
越冬前北方不同类型强冬性冬油菜形态及生理响应研究
2022-09-21 06:16刘丽君姚彦林蒲媛媛王旺田李学才孙万仓武军艳
干旱地区农业研究 2022年5期
李 鹏,马 骊,徐 芳,刘丽君,姚彦林,蒲媛媛 王旺田,李学才,方 彦,孙万仓,武军艳
(1. 甘肃农业大学农学院,甘肃省油菜工程技术研究中心,甘肃 兰州 730070;2.省部共建干旱生境作物学国家重点实验室,甘肃 兰州730070;3.甘肃农业大学生命科学技术学院,甘肃 兰州 730070)
油菜作为我国四大油料作物之一,是继水稻、小麦、玉米、大豆之后我国的第五大农作物。最近几年,虽然我国油菜的播种面积和产量均排在世界前列,但是国产的食用油脂依然比较少,在国内消费市场仅占35%~40%,我国食用植物油的供给一直处于严重短缺的状态[1-3]。我国北方地区冬季寒冷春季干旱,冬季气温在0℃以下,甚至有时候极端低温[4],低温对油菜造成的伤害已经逐渐成为了农业生产中非常重要的问题,因此对品种的抗寒性要求也越来越高。在北方地区能够安全越冬的冬油菜类型主要是白菜型冬油菜,甘蓝型冬油菜因为其越冬率太低而无法大面积推广种植[5]。但甘蓝型油菜具有抗倒伏、抗旱、产量高等优势,最近几年,随着强冬性甘蓝型冬油菜品种的选育成功,其在北方冬油菜产区部分地区开始种植,但因为其抗寒性还是不如白菜型冬油菜[6],遭遇极端低温天气时无法保证正常越冬,产量不稳定,所以改良甘蓝型冬油菜品种的抗寒性成为北方冬油菜产区重要的育种目标之一[7]。
在外界环境变化时叶片是最先感知的器官,同时又作为光合作用的主要场所,它的形态变化是植物适应环境的基础[8]。在低温胁迫下,植物直立生长相对于匍匐生长更容易受到低温损伤,匍匐生长的油菜在生长过程中会随温度降低生长点逐渐下陷[9],同时植株叶片开始卷曲皱缩,叶面积也开始逐渐减小[10]。低温胁迫下,植物体内会发生一系列生理生化反应,致使各项生理指标发生变化,这些生理指标的变化在一定程度上可以反映植物抗寒性[11]。渗透调节物质的作用主要是参与合成细胞内的有机溶质,这些有机溶质中有许多有机小分子,包括脯氨酸(Pro)、甜菜碱、甘油等,还包括一些可溶性物质,比如可溶性糖(SS)、可溶性蛋白(SP)等,在植物受到低温的影响后,这些有机小分子能够通过增加细胞的渗透势来降低细胞的冰点,防止由于过度脱水而对细胞造成危害[12-17]。钙离子作为细胞内重要的第二信使,参与了植物对环境信号的应答反应。各种生物或非生物逆境包括病原菌侵染、干旱、盐害、冷害、热害等均能诱导植物细胞质Ca2+浓度提高[18]。Ca2+通过维持应激状态下的信号转导和细胞结构稳定性来减轻Na+的毒性作用[19]。K+在维持细胞质中正负离子平衡、激活关键的酶促反应、维持液泡的渗透压等方面发挥着重要作用[20]。前期研究结果表明,白菜型冬油菜的抗寒性强于甘蓝型冬油菜品种,蒲媛媛等[7]、马骊等[21]发现不同抗寒性白菜型冬油菜和甘蓝型冬油菜在遭受低温冷冻之后都会通过升高叶片中可溶性物质来保护叶片细胞的膜结构,但增加量的多少又反映了白菜型与甘蓝型冬油菜遭受冷害之后自身代谢的差异。
因此,本研究选择强抗寒性白菜型油菜品种陇油6号和甘蓝型油菜品种2019-QL-GAU-30,比较两个不同类型强抗寒性品种在越冬前形态变化、光合特性和渗透调节特性,探寻不同类型冬油菜抗寒性机理的差异,为选育强抗寒性甘蓝型冬油菜提供理论基础。
1 材料与方法
1.1 试验材料
本试验所用的甘蓝型冬油菜品种2019-QL-GAU-30(2017—2020年多点平均越冬率83%)和白菜型冬油菜品种陇油6号(2017—2020年多点平均越冬率98%)均由甘肃农业大学油菜课题组提供,试验地海拔2 150 m,103°40′ E, 36°03′ N,年平均气温6.5℃,平均降雨量175 mm。
1.2 试验设计
试验于2020年8—11月在甘肃省永登县上川镇甘肃省油菜工程技术研究中心进行。小区面积4 m2,3次重复,随机区组排列,试验材料播种方式为点播,行距20 cm,株距10 cm,于五叶期开始取样。具体取样日期及取样温度见表1。(由于需要根据温度的具体变化和油菜形态特征变化进行测定和取样,故部分指标测定时间不一致)。

表1 取样日期及对应气温表
1.3 试验方法
1.3.1 冬油菜干物质的测定 于越冬前不同温度时期取样,每次3个重复,测定干物质的积累量。在尽可能不损失根系的情况下将植株完整挖出,用清水将植株冲洗干净,从子叶节处切开分成地上部与地下部,用滤纸吸干表面水分,分别用烘箱进行烘干(105℃杀青40 min,85℃烘干至恒重)和电子天平称重(精确度0.001 g,上海越平科学仪器有限公司)。分别计算植株地上部、地下部含水量和根冠比。
1.3.2 叶面积的测定 于10月18日、11月15日、11月22日每一个小区选择有代表性的植株3株分别测定其长柄叶、短柄叶、无柄叶的最大宽和最大长并分别计算3种叶片的叶面积。叶面积(cm2)=长×宽×0.75[22]。
1.3.3 叶绿素SPAD值的测定 选取长势相同的6株植株用叶绿素仪分别测定植株心叶、心外1叶、心外2叶、心外3叶、心外4叶、心外5叶的叶绿素SPAD值。
1.3.4 光合指标Pn、Gs、Tr和Ci的测定 测定日10∶00—12∶00采用GFS-3000光合仪(德国WALZ),使用开放气路,分别测定已标记的6株植株第3片功能叶的光合指标。
1.3.5 渗透调节物质的测定 采集3株植株展开的第3片叶和地下部,用于生理指标的测定。用考马斯亮蓝法测定可溶性蛋白含量[23];用磺基水杨酸法测定脯氨酸含量[23];用蒽酮法测定可溶性糖含量[23];用硝酸消解、ICP-AES法测定钠钾钙离子含量[24],每个指标重复测3次。
1.4 数据处理
采用Origin 2021进行试验数据的处理和图表制作,采用SPSS 22.0软件进行数据的显著性分析。
2 结果与分析
2.1 越冬前冬油菜形态变化
2.1.1 越冬前冬油菜地上部形态变化 分别在10月18日、11月15日和11月29日记录了白菜型冬油菜陇油6号和甘蓝型冬油菜2019-QL-GAU-30的形态特征变化,见图1。10月18日两种冬油菜都是直立或者半直立生长,叶色为绿色(图1A和图1B)。11月15日两种冬油菜叶片匍匐在地面,叶片颜色变成深绿并带有紫色,老叶变黄(图1C和图1D)。11月29日时,白菜型冬油菜叶片萎蔫,并完全匍匐于地面,老叶变黄、干枯;甘蓝型冬油菜叶片萎蔫皱缩,匍匐于地面,叶色变深,老叶变黄干枯(图1E和图1F),但是白菜型冬油菜陇油6号的黄化萎蔫程度大于甘蓝型冬油菜2019-QL-GAU-30,这可能是白菜型冬油菜通过形态变化主动应对低温,而甘蓝型冬油菜是被动适应低温的一个过程。

注:A 、C和E分别为2019-QL-GAU-30在10月18日、11月15日和11月29日形态特征。B、D和F分别为陇油6号在10月18日、11月15日和11月29日形态特征。Note: A, C, and E are the morphological characteristics of 2019-QL-GAU-30 on October 18, November 15, and November 29, respectively. B, D, and F are the pattern characteristics of Longyou 6 on October 18, November 15, and November 29, respectively.
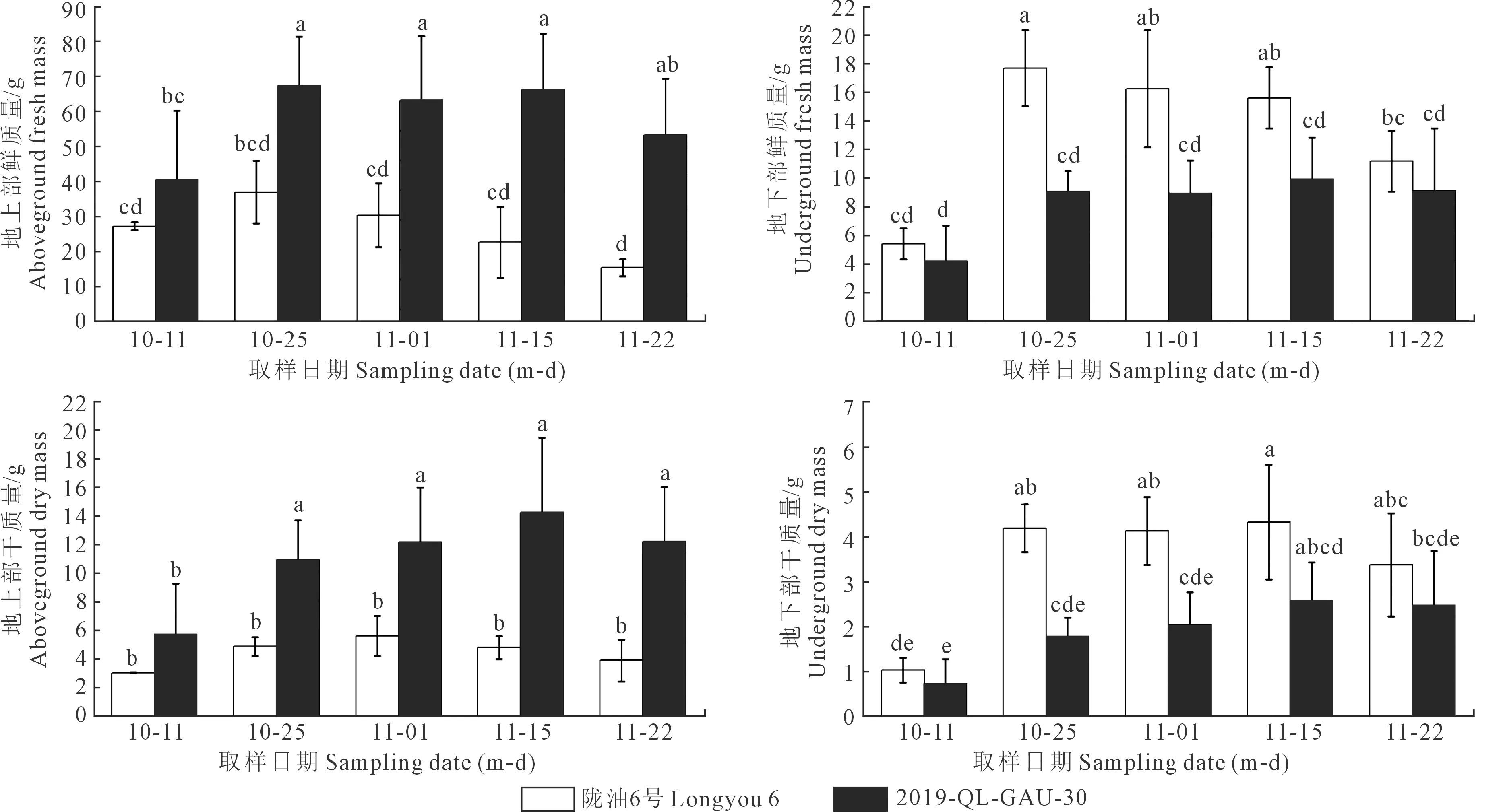
2.1.2 越冬前冬油菜植株形态及其干、鲜质量的变化 越冬前两种冬油菜植株形态如图2所示。陇油6号和2019-QL-GAU-30地下部分的鲜质量和干质量都随着温度变化逐渐增大,但陇油6号大于2019-QL-GAU-30;2019-QL-GAU-30的地上部鲜质量和干质量都大于陇油6号。这可能是由于越冬前白菜型冬油菜陇油6号主动应对低温已经开始向地下部储存营养物质来保证正常的越冬,而甘蓝型冬油菜2019-QL-GAU-30被动应对低温还是在进行正常的营养生长。

注:A、B分别为10月18日和11月1日陇油6号和2019-QL-GAU-30整株形态特征对比。Note: A and B are the comparison of the morphological characteristics of the whole plant of Longyou 6 and 2019-QL-GAU-30 on October 18 and November 1, respectively.
2.1.3 越冬前冬油菜根冠比和组织含水量的变化 由图4可见,陇油6号和2019-QL-GAU-30的根冠比都逐渐增大,干物质逐渐向地下部分积累,为越冬准备充分的营养物质;陇油6号根冠比的增加量显著高于2019-QL-GAU-30,这可能与二者抗寒方式的差异有关。叶片和根系的含水量都表现逐渐降低的趋势,但变化幅度较小,品种间差异不显著。
2.1.4 越冬前冬油菜叶面积的变化 由图5可见,陇油6号和2019-QL-GAU-30的长柄叶、短柄叶和无柄叶的叶面积均呈现出减小的趋势,但变化不显著。2019-QL-GAU-30的长柄叶叶面积下降幅度大于陇油6号,可能是因为温度过低冬油菜营养生长受阻,叶片卷曲皱缩,阻碍了甘蓝型冬油菜长柄叶叶面积的增加。
注:不同字母表示处理间差异显著(P<0.05),下同。Note: Different letters indicate significance differences between treatments (P<0.05), the same below.
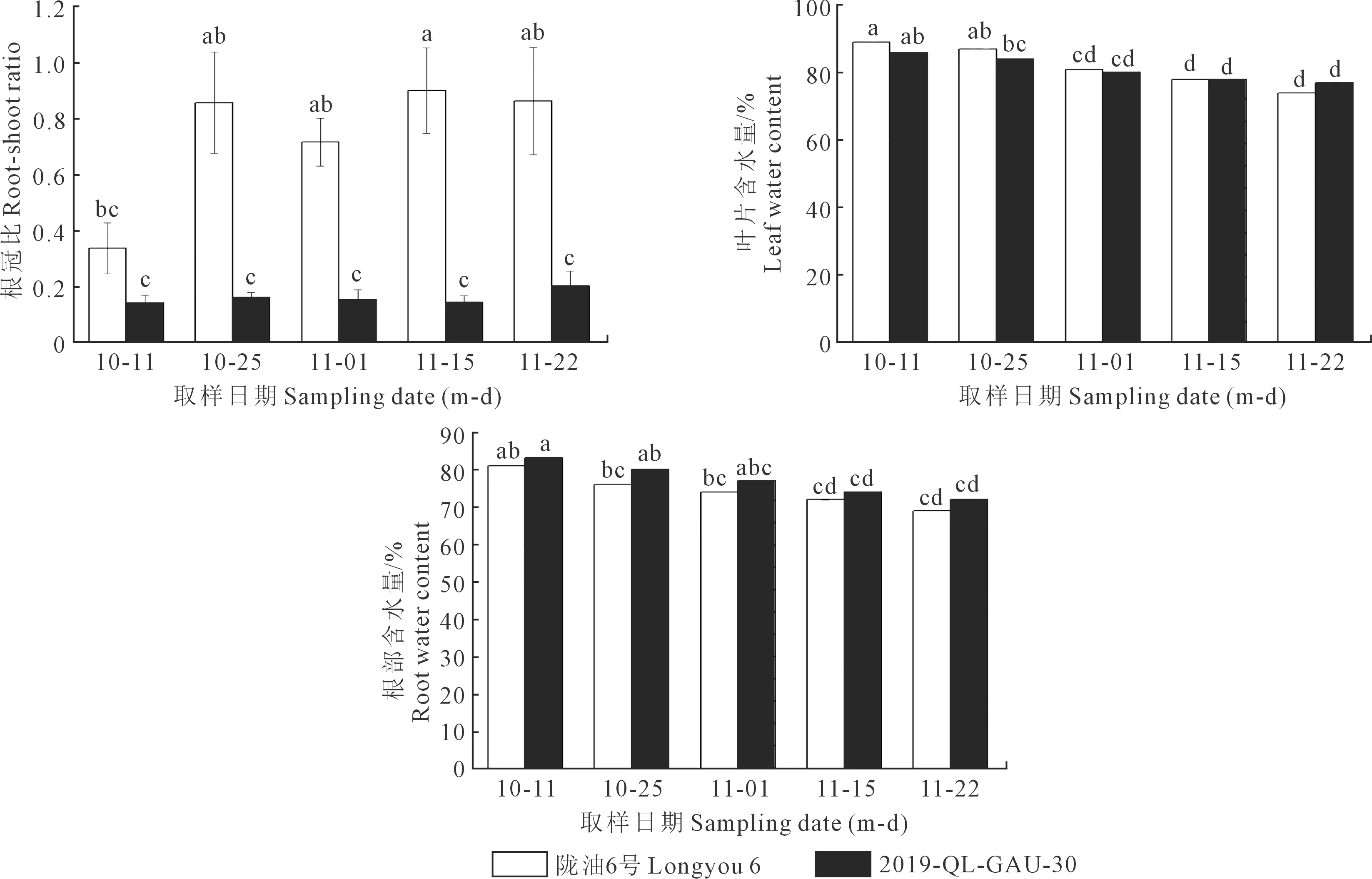
图4 冬油菜根冠比和组织含水量的变化Fig.4 Changes in root-shoot ratio and tissue water content of winter rapeseed
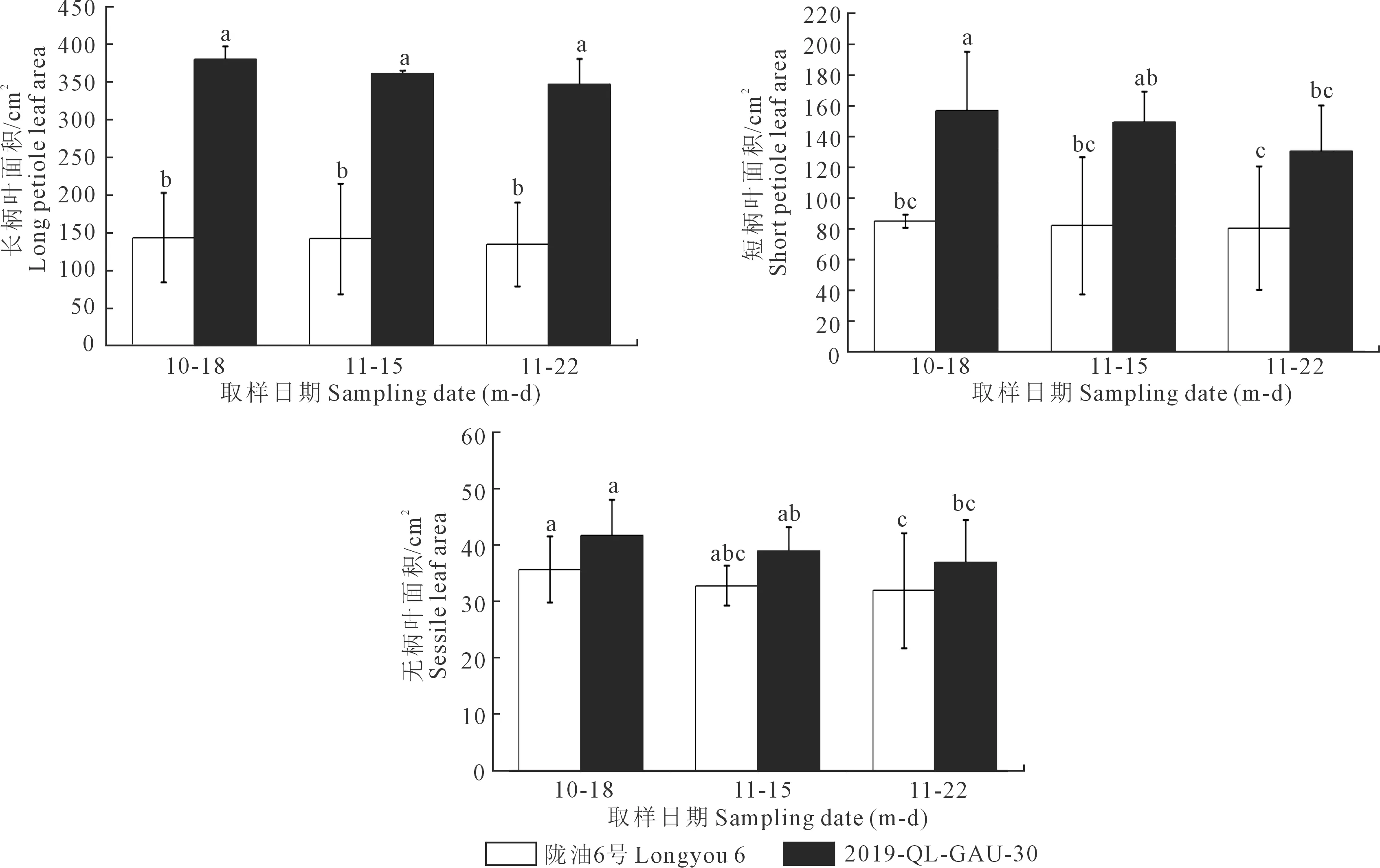
图5 冬油菜长柄叶、短柄叶和无柄叶叶面积的变化Fig.5 Changes in leaf area of long-stalked, short-stalked, and sessile leaves in winter rapeseed
2.2 越冬前降温过程中冬油菜光合指标的变化
2.2.1 叶绿素SPAD值的变化 陇油6号心外1叶、心外2叶SPAD值表现为先增大后减小,而心外3叶、心外4叶和心外5叶变化不明显。这可能因为温度降低后合成叶绿素的酶活性降低导致叶绿素合成受阻。而甘蓝型冬油菜所有叶片的SPAD值都逐渐增大,但差异不显著,这可能是由于细胞脱水导致,这一结果与其形态变化相一致。
2.2.2 光合参数的变化 如图6所示,陇油6号和2019-QL-GAU-30的净光合速率、蒸腾速率、气孔导度都随时间进程显著下降,而胞间CO2浓度显著上升,且陇油6号的气孔导度、净光合速率和蒸腾速率高于20119-QL-GAU-30,但差异不显著,可能是因为低温导致气孔关闭,但陇油6号抗寒性较强,故气孔开度大于2019-QL-GAU-30。
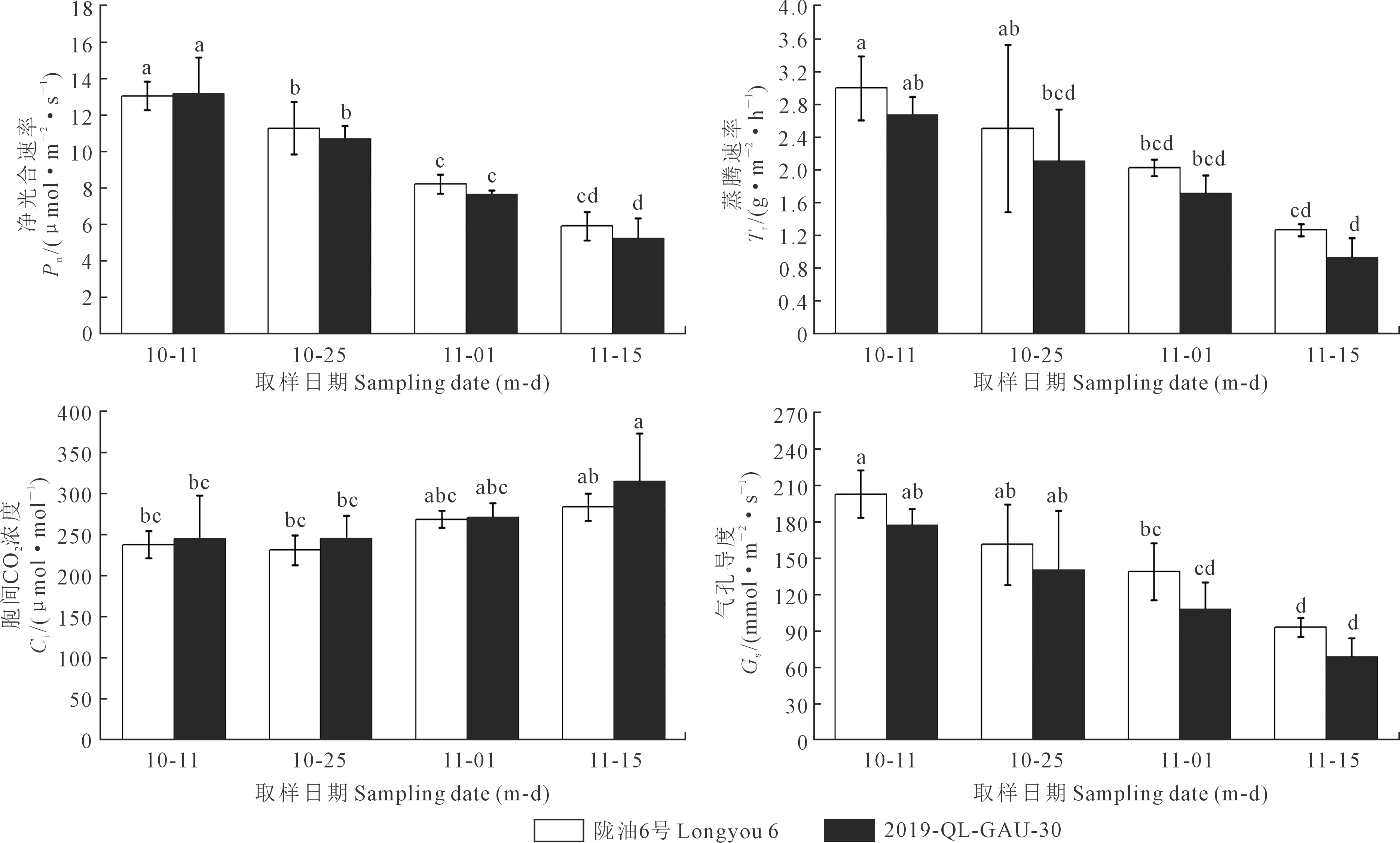
图6 净光合速率、蒸腾速率、胞间CO2浓度和气孔导度的变化Fig.6 Changes in Pn,Tr,Ci and Gs
2.3 越冬前冬油菜渗透调节物质的变化
2.3.1 叶片渗透调节物质的变化 陇油6号和2019-QL-GAU-30叶片的可溶性蛋白含量和脯氨酸含量均显著上升,但随着温度逐渐降低其积累速度变缓。可溶性糖含量先升高后降低,11月1日达到最大值。叶片钠离子含量的变化都呈现出逐渐降低的趋势;而钾离子和钙离子含量的变化均为先上升后下降,且在10月25日达到最大值。
2.3.2 根部渗透调节物质的变化 陇油6号和2019-QL-GAU-30根部的可溶性蛋白含量和脯氨酸含量都表现为平缓上升的趋势;而可溶性糖含量呈现出先下降然后上升的趋势,且上升幅度较大。比较叶片和根部的可溶性蛋白含量、脯氨酸含量和可溶性糖含量,发现这3种渗透调节物质在根部的含量均大于叶片,这可能因为冬油菜为了能够正常越冬根部积累大量的营养物质来保证越冬期间正常的营养供给。随着气温的降低,白菜型冬油菜根部钠离子含量逐渐升高,甘蓝型冬油菜根部钠离子含量先上升然后下降,在11月1日达到最大值。两种冬油菜钾离子和钙离子的含量都是先降低然后升高,都在11月1日有最小值。与叶片离子含量相比,陇油6号根部Na+含量逐渐增加,这可能是低温限制了气孔的开度,降低了叶片的蒸腾速率,阻碍了Na+由根部向叶片的运输,减小了Na+对叶片的毒害作用。而叶片Ca2+的含量远大于根部,这可能是由于较高的Ca2+含量可以保证冬油菜正常的光合作用和增加细胞渗透势,保证正常的生长发育。
2.4 越冬率与各指标间的相关性
相关性分析结果表明,越冬率与可溶性糖含量、叶鲜质量、叶干质量和Ca2+含量(根)呈极显著负相关(P<0.001);越冬率与根冠比和根干质量呈极显著正相关(P<0.001),与根鲜质量和Ca2+含量(叶)呈显著正相关(P<0.05)。表明叶片大量的营养物质向根部运输,根部通过积累营养物质和调节离子转运来增加冬油菜的抗寒性,以适应低温环境。

表2 叶片叶绿素SPAD值的变化

图7 冬油菜叶片渗透调节物质的变化Fig.7 Changes in leaf osmotic adjustment substances of winter rapeseed
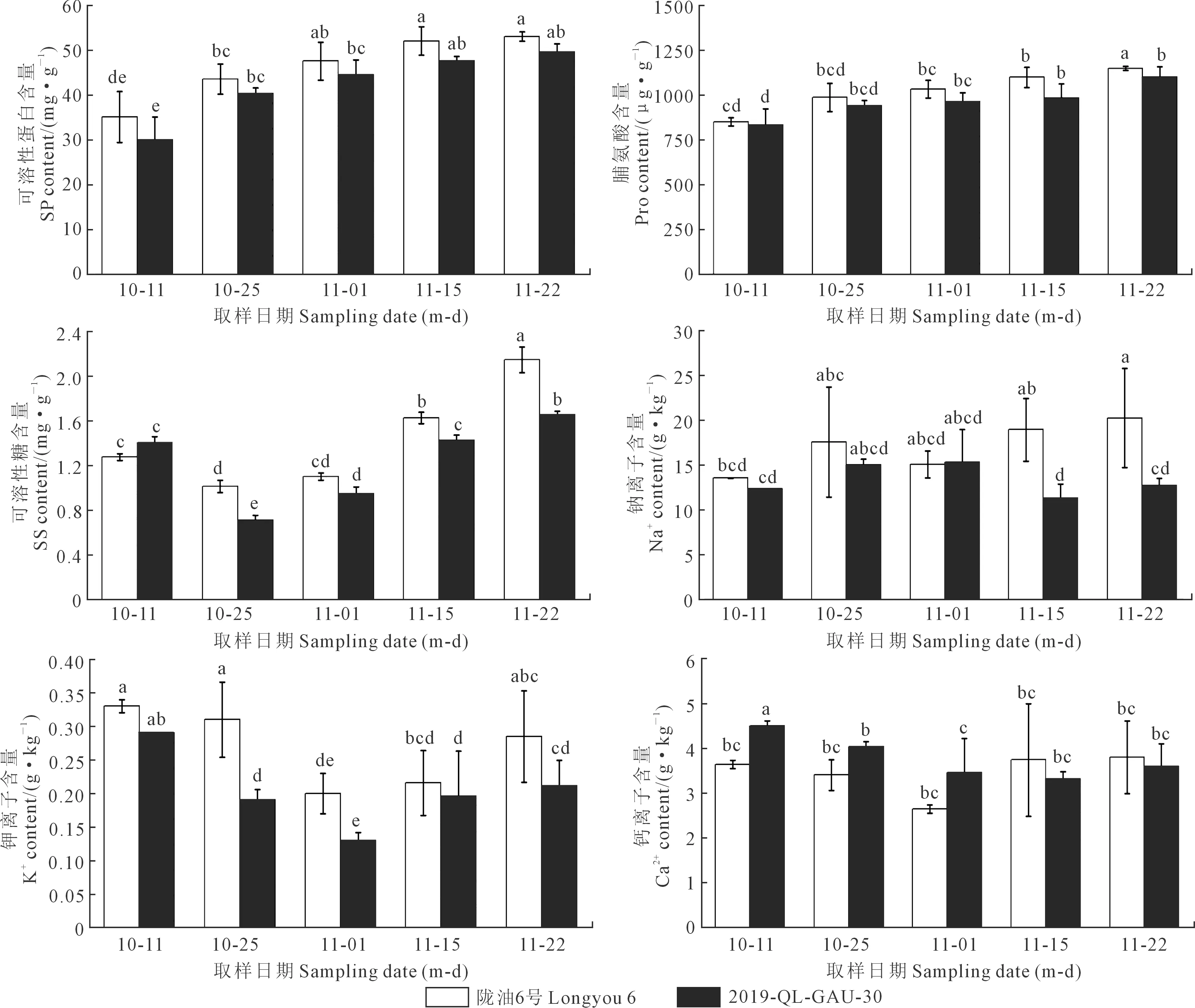
图8 冬油菜根部渗透调节物质的变化Fig.8 Changes in osmotic adjustment substances in the roots of winter rapeseed
3 讨 论
3.1 低温胁迫对冬油菜形态特征的影响
由于北方地区冬季寒冷干燥,冬油菜更容易受到冻害的影响,其中越冬前是油菜最易发生冻害的3个关键时期之一。油菜植株的叶片、根部也是容易遭受冻害的主要部位,当油菜植株对温度敏感的部位受到冻害时,会导致叶片干枯萎蔫、烂根、死苗等,严重影响其营养生长及生殖生长[25]。在越冬前为了适应低温,冬油菜叶片形态发生变化,植株呈半直立或者匍匐生长,叶色深绿,稍微带一点紫色或红色。本研究中,随着温度的逐渐降低,两种不同类型冬油菜都能通过增大根冠比、降低组织含水量、降低叶面积来保证正常的越冬,营养物质向根部运输,根部积累大量有机物,导致根冠比增加。白菜型冬油菜陇油6号的根系直径、根冠比均显著大于甘蓝型冬油菜2019-QL-GAU-30,含水量小于2019-QL-GAU-30,可见根系、根冠比是白菜型冬油菜越冬抗寒性强的重要形态特征。这与蒲媛媛等[7]、马骊等[21]、武军艳等[26]研究结果一致。因此,选育根系粗、根冠比大的材料,对提高其越冬抗寒性可能更有利 。
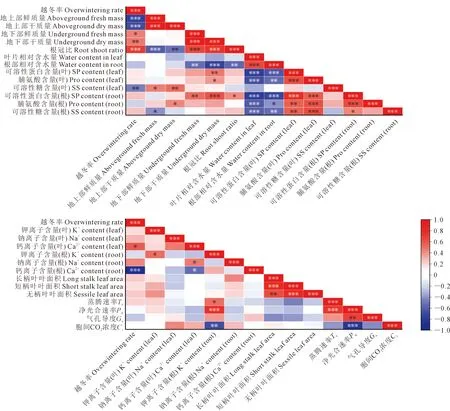
注:*,**,***分别表示在P<0.05,P<0.01,P<0.001水平差异显著。红色表示正相关性,蓝色表示负相关性;颜色深浅表示相关性系数值的大小,颜色越深,相关性越强。Note: *, ** and *** indicate significant differences at the level of P<0.05,P<0.01 and P<0.001, respectively. Red indicates positive correlation, blue indicates negative correlation; the depth of color indicates the magnitude of the correlation coefficient value, the darker the color, the stronger the correlation.
3.2 低温胁迫对冬油菜光合特性的影响
光合作用是多数植物受冷胁迫影响最明显的过程之一[27-28],气孔导度控制着叶片的蒸腾能力[29],气孔导度下降导致胞间CO2供应不足,因光合原料的缺乏,净光合速率的降低使光合产物的积累量减小[30]。本研究表明,随着温度的降低,两种不同类型冬油菜净光合速率、蒸腾速率和气孔导度都有不同程度的下降,而胞间CO2浓度增大,这可能是冬油菜通过调整气孔的开闭状态来适应低温,这与朱惠霞等[31]研究结果一致。光合指标的变化与植株自身的形态特性也密不可分,遭受低温胁迫后,两种不同类型冬油菜光合速率都降低,白菜型冬油菜陇油6号地上部分营养物质开始向地下部运输,而甘蓝型冬油菜2019-QL-GAU-30还在进行正常的营养生长。由于陇油6号地上部分向地下部运输的营养物质多,导致陇油6号的根冠比和根干质量大于2019-QL-GAU-30,更有利于保证正常的越冬。因此推断强抗寒性冬油菜通过调节光合特性来影响其营养物质的输送、储存方式,进一步影响其生长发育特性。
3.3 低温胁迫对冬油菜渗透调节的影响
低温造成植物伤害的最初部位是细胞膜的损伤[32],植物感受低温和其他胁迫后在体内合成大量的保护性物质来调节生理生化变化,使细胞渗透势保持平衡来维持细胞膜的稳定性[33-34]。研究表明可溶性糖、可溶性蛋白、游离脯氨酸的含量累积与抗寒性呈显著的正相关关系[35-36],本研究结果显示除叶片可溶性糖含量表现出先增加后减小之外,叶片和根部的其他可溶性物质均增加,这与蒲媛媛[7]、马骊[21]等研究结果一致,表明冬油菜通过增加渗透调节物质的含量来调节细胞渗透势,使细胞质的浓度升高,防止细胞内结冰,保证植株正常生长,但白菜型冬油菜陇油6号根部的可溶性物质含量均高于甘蓝型冬油菜2019-QL-GAU-30,说明陇油6号通过将营养物质运输到根部,增加根冠比和根直径保证安全越冬。通过两种冬油菜的对比,白菜型冬油菜陇油6号的叶片钙离子含量远大于甘蓝型冬油菜2019-QL-GAU-30,且根部钠离子含量大于叶片,推测冬油菜为了抵御低温,通过增加叶片钙离子含量保证冬油菜正常的光合作用和维持细胞内渗透压稳定,通过减少钠离子的含量、降低钠离子的毒害作用来维持冬油菜正常的生长发育。
4 结 论
本研究结果表明在越冬前遭受冷胁迫后白菜型冬油菜陇油6号的根冠比、累积的渗透调节物质含量、叶片积累钙离子含量、气孔导度和净光合速率都大于甘蓝型冬油菜2019-QL-GAU-30,而叶片钠离子的含量和胞间CO2浓度小于2019-QL-GAU-30,由此可见较大的根冠比、较高的渗透调节物质含量、钙离子含量和净光合速率可有效提高冬油菜的抗寒性,可为选育抗寒性强的甘蓝型冬油菜提供一定的理论依据。
猜你喜欢
今日农业(2022年1期)2022-11-16
电脑迷(2022年10期)2022-11-08
农业工程学报(2022年5期)2022-06-22
热带作物学报(2022年5期)2022-06-01
农技服务(2021年6期)2021-09-22
西藏文学(2020年5期)2020-09-21
快乐语文(2019年35期)2020-01-06
中国诗歌(2019年6期)2019-11-15
安徽农学通报(2016年24期)2017-01-12
董事会(2016年8期)2016-05-14
