
脱酚棉籽粕替代鱼粉对卵形鲳鲹肌肉品质的影响
2022-09-20 06:57苗玉涛钟宛清卢宝鑫付胜利陈义烘
华南师范大学学报(自然科学版) 2022年4期
苗玉涛, 钟宛清, 卢宝鑫, 付胜利, 宋 飞*, 陈义烘, 汪 蕾*
(1. 华南师范大学生命科学学院/现代水产养殖科学与工程研究院/广东省水产健康安全养殖重点实验室, 广州 510631;2. 南方海洋科学与工程广东省实验室(珠海), 珠海 519000; 3. 南方海洋科学与工程广东省实验室(湛江), 湛江 524025)
卵形鲳鲹(Trachinotusovatus)又名金鲳、鲳鲹,隶属于鲹科(Carangidae)。由于其生长速度快、肉质鲜美和环境适应力强的特点,在我国南方沿海地区得到了广泛的养殖。但在实际生产中,卵形鲳鲹饲料中需要添加大量鱼粉,导致其饲料价格昂贵,制约了该养殖业的发展。而且,由于全球气候变化、环境污染和过度捕捞,鱼粉的生产已经无法满足行业发展的需要[1-2]。因此,降低鱼粉在水产饲料中的使用比例对于实现该行业的可持续发展至关重要。
植物蛋白来源广泛且便宜,为取代鱼粉的理想蛋白源[1]。早在20世纪70年代中期,FOWLER和BANKS[3]就在鱼类饲料中使用小麦胚芽粉取代鱼粉。到目前为止,积极寻找其他合适蛋白替代鱼粉仍然是鱼类营养和饲料研究的热点[4],如在罗非鱼(Oreochromisniloticus)[5]、大西洋鲷(Sparusaurata)[6]等鱼类饲喂中添加多种植物蛋白代替部分鱼粉。研究[7]表明,饲料中添加少量植物蛋白对水产动物的生长性能、肌肉品质和免疫反应不会产生显著影响。但也有研究证实,饲料中添加植物蛋白会对养殖水产动物产生不良影响,带来了诸如蛋白质消化率低[8]、炎症[9]以及抗氧化力能力下降[10]等问题。各种植物蛋白对不同水产动物产生的影响并不一致,并且影响程度与添加量有一定关系,其中主要的影响因素、最佳添加量均有待深入研究。
棉籽粕(CSM)是一种重要的植物蛋白来源,被广泛用于不同鱼种的饲料。大量研究者在大菱鲆幼鱼[11]、西大西洋笛鲷(Lutjanuscampechanus)[4]等品种上进行了棉籽蛋白替代鱼粉的应用研究。但棉籽粕中含有的游离棉酚对鱼类有毒,限制了其在鱼类饲料中的大量添加。已有研究[12]证明,脱酚棉籽粕(LCSM)可被用于罗非鱼饲料中鱼粉的部分替代,但目前尚未有在卵形鲳鲹幼鱼中使用脱酚棉籽粕替代饲料中鱼粉的报道。因此,本研究通过解析脱酚棉籽粕替代鱼粉对卵形鲳鲹的肌肉品质、抗氧化能力及相关基因表达的影响,以探究脱酚棉籽粕替代不同比例的鱼粉在卵形鲳鲹养殖中的应用效果,为脱酚棉籽粕作为动物蛋白的应用开发提供理论支撑。
1 材料和方法
1.1 试验饲料
以鱼粉、脱酚棉籽粕、玉米蛋白粉、家禽副产品粉、豆粕和花生粕为主要蛋白源,以鱼油、大豆油和大豆卵磷脂为脂源,添加脱酚棉籽粕分别替代0%、20%、40%、60%的鱼粉,配制了4种等氮(质量分数约为42.5%的粗蛋白)和等脂(质量分数约为14.0%的粗脂肪)饲料,分别记为LCSM0、LCSM20、LCSM40、LCSM60组。饲料的一般成分如表1所示。将颗粒通过造粒机(F-26,中国广州华南理工大学),在45 ℃下风干,水的质量分数不超过10%。其中,LCSM0组为对照组,饲料制备结束后,置于-20 ℃密封保存。
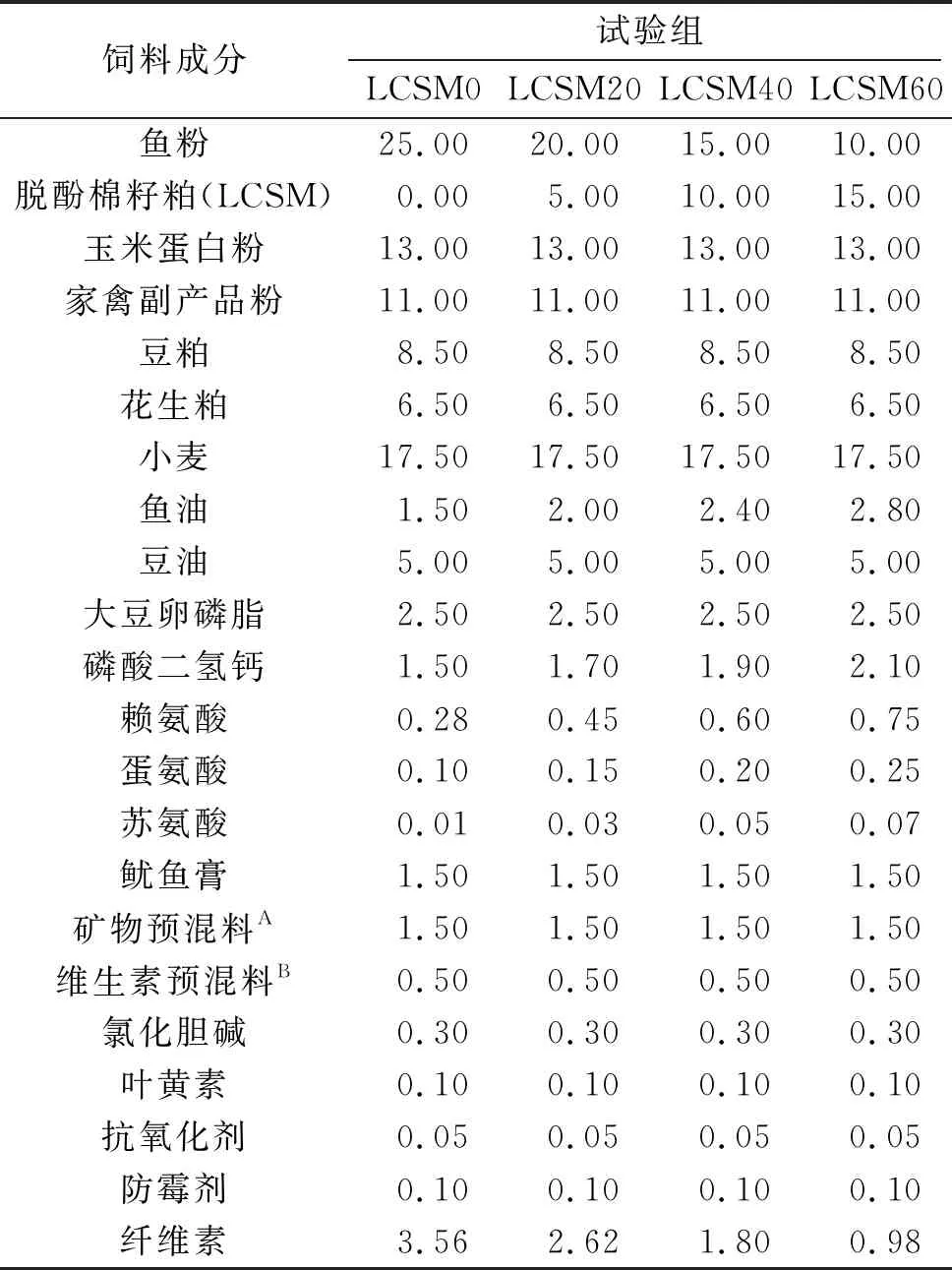
表1 试验饲料配方
1.2 试验动物和饲养管理
试验鱼购自阳西养鱼场(中国广东),喂食LCSM0组饲料2周以适应试验的饲料和条件。试验前禁食24 h,选取体质量为28.42±0.02 g的幼鱼随机分入网箱(1.5 m×1.5 m×1.5 m),每组3个重复。每天人工投喂2次(7:00和17:00),饱食投喂,连续6周。试验期间水温为28~30 ℃,盐度为15~17 g/L,pH为7.6~7.8,氨氮的质量浓度为0.05~0.1 mg/L,溶解氧的质量浓度为6~7 mg/L。
1.3 样品采集
养殖试验结束后,每个试验组取3尾鱼,切小块背肌(2 cm×2 cm×1 cm)冷藏,以用于肌肉质构的测定,另取3 g左右的肌肉在液氮速冻后-80 ℃保存待用。
1.4 肌肉质构测定
利用TA.XT.plus型物性测试仪(英国Stable Micro System公司)在质地剖面分析(Texture Profile Analysis,TPA)模式下,采用探头为圆柱形的P/50探头测试肌肉质构特性(硬度、胶着性、弹性、内聚性、胶粘性、咀嚼性、回复性)。质构仪使用参数设置为:圆柱状不锈钢探头,直径5 mm;测试前、中、后速度分别为1、2、5 mm/s;应变模式:下压距离为样品厚度的75%,二次压缩间隔时间为1 s;触发模式为自动,触发力为5 g。测试温度为常温。
1.5 肌肉理化特性测定
pH值测定:参照文献[13]的方法,采用直接测定法,称取鱼肉1±0.02 g,4 ℃冷冻研磨后用蒸馏水定容至8 mL,4 000 r/min离心10 min,取上清液用精密pH仪测定,3次重复,取平均值。
肌肉滴水损失率的测定及计算:每个试验组分别取3尾鱼背肌,用吸水纸吸干水分后,称取2±0.06 g肌肉(将此肌肉重量记为W0),置于离心管中(底部垫有适量吸水纸),于4 ℃、4 000 r/min离心20 min。取出肌肉,用吸水纸吸干肌肉表面水分,称取其重量,记为W1。计算公式为:
滴水损失率=(W0-W1)/W0×100%。
(1)
蒸煮损失率的测定及计算:每个试验组分别取3尾鱼背肌,用吸水纸吸干水分后,称取2±0.02 g肌肉(将此肌肉重量记为M1),置于离心管中,在80 ℃恒温水浴锅蒸煮10 min。取出后用吸水纸擦干表面水分,称取其重量,记为M2。计算公式为:
蒸煮损失率=(M1-M2)/M1×100%。
(2)
1.6 肌肉胶原蛋白的质量分数测定
肌肉胶原蛋白(Collagen)的质量分数采用南京建成生物工程研究所的试剂盒测定,具体测定方法参照试剂盒说明书。
1.7 抗氧化指标测定
MDA、SOD、GSH-Px含量采用南京建成生物工程研究所的试剂盒测定,具体测定方法参照试剂盒说明书。
1.8 RNA提取、cDNA合成及实时荧光定量PCR
按RNA提取方法提取肌肉总RNA,用分光光度计(Nanodrop 2000,美国Thermo公司)测定其质量浓度,用φ=1%的琼脂糖凝胶电泳鉴定其完整性。用PrimeScript反转录试剂盒将RNA定量反转录为1 μg cDNA,并将其保存在-20 ℃中。
采用SYBR Green染料(Takara),在Bio-rad CFX96上进行qRT-PCR。以β-actin基因为内参,分别检测肌肉中Decorin、MSTN、MyoG、Col1α1、CatB、CatL基因的表达,引物信息详见表2。采用Bio-Rad CFX Maestro检测阈值周期数(Ct),采用2-△△Ct方法对数据进行处理。

表2 试验所用引物序列
1.9 统计分析
2 结果与讨论
2.1 脱酚棉籽粕替代鱼粉对卵形鲳鲹肌肉质构指标的影响
质构是肌肉组织的重要物理特性,是评价鱼肉品质的重要指标。由表3可知:LCSM20组的卵形鲳鲹肌肉的硬度、弹性、内聚性、胶粘性、回复性与LCSM0组的相比差异无统计学意义,表明其肌肉紧实感、弹性与LCSM0组的相当;LCSM40组和LCSM60组的卵形鲳鲹肌肉的内聚性显著提高(P<0.05),其余质构指标显著下降(P<0.05),说明随着替换比例升高,肌肉弹性和嚼劲下降。综上说明,脱酚棉籽粕替换鱼粉的比例越高,其肌肉的硬度、胶着性、弹性、胶粘性、咀嚼性、回复性越小,内聚性越大,肌肉品质越差。
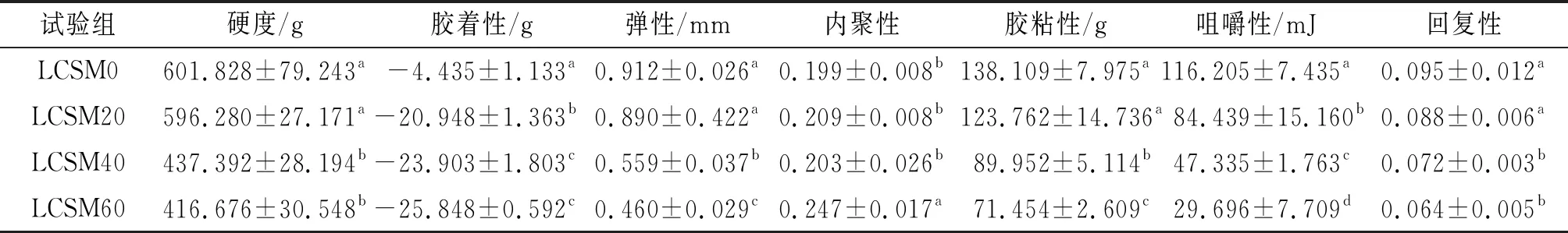
表3 卵形鲳鲹肌肉的质构指标
2.2 脱酚棉籽粕替代鱼粉对卵形鲳鲹肌肉理化特性相关指标的影响
肌肉的pH值与肉品的保水性、嫩度、色泽等有很大关系,直接影响肌肉的品质。当肌肉受外力作用时,保持原有水分和添加水分的能力叫做系水力,又叫保水力或亲水力,是肌肉评价的一个重要指标。系水力的高低直接影响肉的风味、颜色、质地、嫩度和凝结性。评定系水力的指标很多,有滴水损失率、蒸煮损失率和解冻减量等,本试验主要测定肌肉的滴水损失率和蒸煮损失率。由表4可知:4个试验组的pH值均低于7.0,其中,LCSM20组的pH值显著低于LCSM0组(P<0.05),说明pH值下降可增加鱼肉的嫩度和风味物质;与LCSM0组相比,LCSM20、LCSM40、LCSM60组的滴水损失率的差异无统计学意义(P>0.05),而蒸煮损失率都显著下降(P<0.05),表明鱼肉的失水率下降,即提高了系水力。张志勇等[14]研究发现投喂植物蛋白会降低花鲈和西伯利亚鲟的肌肉失水率,而姜俊等[15]研究发现投喂植物蛋白会升高建鲤的肌肉失水率,说明植物蛋白可能对不同鱼类的系水力影响有差异,其机制有待进一步研究。
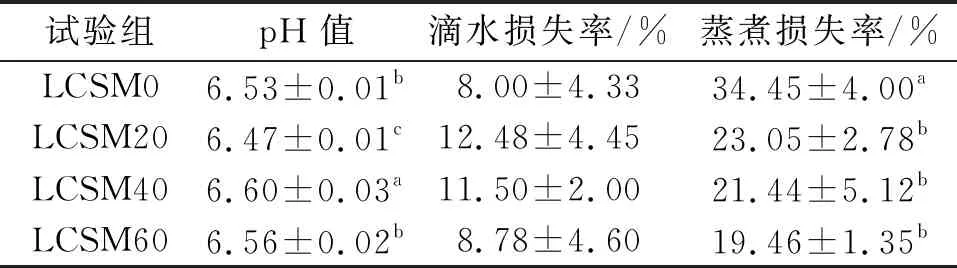
表4 卵形鲳鲹肌肉理化特性的相关指标
2.3 脱酚棉籽粕替代鱼粉对卵形鲳鲹肌肉中胶原蛋白的质量分数的影响
胶原蛋白主要分布于形成肌原纤维被膜的结缔组织中,胶原蛋白的质量分数越高,肌原纤维被膜的韧性越强。胶原蛋白可在肌纤维间和肌束周围形成致密的膜鞘,使肌肉具有较高的硬度和咀嚼性,从而影响肌肉的结构和功能特性。由图1可知:LCSM0组的卵形鲳鲹肌肉中胶原蛋白的质量分数为310.42 μg/g;LCSM20、LCSM40组的卵形鲳鲹肌肉中胶原蛋白的质量分数分别为290.11、287.19 μg/g,略低于LCSM0组的;LCSM60组的卵形鲳鲹肌肉中胶原蛋白的质量分数为229.66 μg/g,比LCSM0组的显著降低了26.02%。综合来看,当用脱酚棉籽粕替代20%或40%的鱼粉时,胶原蛋白的质量分数无明显变化,而替代比例达到60%时,胶原蛋白的质量分数显著降低。安玥琦等[16]研究表明,随着饲喂蚕豆时间的延长,草鱼肌肉中胶原蛋白的质量分数显著增加,其肌肉硬度和咀嚼性也显著增加。于晓彤等[17]等用混合植物蛋白质(豆粕和大豆浓缩蛋白)替代鱼粉饲喂草鱼和花鲈,发现:当替代比例为100%时,胶原蛋白的质量分数显著低于全鱼粉组的,硬度、黏性、咀嚼力及回弹力也显著降低。以上研究说明,肌肉中胶原蛋白的质量分数与肌肉硬度、咀嚼力及弹性呈正相关,胶原蛋白质量分数的降低是导致卵形鲳鲹肌肉硬度和咀嚼性显著下降的原因之一。
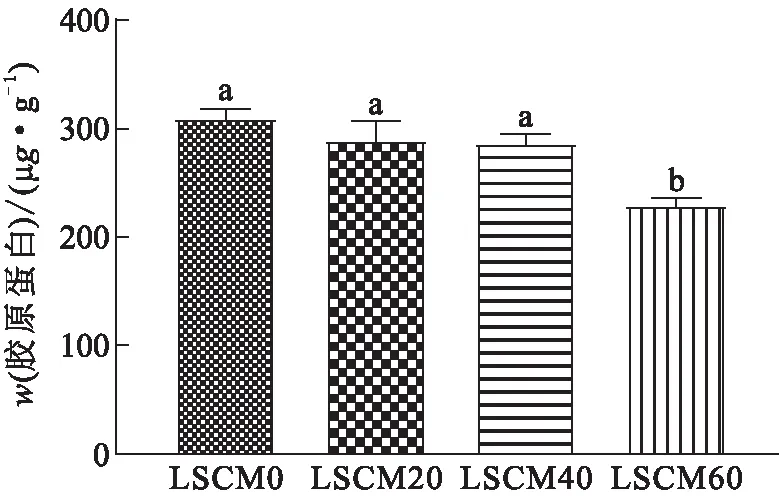
图1 卵形鲳鲹肌肉中胶原蛋白的质量分数
2.4 脱酚棉籽粕替代鱼粉对卵形鲳鲹抗氧化指标的影响
机体抗氧化防御系统可保护细胞和细胞膜免受氧化损伤,维持机体的正常生理功能。SOD和GSH-Px作为鱼类酶促抗氧化系统的主要成员,可与多种抗氧化酶相互协调,两者含量是指示机体内部抗氧化作用动态稳定程度的重要指标;MDA含量则是指示机体内脂质过氧化程度的常用指标。由图2可知:(1)各组之间肌肉的MDA含量的差异无统计学意义(P>0.05);(2)与LCSM0组相比,SOD含量随着脱酚棉籽粕替代鱼粉的比例的增加显著下降(P<0.05),其中,LCSM60组的SOD含量下降了34.94%,LCSM20组和LCSM40组的差异无统计学意义;(3)LCSM0组的GSH-Px含量与LCSM20组相比差异无统计学意义(P>0.05),LCSM60组的GSH-Px含量与LCSM0组相比显著降低(降低了41.79%)。说明用脱酚棉籽粕替代20%~40%的鱼粉时,对卵形鲳鲹机体的抗氧化性能影响不大,这与在黑鲷幼鱼饲料中用发酵棉籽粕替代鱼粉(替代比例低于16%)的研究结论[18]相似。在本研究中,当脱酚棉籽粕替代60%鱼粉时,机体的SOD含量和GSH-Px含量显著下降,抗氧化系统的防御能力降低,机体清除自由基的能力下降,易受到过氧化物的损伤。申建飞等[19]研究发现浓缩棉籽粕替代鱼粉的比例超过48%时,卵形鲳鲹抗氧化能力显著低于最适比例(26%)时的,这与本研究结果相一致。
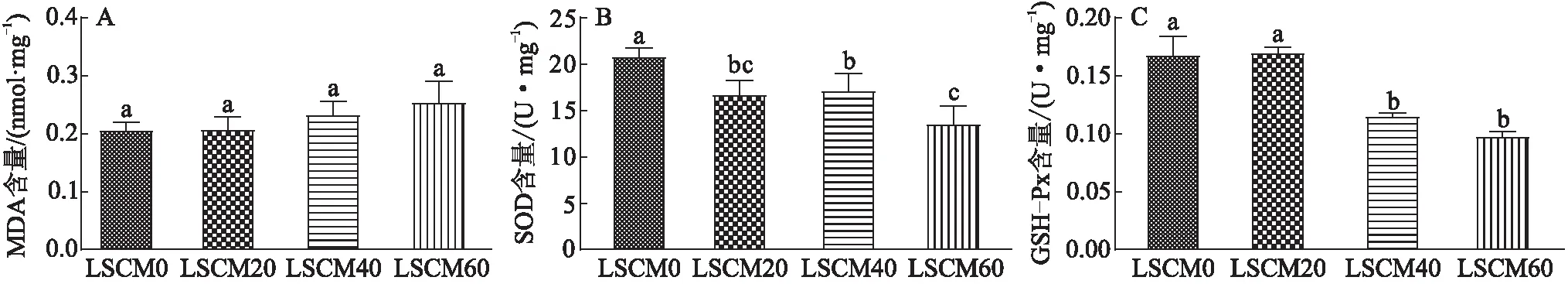
图2 脱酚棉籽粕替代鱼粉对卵形鲳鲹肌肉抗氧化指标的影响
2.5 脱酚棉籽粕替代鱼粉对卵形鲳鲹肌肉品质相关基因表达的影响
基因效应是影响肉质的内在因素。研究[20]表明,Decorin可通过抑制肌肉生长抑制素(Myostatin,MSTN)参与成肌细胞增殖分化过程的调控,促进肌肉的生长发育。但关于Decorin调控MSTN的表达在鱼类肌肉的研究较少见报道。肌肉生长抑制素(Myostatin,MSTN)又称为GDF-8(Growthdifferentiationfactor8),是转化生长因子β(Transforminggrowthfactor-β,TGF-β)家族成员。XU等[20]发现:畜禽类的肌肉量在敲除MSTN后可显著增加,表明MSTN在生肌过程和肌肉生长中起重要的抑制作用。XIE等[21]在研究禁食和复食对长臀鮠(Cranoglanisbouderius)的MSTN基因表达的影响中发现:肌肉中MSTN的mRNA表达水平在饥饿过程中逐渐降低,再喂养后恢复到正常水平。李卓钦等[21]发现:肌细胞生成素(Myogenin,MyoG)主要负责肌肉分化,MSTN通过启动下游MyoG的反应,阻碍肌细胞的生长。
由图3A-C可知,肌肉中Decorin、MSTN、MyoG基因的相对表达量受到脱酚棉籽粕替代鱼粉的比例的影响:与LCSM0组相比,在脱酚棉籽粕替代20%~40%的鱼粉时,Decorin的相对表达量的差异无统计学意义,LCSM60组的显著下降了19.98%;Decorin下游的MSTN在LCSM20组的相对表达量与在LCSM0组的差异无统计学意义,在LCSM40组和LCSM60组则显著上升(P<0.05),与Decorin的相对表达量呈现相反的趋势;MSTN过表达会阻碍动物机体骨骼肌的生长,本研究中MSTN的相对表达量随着脱酚棉籽粕替代比例的增加而升高,肌肉品质相关指标下降,说明MSTN的基因水平调控着卵形鲳鲹幼鱼的生长,机体生长受到抑制;MyoG的相对表达量在各组之间的差异无统计学意义。李卓钦等[22]发现:MSTN的相对表达量降低、其下游的MyoG的相对表达量升高可能会导致斑点叉尾鮰(Ictaluruspunctatus)体质量的增加和肌肉中脂肪含量的减少,这与本研究结果不一致。MSTN可以通过抑制MyoD家族成员的转录活性负向调控肌细胞的生长发育,除了MyoG之外,MyoD家族成员还包括成肌细胞决定蛋白(MyoD)、肌源性因子5(MYF5)、肌源性因子6(MYF6)和肌细胞特异性增强因子2C(MEF2C)等。ZHANG等[23]通过敲除山羊成纤维细胞的MSTN,发现突变山羊的MSTN的mRNA相对表达量显著下降,继而检测结果表明MyoD、MYF5、MYF6、MEF2C基因的相对表达量均显著上升,说明MSTN水平降低能够增强4种肌源性转录因子的表达,从而促进生长。由本研究的结果可以看出,Decorin和MSTN影响卵形鲳鲹幼鱼的生长,但影响MSTN和MyoG表达的确切机制尚不清楚,可能是MSTN可能通过负调控MyoD家族成员其他转录因子来调节肌肉的生长,有待进一步探究。
许晓莹等[24]证实鲤鱼(Cyprinuscarpio)、斑马鱼(Barchydanioreriovar)、鲟鱼(Acipensersinensis)和日本鳗鲡(Anguillajaponica)肌肉中主要为COL1α1编码的Ⅰ型胶原蛋白。本研究发现,LCSM20、LCSM40、LCSM60组的COL1α1的相对表达量与LCSM0组相比显著升高(P<0.05),其中LCSM20组和LCSM40组的差异无统计学意义,LCSM60组的相对表达量约为LCSM0组的1.92倍(图3D)。李迪等[25]研究发现草鱼(Ctenopharyngodonidella)×眼鳟(Squaliobarbuscurriculus)杂交F1的COL1α1的相对表达量显著高于其母本的,肌肉硬度与弹性也显著高于其母本的;许晓莹等[25]研究发现,使用杜仲饲喂草鱼4周后,其肌肉的COL1α1、COL1α2基因的相对表达量在短期内低于LCSM0组的,胶原蛋白的质量分数高于LCSM0组的。本研究同样发现,在用脱酚棉籽粕替代鱼粉之后,卵形鲳鲹的COL1α1的相对表达量升高,而胶原蛋白的质量分数降低,具体作用机制有待进一步研究。
组织蛋白酶主要存在于溶酶体中,当其从溶酶体中释放出来,能够破坏肌肉蛋白完整性,加快肌肉的软化。本研究发现CatB在LCSM40组的相对表达量与LCSM0组相比差异无统计学意义(P>0.05)(图3E),CatL在LCSM20组和LCSM40组的相对表达量与LCSM0组相比差异无统计学意义(P>0.05)(图3F),而CatB和CatL在LCSM60组的相对表达量均是LCSM0组的2.12倍(图3E、F)。以上研究结果表明:随着脱酚棉籽粕替代鱼粉的比例的增加,组织蛋白酶的相对表达量提高,蛋白质加速降解,从而导致肌肉硬度的下降。已有研究[26]表明,饲料中添加适宜水平苏氨酸均能降低建鲤(Cyprlnuscarpiovar Jian)肌肉中CatB和CatL的相对表达量,减少肌原纤维的断裂,从而明显改善了肌肉的硬度品质,说明CatB和CatL的相对表达量与硬度呈现负相关,这与本研究结果较一致。
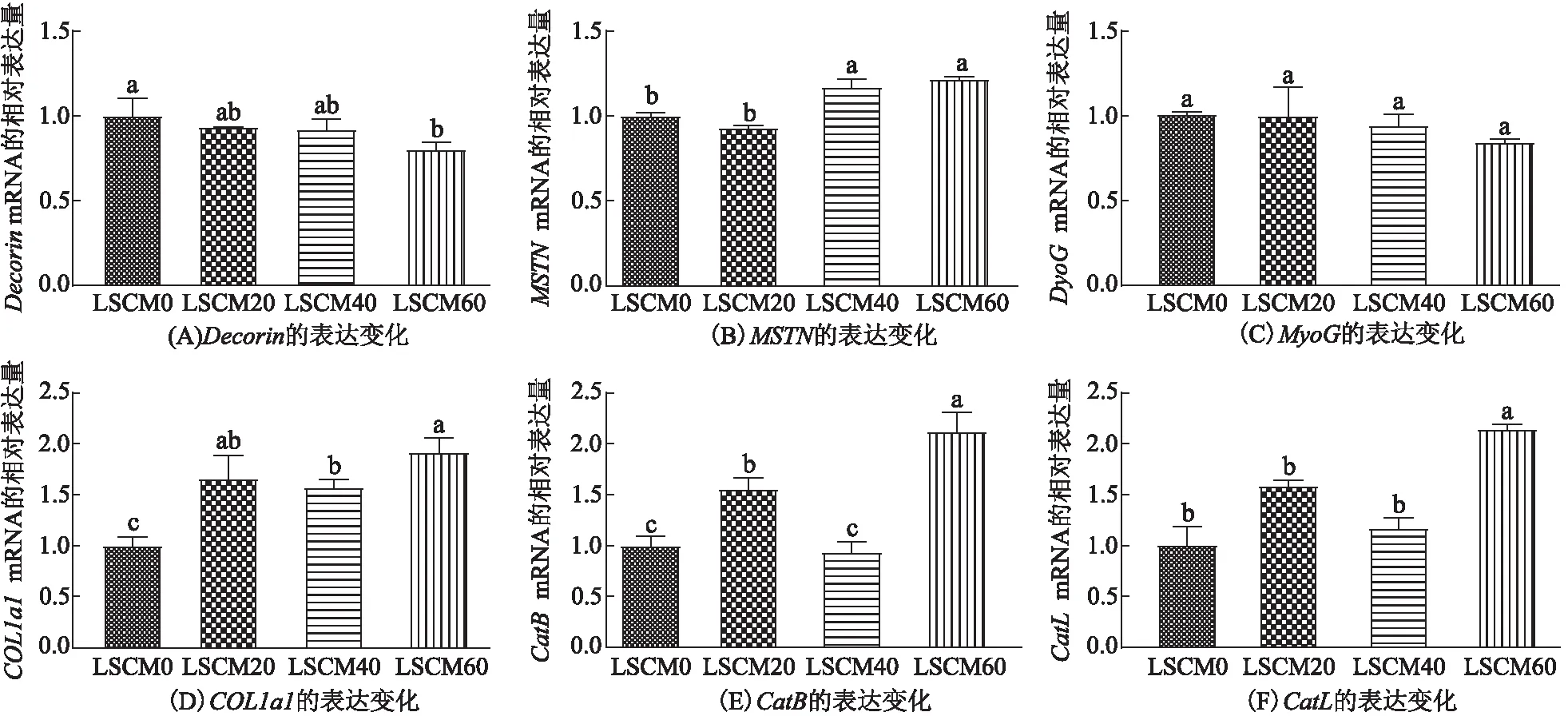
图3 肌肉相关基因Decorin,MSTN,MyoG,COL1a1,CatB和CatL的表达变化
3 结论
本研究探究了饲料中用脱酚棉籽分别替代0%、20%、40%和60%的鱼粉对卵形鲳鲹的肌肉品质、抗氧化能力和肌肉相关基因的影响。结果显示在卵形鲳鲹幼鱼饲料中,利用脱酚棉籽粕替代20%鱼粉时,不会降低肌肉品质:饲料中以脱酚棉籽粕替代20%~40%鱼粉,对卵形鲳鲹的肌肉品质无显著影响;替代比例为20%时,对卵形鲳鲹肌肉的MDA、GSH-Px含量无影响;随着饲料中脱酚棉籽粕替代鱼粉的比例的增加,组织蛋白酶的相对表达量提高,蛋白质加速降解,影响肌细胞合成,从而导致肌肉硬度的下降,但当替代比例为20%时则无影响。综上所述,在卵形鲳鲹养殖中,建议脱酚棉籽粕替代鱼粉的最适比例为20%。
猜你喜欢
水产科技情报(2022年5期)2022-11-25
当代水产(2022年8期)2022-09-20
——奇妙的蛋
趣味(数学)(2022年5期)2022-07-29
当代水产(2022年6期)2022-06-29
当代水产(2021年4期)2021-07-20
新农业(2021年9期)2021-06-20
老年教育(2021年1期)2021-02-02
——全棉籽的加工与利用
中国畜牧杂志(2020年8期)2020-12-18
当代水产(2020年4期)2020-06-16
西藏人文地理(2019年2期)2019-03-26
