹,7月下旬雌性开始结茧化蛹,一直持续到10月初才结束。8月上、中旬为化蛹盛期,8月初成虫开始羽化、交配和产卵。9月初,卵开始孵化,9月底至10月中旬为孵化盛期(王兰英,2012)。
草原毛虫是一种草地害虫,具有分布密度高、发生面积大、危害程度严重的特点(Wangetal.,2019;Wangetal.,2020;Wangetal.,2021)。据报道,西藏那曲地区聂荣县连续5年(1998-2002年)草原毛虫的成灾面积都在2.0×105hm2以上,其中2001年达到了5.9×105hm2,平均虫口密度200~500 头/m2,一些地区的虫口密度甚至达到1 000 头/m2以上(杨爱莲,2002;范小建,2011;顾慧洁等,2021);来自青海省的草原毛虫调查资料显示,2003年,青海省草原毛虫的发生面积达1.0×106hm2(何孝德和王薇娟,2003;张棋麟,2014);2009年,青海省海北州草原毛虫危害面积共计1.0×105hm2,严重危害面积6.7×104hm2(史国菊和吉汉忠,2010);2016年青海省黄南州草原毛虫发生面积约1.4×105hm2,危害面积约1.0×105hm2,虫口密度约128.5 头/m2(马青山,2018;王海春,2020)。草原毛虫不仅破坏草地植被,而且能引发牲畜口膜炎和口腔溃疡疾病,影响牲畜健康和草地畜牧业发展。2008年6月,青海省西宁市海晏县甘子河乡地区部分冬春草场爆发草原毛虫灾害,由于没有及时防治,从而导致该地区在2009年3-4月40多户牧民饲养的马、牦牛、藏羊等牲畜发生不同类型的口膜炎,发病率达到100%(尼玛等,2011;拉毛央尕,2020)。
玉树州位于青海省西南青藏高原腹地的三江源头,素有“中华水塔”的美誉,占据着我国重要的生态地位。然而,这里由于海拔高、气候寒冷等恶劣环境条件,导致生态环境非常脆弱,极易遭到破坏,且破坏后又极难修复(陈和庆等,2014)。同时,草场生态环境不仅是玉树州畜牧业的基础,也是三江源地区经济社会发展的重要组成部分,保护好草场生态资源,对玉树州乃至整个青海省的环境和经济社会可持续发展都具有非常重要的意义。近年来,随着全球气候变暖,雪线逐年上升,冰川逐渐消失,草地不断退化,虫害频繁爆发。有资料表明,玉树州植被的退化面积在逐年增多。全州范围内植被退化面积约560 hm2,占总植被面积的50%。其中,草原毛虫虫害就会导致玉树州高寒牧区草场50%~80%的牧草被蚕食殆尽(求松保和赵强国,2020),尤其在草原毛虫灾害爆发的严重地区,经常是牧草刚返青就被啃食一光,危害期长达半年之久,草场植被被严重破坏,加剧了草场退化及草地沙化的进程,曾经水草丰美的辽阔草场逐渐变成黄沙肆虐的“无人区”。有研究者对玉树州草原毛虫的种类进行了调查,发现主要为青海草原毛虫Gynaephoraqinghaiensis,集中分布在玉树、治多等地区(周尧和印象初,1979),但对于玉树州草原毛虫种群分布以及对植被危害的研究,近年来鲜有报道。本研究对玉树州草原毛虫的分布以及对生境草场植被的影响进行了调查分析,为玉树州草原毛虫的防控提供参考依据,对保护青藏高寒牧区草场生态环境,促进高寒牧区农牧业健康有序发展具有重要意义。
1 材料与方法
1.1 调查区域概况
调查区域涵盖青海省玉树州高寒牧区境内的嘉塘草原、巴塘草原、治多草原、曲麻莱草原和隆宝草原。研究区域中心位置地理坐标为34°17′53.052″N,94°25′38.964″E。东西跨度约736 km,南北跨度约499 km(图1)。

图1 草原毛虫及生境植被调查区域Fig.1 Survey area of Gynaephora qinghaiensis and its habitat vegetation注:图中红色方框内为本次调查区域;该图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2019)3333号的标准地图制作,底图无修改。Note:The red box in the figure was the survey area;This map was made based on the standard map No.GS(2019)3333 downloaded from the standard map service website of the National Administration of Surveying,Mapping and Geographic Information,without modification of the base map.
1.2 调查样地的布设
在玉树州高寒牧区(治多县、杂多县、称多县、曲麻莱县、玉树市)进行了为期5年(2015-2019年)的野外调查,并以国道G214和省道308为主线,在海拔4 188~4 580 m之间设置了10个调查样地(1#~10#),每个调查样地之间的距离均在20 km以上。10个调查样地的地理信息见表1。

表1 玉树州高寒牧区10个调查样地地理信息Table 1 Geographic information of ten sample plots in the alpine pastoral area of Yushu Prefecture
1.3 野外调查时间与方法
1.3.1调查时间
草原毛虫种群密度调查时间为2015-2019年每年6月下旬,草场植被调查时间为2016年6月下旬,且与2016年的草原毛虫种群密度调查同步进行。
1.3.2调查方法
(1)草原毛虫种群密度调查方法
草原毛虫种群密度调查以4~5龄幼虫为研究对象,采用随机抽样的调查方法,在每个调查样地内随机抽取5个样方(每个样方的规格为1 m×1 m),记录每个样方内草原毛虫幼虫的数量,取5个样方内草原毛虫幼虫数量的平均值作为该样地草原毛虫的种群密度。
(2)草甸植被调查方法
①植被抽样调查与物种鉴定
在每个调查样地内随机抽取5个样方,每个样方用1 m×1 m铁丝框(每个铁丝框用细绳分隔成25个小方格)取样(胡志坚,2010;李少松,2016),同时记录每个样方内的植物的种类、数量、盖度和总盖度,并计算物种丰富度指数(Species richness indexes,S)、物种多样性指数(Plant diversity index,D)和均匀度指数(Plant evenness index,E)(郭涛等,2007)。对于野外无法识别的植物物种,先做标记,并采集该植物标本,装在标本夹内带回实验室,经广东省热带亚热带植物资源重点实验室进行物种分类与鉴定。植物物种的分类与鉴定参考《中国植物志》全文电子版网站(http://frps.iplant.cn/)和中国自然标本馆(Chinese Field Herbarium,CFH)自然图库(http://www.cfh.ac.cn/Album/Albums.aspx)。
②植被盖度的测定
植被盖度是指植物地上部分覆盖地面的程度,分为总盖度、层盖度和分盖度。可借助1 m2的正方形采样框(框上用线绳分隔成25个小方格)调查植物盖度,每种植物想象地集中在一些网格内,并根据覆盖面积估算出总盖度和分盖度(李少松,2016)。
③物种多样性计算方法
A.物种多样性指数
物种多样性指数是指群落内种类多样性的程度,用来衡量群落或生态系统的稳定性。物种多样性指数是均匀度和丰富度相结合的函数,根据两个变量赋予的不同权重,物种多样性指数有多种计算方法。本研究选择Simpson指数(D)和Shannon-Wiener(H)指数来表示植被的物种多样性,其表达公式分别为:
D=1-∑Pi2
公式(1)
H=-∑PilnPi
公式(2)
B.均匀度指数(E)
公式(3)
C.物种丰富度指数(S)
S=样方内出现的物种数
公式(4)
以上各式中,Pi是第i个种的个数Ni占总个体数N的比例,各样地的多样性指数、均匀度指数和丰富度指数以每个样地内5个样方统计量的平均值表示。
1.1.4数据处理
利用SPSS 22.0软件分别对2015-2019年不同调查样地的草原毛虫种群密度及其2016年不同调查样地的植被指数等统计量进行单因素方差分析(One-way ANOVA);依据草原毛虫种群密度,对各个调查样地进行聚类分析(K-Mean聚类法);海拔高度对草原毛虫种群密度的影响采用单因素方差分析(One-way ANOVA);各统计量之间的相关关系采用皮尔森(Pearson)相关分析(Sig.2-tailed)。
2 结果与分析
2.1 草原毛虫种群分布调查
2.1.1草原毛虫种群密调查结果与显著性检验
2015-2019年的调查结果显示(表2),调查区域内草原毛虫种群密度在1.0~200.6 头/m2之间。同一个调查样地不同调查年份草原毛虫种群密度方差分析结果显示,2015-2019年,9#样地草原毛虫种群密度差异极显著(P<0.01),其它9个调查样地草原毛虫种群密度差异均不显著(P>0.05),表明在一定时期内草原毛虫种群总体上波动较小;同一调查年份不同调查样地草原毛虫种群密度方差分析结果显示,10个调查样地草原毛虫种群密度差异均极显著(P<0.01),表明草原毛虫在玉树州境内的分布情况并不相同,且具有聚集性分布的特点,集中分布在治多草原、嘉塘草原和隆宝草原。

表2 2015-2019年玉树州高寒牧区10个调查样地的草原毛虫种群密度调查结果Table 2 Survey result of population density of Gynaephora qinghaiensis from ten sample plots in the alpine pastoral area of Yushu Prefecture from 2015 to 2019
2.1.2调查样地海拔高度和坡向对草原毛虫种群密度的影响分析
(1)调查样地海拔高度对草原毛虫种群密度的影响分析
方差分析结果显示,海拔高度对草原毛虫种群密度的影响极显著(P<0.01);草原毛虫种群密度对应海拔高度的变化趋势如图2,海拔在4 188~4 580 m之间,随着海拔高度的增加,草原毛虫的种群密度无明显的变化规律,但种群密度较高的调查样地海拔大部分在4 200 m以上。
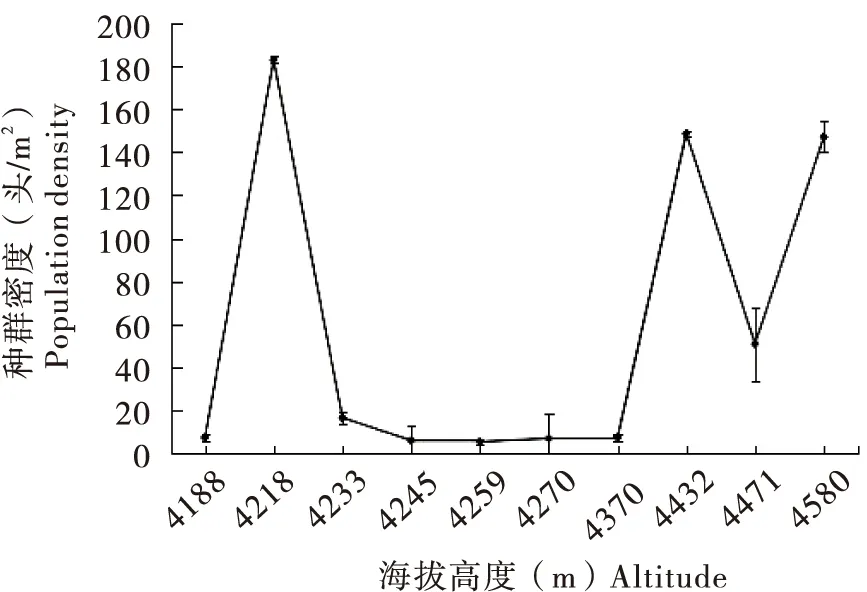
图2 草原毛虫种群密度对应海拔高度的变化趋势Fig.2 Change trend of population density of Gynaephora qinghaiensis to altitude
(2)调查样地坡向对草原毛虫种群密度的影响分析
依据草原毛虫的种群密度,对玉树州高寒牧区10个调查样地进行聚类分析,共获得3个类别,即高密度、中密度和低密度类别(表3)。其中,高密度类别调查样地包括6#、9#和10#样地,坡向分别为阳坡或无坡向;中密度类别调查样地包括4#样地,坡向为阳坡;低密度类别调查样地包括1#、2#、3#、5#、7#和8#样地,除2#样地无坡向外,其它样地均为阴坡。在10个调查样地中,高密度和中密度调查样地的坡向多为阳坡或无坡向,低密度调查样地的坡向大部分为阴坡,说明草原毛虫主要分布在阳坡或无坡向的高寒草甸,这与草原毛虫幼虫喜好阳光的生活习性有关。

表3 草原毛虫调查样地聚类及坡向信息Table 3 Clustering and slope direction of sample plots of Gynaephora qinghaiensis
2.1.3草原毛虫危害程度等级划分
根据于健龙和石红霄(2010)对高寒草甸草原毛虫危害等级划分标准(表4),在10个调查样地中,重度、极重度危害级调查样地共3个(6#、9#、10#),占调查样地总数量的30%;轻度危害级调查样地1个(4#),占调查样地总数量的10%;生态平衡级调查样地6个(1#、2#、3#、5#、7#、8#),占调查样地总数量的60%(表5)。

表4 高寒草甸草原毛虫危害等级划分标准Table 4 Standard for Hazards of Gynaephora qinghaiensis in the alpine meadow

表5 草原毛虫调查样地危害等级统计Table 5 Statistics on hazard levels of sample plots of Gynaephora qinghaiensis
2.1.4草原毛虫种群增长趋势分析
2015-2019年玉树州高寒牧区10个调查样地的草原毛虫种群密增长趋势图显示(图3),除 6#、7#和9#样地外,其它各个样地的草原毛虫种群密度总体呈逐年波动趋势,表明草原毛虫种群增长具有大小年变化规律。

图3 2015-2019年玉树州高寒牧区10个样地的草原毛虫种群增长趋势Fig.3 Increased trend of population density of Gynaephora qinghaiensis from ten sample plots in the alpine pastoral area of Yushu Prefecture from 2015 to 2019
2.2 草甸植被调查与分析
2.2.1调查样地植被组成分析
10个样地共有植被19科、38属、67种,其中,共有种为小嵩草Kobresiaparva,其次分布样地最多的种为珠芽蓼Polygonumviviparum。8个样地有特有种,其中,1#样地特有种为尖苞风毛菊Saussureasubulisquama、细叶西伯利亚蓼Polygonumsibiricum;2#样地特有种为卵叶风毛菊Saussureaovatifolia、甘青铁线莲Clematistangutica;3#样地特有种为细裂亚菊Ajaniaprzewalskii、鸦跖花Oxygraphisglacialis、高山米口袋Gueldenstaedtiaverna、美丽马先蒿Pedicularisbella、双花堇菜Vidabiflora;4#样地特有种为珠峰火绒草Leontopodiumhimalayanum、大黄Rheumpalmatum、草甸马先蒿Pedicularisroylei;5#样地特有种为条裂委陵菜Potentillalancinata、银光委陵菜Potentillaargyrophylla、圆叶点地梅Androsacegraceae、鼠掌老鹳草Geraniumsibiricum、马先蒿Pedicularissp.;7#样地特有种为节节草Equisetumramosissimum;8#样地特有种为风毛菊Saussureajaponic、穆坪高山耳蕨Polystichummoupinense;9#样地特有种为发草Deschampsiacaespitosa、大萼蓝钟花Gyananthusmacrocalyx;6#和10#样地无特有种。
2.2.2草甸植被调查与分析
(1)植被指数调查结果与显著性检验
植被指数调查结果显示(表6),植被总盖度在58.2%~94.4%之间,Simpons多样性指数在0.54~1.19之间,Shannon-Wiener多样性指数在0.99~1.41之间,均匀度指数在0.51~0.70之间。方差分析结果显示(表8),不同调查样地的Shannon-Wiener多样性指数和均匀度指数差异性均不显著(P>0.05),但Simpons多样性指数和植被总盖度存在极显著差异(P<0.01)。

表6 植被指数调查结果Table 6 Survey results of vegetation indexes

表7 10个调查样地植被指数方差分析Table 7 Variance analysis of vegetation index from ten sample plots
(2)相关性分析
利用SPSS 22.0软件对2016年调查的10个样地草原毛虫种群密度、植被总盖度、均匀度指数、Simpons多样性指数、Shannon-Wiener多样性指数两两之间进行皮尔森相关分析,从分析结果得出(表8),草原毛虫种群密度与草甸植被多样性指数以及均匀度指数之间的相关性均不显著(P>0.05),表明草原毛虫种群密度对草甸植被多样性指数和均匀度指数的影响较小。草原毛虫种群密度与植被总盖度之间呈极显著的负相关关系(P<0.01),表明随着草原毛虫种群密度的增大,草甸植被总盖度总体呈逐渐减小的趋势(图4)。

表8 2016年草原毛虫种群密度和植被指数之间的皮尔森相关系数Table 8 Pearson’s correlation coefficient between population density of Gynaephora qinghaiensis and vegetation indexes in 2016

图4 2016年草原毛虫种群密度对植被总盖度的影响趋势Fig.4 Influence trend of population density of Gynaephora qinghaiensis on total vegetation coverage in 2016
3 结论与讨论
青藏高寒牧区居青藏高原腹地,是牦牛、藏羊的主要产地,也是最主要的牧业基地。这里气候凉爽,太阳辐射强,5-7月份降水丰富,为草原毛虫的分布、为害提供了有利的外界条件。仅玉树州境内的草原毛虫分布面积就高达5.4×105hm2,占青海省草原毛虫分布总面积的50.9%,其中,重度以上为害面积达3.2×105hm2,占全省为害总面积的45.7%(何孝德和王薇娟,2003)。根据何孝德和王薇娟(2003)对青海省草原毛虫分布区域的划分,玉树州被定位为江河源头草原毛虫重灾区Ⅰ类亚区。根据于健龙和石红霄(2010)对高寒草甸草原毛虫危害等级划分标准,在玉树州调查的10个样地中,有3个样地的草原毛虫已达重度、极重度危害等级(种群密度大于或等于130 头/m2),占调查样地总数的30%。说明玉树州境内的草原毛虫分布集中,部分地区危害严重,需及时采取防治措施。
草原毛虫雌成虫翅已退化,与雄虫交配后在原地结茧产卵,因此,草原毛虫只能依靠幼虫近距离迁移,这直接导致草原毛虫种群呈聚集分布型,并随着长期以来的生存特点逐渐形成点状与岛屿状分布(严林,2006)。据报道,青海草原毛虫分布中心点密度高达1 000 头/m2(杨爱莲,2002;严林,2006)。本研究通过野外调查发现,青藏高寒牧区草场的草原毛虫同样呈聚集型分布,集中分布在嘉塘草原、隆宝草原和治多草原,种群密度最大达200.6 头/m2,以此为核心的周边区域草原毛虫种群密度较小,有些地方甚至没有草原毛虫分布。草原毛虫聚集分布习性以及较强的子代繁殖能力可能是玉树州部分地区草原毛虫成灾的主要原因。草原毛虫喜食植物种类较少,尤其喜食嵩草属植物,如小嵩草、矮嵩草Kobresiahumilis等,并以这些植物的茎尖、叶端和叶缘为主要取食部位(万秀莲和张卫国,2006)。因此,草原毛虫种群分布还受限制于喜食植物的空间格局。万秀莲和张卫国(2006)研究表明,草原毛虫总体空间格局上呈聚集型分布,但在小尺度下则呈均匀型分布,且虫口密度与喜食植物的丰富度和多样性具有紧密的关联。随着喜食植物丰富度和多样性的增加,草原毛虫种群密度显著上升,随着喜食植物丰富度和多样性的减少,草原毛虫种群密度显著下降。在野外调查中同样发现,嵩草属植物盖度较高的草甸,草原毛虫种群密度较大,嵩草属植物盖度较低,且其他种类植物较多的草甸,草原毛虫种群密度较小。此外,草原毛虫种群分布还受海拔高度和坡向的影响,草原毛虫种群密度受海拔高度的影响极显著,且高密度的调查样地海拔大部分在4 200 m以上。阚绪甜(2016)对青藏高原草原毛虫种群密度调查结果显示,草原毛虫主要分布在海拔4 074~4 450 m之间,低于4 000 m的海拔区域基本没有毛虫分布。青藏高原高海拔地区是一种寒冷且缺氧的极端生境,草原毛虫对这种极端生境表现出的适应性是由其自身的遗传机制决定的(张棋麟,2014;Yuanetal.,2015;杨兴卓等,2018)。研究表明,高海拔极端生境生活的昆虫能够调整自身基本代谢过程适应低氧压力(Zhaoetal.,2013)。在野外调查中还发现,草原毛虫幼虫主要分布在阳坡或无坡向的高寒草甸,这可能与草原毛虫幼虫喜好温暖、阳光充足的气候有关,尤其是进入快速生长期的幼虫(5~6龄),需在白天吸收阳光以满足自身生长发育的能量需求。
高寒草甸是青藏高原主要的天然草地类型(孙飞达等,2009),不仅是畜牧业发展的资源依托,而且也是许多适应极端生境物种分化变异的中心(江小雷等,2004;于健龙和石红霄,2010)。植物群落结构和土壤环境是高寒草甸生态系统最基本的参照特征(孙飞达等,2009),其稳定性是草甸生态系统存在的必要条件和功能表现。草原毛虫作为青藏高原高寒草甸的主要害虫之一,主要取食莎草科、禾本科、豆科等各类占植被群落优势地位的牧草,尤其对对嵩草属植物表现出强烈的偏好和倾向性(万秀莲和张卫国,2006)。在草原毛虫密集区,大量的嵩草属植物被草原毛虫蚕食殆尽,导致其在植物群落结构中的竞争力下降,其他植物物种的竞争力增强,随之,植物群落中物种生态位发生变化,进而影响到整个草甸生态系统的稳定性。草甸生态系统的失衡,又进一步加剧草甸退化和草甸生态系统恶化。马培杰等(2016)研究表明,草原毛虫对小嵩草草甸植被总生物量、盖度和丰度的影响不大,但能显著降低植被的Simpson指数、Shannon-Wiener指数和Pielou指数。于健龙和石红霄(2010)对高寒嵩草草甸植被调查结果显示,草原毛虫虫口密度与高寒嵩草草甸地上生物量具有显著的正相关关系,与草甸地下生物量、生草层厚度和牧草高度具有显著的负相关关系,与植被总盖度的相关性不显著,但当虫口密度达到重度、极重度危害等级时,植被总盖度显著降低。在本研究中,草原毛虫种群密度与高寒草甸植被多样性指数和均匀度指数之间的相关性均不显著(P>0.05),但与植被总盖度之间具有极显著的负相关关系(P<0.01),随着草原毛虫种群密度的增加,草甸植被总盖度总体呈逐渐减小的趋势。草原毛虫幼虫喜食小嵩草、矮嵩草和大花嵩草Carexnudicarpa等嵩草属的植物,喜食植物种类较少(万秀莲和张卫国,2006),因此不会影响草甸植被的多样性和均匀度,但嵩草属植物大多为草甸优势种,植被覆盖度较大,当草原毛虫种群密度过高时,这些优势植物就会被大面积破坏,从而使草甸植被总盖度明显减小。
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
作文小学高年级(2023年9期)2023-10-19 02:54:20
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
疯狂英语·新读写(2022年1期)2022-11-23 04:44:18
青海草业(2022年2期)2022-07-23 09:34:58
西藏艺术研究(2019年1期)2019-09-04 09:39:54
活力(2019年21期)2019-04-01 12:17:10
读者·校园版(2015年8期)2015-05-14 13:11:41
小雪花·小学生快乐作文(2009年5期)2009-05-22 02:18:34
