
基于双向长短时记忆神经网络的步态时空参数脑肌电解码方法
2022-09-20 06:48魏鹏娜马鹏程张进华洪军
西安交通大学学报 2022年9期
下肢外骨骼机器人是可用于帮助运动功能障碍患者行走和恢复运动功能、提高老年人运动能力、增强体力劳动者和士兵负载强度和耐力的可穿戴人机一体化辅助装置
。与智能假肢不同,外骨骼需要与人体融合,感知人体的运动意图,从而以一定的方式配合穿戴者完成特定动作
。对于下肢外骨骼而言,步态的分析和识别是其完成人体运动意图感知和预测的关键环节。对穿戴者运动意图的精确感知,是外骨骼配合穿戴者完成目标运动的首要条件,也是解决人机协调问题的关键技术
。患者通过自身意图对外骨骼进行控制,一方面可以提高患者的参与度和积极性,另一方面可以促进大脑的可塑性
。目前,用于下肢外骨骼的步态信息形式包括:足底压力、加速度、关节运动角度
、脑电(electroencephalography,EEG)、表面肌电信号(surface electromyography,sEMG)等
和多模态信号融合等
。基于步态相位和运动模式识别
的离散控制方式,存在运动不平稳、步态连续性差等问题,为了实现对外骨骼步态的连续、平滑控制,步态轨迹的连续解码对下肢外骨骼的发展至关重要。
(1)由于冷凝式热水器内部存在系统热容会导致热水器内管路对热量有蓄热的作用,热量传递给水在时间上发生延迟,因此热水温升与加热时间呈非线性增长趋势,热水器系统热量分布可由 表示,热水温升与加热时间关系数学模型由表示。热水器加热时间的影响因素包括系统热容、进水流量以及进气流量。
目前对连续步态轨迹解码的研究有基于EEG信号、sEMG信号和关节运动学数据等,比如休斯顿大学的Nakagome等
利用EEG信号对下肢关节的运动角度进行解码。杜克大学利用恒河猴的侵入式脑电信号对下肢关节三维运动位移、肌电信号、步长等运动信息进行解码
。哈尔滨工业大学的研究人员利用sEMG信号和惯性测量单元的数据对下肢关节运动角度等进行预测
。然而,基于EEG信号的步态运动轨迹预测相关性较低,难以达到实际应用的程度。侵入式脑电信号采集方式对人体伤害性较大,难以广泛应用。基于sEMG和惯性测量单元数据的步态预测,由于惯性测量单元所测量的数据属于运动学数据,将其与sEMG信号融合对运动学数据进行预测,很难评价sEMG的解码能力。
本研究设计双向深度长短时记忆神经网络模型(bidirectional long short term memory,BiLSTM),利用深度学习模型自动提取运动相关频段sEMG和EEG的特征,实现对膝关节和髋关节运动时空参数的连续解码,实现了基于双信号模式的,下肢关节运动时空参数的连续精确解码;利用sEMG和EEG的相关关系,设计基于BiLSTM的EEG解码sEMG信号的神经网络模型。
1 BiLSTM神经网络结构设计
LSTM是在递归神经网络(recurrent neural network,RNN)的基础上发展而来,该算法是为了解决RNN梯度消失和梯度爆炸的问题。通过选择性记忆和忘记,LSTM可解决对有时序关系的时间序列的长时记忆问题。下肢运动中的sEMG和EEG信号属于有序时间序列,可采用LSTM对其进行分析
。LSTM神经网络一个神经元包含网络的当前输入信号
(该信号在本文中为EEG和sEMG融合信号或单独的EEG信号),上一时刻隐含状态
-1
,遗忘门控信号
,选择性记忆门控信号
,输出门控信号
,新的输入信号
。3个门控信号是由
和
-1
的拼接矩阵乘以权重再通过
激活函数转换为0到1之间的数值,其计算方法分别如式(1)~(3)所示,新的输入信号
由
和
-1
的拼接矩阵乘以权重再通过tanh激活函数转换为-1到1之间的数值,计算方法如式(4)所示
=
(
[
,
-1
])
(1)
式中:
和tanh是激活函数;
、
、
、
是权重矩阵;
为当前输入的用于解码的信号;
-1
为上一时刻隐含状态。
(2)
=
(
[
,
-1
])
(3)
=tanh(
[
,
-1
])
(4)
=
(
[
,
-1
])
CFRP在轨道交通车辆车体上的推广应用是复合材料行业的一大机遇和挑战,虽然目前受制造成本、成型工艺、生产效率等方面的制约,难以在短期内实现批量生产,但是全碳纤维复合材料地铁车辆车体的成功研制经验为后续轨道交通复合材料承载结构的研究奠定了基础。随着新材料、新技术日新月异的进步,相信在不久的将来,中国将迎来轨道交通车辆车体新材料应用的又一次变革。
=
(
′
)
4.在课后组织方面——重视课堂小结。由于同层次学生对知识的学习掌握能力不同,教师要定期总结,帮助学习能力不强的学生跟上教师的讲课进度。
=
⊙
-1
+
⊙
(5)
大港油田积极响应中国石油“稳健发展”和“保持一亿吨以上原油产量规模”的发展战略,坚持把提高采收率增加老油田可采储量作为油气开发的核心价值观,规模开展高孔高渗、高温高盐、深层低渗三大类油藏的开发试验,大幅度提高老油田采收率,努力把资源优势转化为产量优势和发展优势,努力实现产量稳定和可持续发展。
人类的下肢运动意图由大脑运动相关皮层发起,由脑干控制执行过程,受小脑、丘脑和基底神经节的调节,由大脑皮层产生的动作电位通过神经-肌肉接头传入骨骼肌中并形成终板电位,当终板电位达到一定阈值后,会在肌细胞中产生强大的肌细胞动作电位,随着动作电位的传播,肌浆网释放钙离子,根据肌丝滑行理论,细肌丝沿着粗肌丝滑行,实现肌肉收缩,完成相应的动作,如图3所示,为运动的产生和执行通路。在运动意图产生和传播过程中,所产生的EEG和sEMG信号真实记录了整个运动过程的发生。因此,为了全面描述步态的真实过程,下肢运动实验对匀速行走时,运动皮层EEG、运动相关肌肉sEMG和下肢关节三维运动轨迹进行同步采集。EEG和sEMG信号的同步,通过Brain Products公司的无线32导运动状态下脑肌电同步采集设备,该设备是为运动中EEG和sEMG信号采集所设计,并通过设备自带的同步核实现和Vicon系统的同步,从而实现对下肢运动学数据的同步采集。
=
⊙tanh(
)
(6)
LSTM的执行过程主要包括以下几个步骤:首先,是遗忘阶段,对传入的上一时刻的细胞状态
-1
通过遗忘门
进行选择性忘记,记住重要的信息,对不重要的信息进行遗忘。其次,是选择性记忆阶段,首先对网络的当前的输入信号
和网络的上一时刻的隐藏状态
-1
进行拼接,乘以权重并通过tanh激活函数转换为新的输入信号
,再通过记忆门控单元
进行选择性记忆,记住重要的输入信息。前两个阶段的计算如下式所示
(7)
式中:
和tanh是激活函数;
是权重矩阵;
为当前时刻细胞状态。当前状态的输出
由当前隐含状态
计算得到,计算过程如下式所示
式中:
-1
为上一时刻细胞状态;
、
、
分别为遗忘、选择性记忆和输出门控信号;
为新的输入信号;⊙运算表示对两个矩阵中的对应元素进行相乘操作。
=
(
′
)
(8)
式中
保存了当前时刻隐含状态,具有短期记忆功能。
综合了当前输入和前一时刻的状态信息,可保存长期的状态,因此该神经网络被称为长短时记忆神经网络,LSTM神经网络一个神经元的结构如图1所示。


LSTM神经网络利用过去的信息进行信号的分析,而BiLSTM神经网络在LSTM的基础上加入未来的信息。BiLSTM的运算如图2所示。在前向计算的基础上加入反向计算(不是反向传播),将未来的信息加入当前状态的计算,比如
+1时刻的信息
+1
传入
时刻,参与该时刻细胞状态的计算。本文将建立面向步态时空参数解码的BiLSTM神经网络模型,对膝关节和踝关节的步态时空参数进行解码。LSTM神经网络模型的结构包括一个输入层、两个LSTM层、两个全连接层和一个输出层,其中输入层包含的节点个数和输入的信号特征个数相等,每个LSTM层包含一个dropout层,参数设置为0.5,即随机丢弃概率为0.5,其作用是防止数据过拟合。为防止梯度爆炸,将梯度阈值设置为1,LSTM层的隐含单元个数为200,训练样本最大迭代次数设置为100,采用adam优化器对训练过程进行优化,学习率即步长设置为0.01,并设置验证集,对训练过程进行校验。由于研究的数据为连续的信号对连续运动轨迹的解码,因此选择序列到序列(sequence-to-sequence)的回归方式。BiLSTM神经网络模型在LSTM结构的基础上增加了BiLSTM层,隐含单元个数为200。
一是反腐职责与保护地方经济的矛盾。腐败的主要表现形式为官权与商业资本的联合,并且腐败案件涉及的商业项目资金数额都较为庞大,若依照程序严惩腐败分子,则与腐败案件关联的商业项目必然随之枯萎,地方经济遭受损失。反之则是容忍腐败分子侵吞公共财产,扰乱市场秩序。在反腐工作与经济保护之间的价值选择成为阻碍反腐绩效的矛盾之一。二是价值与执行相脱节。在复杂的利益纷争下,公共部门所推崇的政治价值、管理价值等基本公共价值在官僚系统运行过程中容易走样,无法真正指导和落实到执行过程。
2 步态时空参数的脑肌电解码
2.1 下肢sEMG和EEG信号采集实验范式
最后,是输出阶段,决定哪些信息将输入下一个状态,将当前细胞状态经过tanh激活函数转换后通过门控信号
进行控制,计算过程如下式所示

实验招募了7名来自西安交通大学的健康研究生,平均年龄为24岁,由于本次实验为短期实验,短期实验对象个数为7~12,因此本次实验被试人数在合理范围内。实验所有程序均经西安交通大学伦理委员会批准,且符合1975年赫尔辛基宣言的相关规定,实验前所有被试均签署了知情同意书。在进行步态相关实验时,采集EEG和下肢肌肉sEMG信号时,每位被试按照自己日常习惯行走,尽量保证步态协调,不要产生过大的加速度,特别是在步态实验中,被试除了下肢运动外,身体其他部位尽量不要做过多的运动,尽量减少肌电的干扰。比如,尽量不要做转头、低头等动作,以减少颈部肌肉对EEG信号的影响,同时尽量避免频繁眨眼,以减少眼电干扰。在实验过程中,被试尽量减少咀嚼、吞咽动作和夸张的面部表情等,并保持注意力集中,以免在其他大脑皮层产生与运动无关的EEG信号,干扰运动区的EEG信号。脑电帽佩戴要符合规范,紧贴头皮,以防止脑电帽电极的滑动引起EEG信号的干扰。每位被试在跑步机上以自己舒适的速度行走,经过7名被试的反馈,最终综合确定最佳速度为2.0 km/h。
为采集与下肢运动最相关肌肉的sEMG信号,通过查阅相关的国内外文献和欧洲发布的用于非侵入式肌肉评价的sEMG信号采集计划(surface electromyography for the non-invasive assessment of muscles,SENIAM)(www.seniam.org)的内容,本实验对左右腿的股二头肌(biceps femoris,BF)、股内侧肌(vastus medialis,VM)、内侧腓肠肌(gastrocnemius medialis,GM)和胫骨前肌(tibialis anterior,TA)8块肌肉的sEMG信号进行采集,单腿4块肌肉的作用如表1所示,肌肉位置如图4所示。
试验设置2个组,配合饲料组和血液组,每个组3个重复。养殖密度为40尾/箱。试验组采用自制配合饲料灌入风干猪血肠衣中投喂,对照组采用鸡血打成血浆灌入风干猪血肠衣中投喂,养殖周期60 d,投喂频率为每3 d 1次,每天换水量50%,停饲1周采样。


实验过程中,首先,被试在平地上站立30 s,采集静息态下的EEG、sEMG信号和关节的运动数据,用于运动学模型的校正。其次,等待跑步机以2.0 km/h的速度运行正常后,被试在跑步机上匀速行走30 s为一组,每组中间休息10 s,采集15组后,完成整个步态信息采集实验。保证整个实验中对时间严格要求,被试状态良好,注意力集中。
2.2 实验平台搭建
根据设计的下肢运动实验范式,搭建下肢运动数据采集实验平台。实验台硬件设备主要包括:Brain Products公司的32Ch LiveAmp脑电帽、LiveAmp无线放大器和同步核、三维步态分析系统Vicon T40S、跑步机和计算机等;数据采集软件包括:Recorder脑肌电数据采集软件、Analyzer 2.0脑肌电数据分析软件、Nexus 2.6下肢关节运动轨迹捕捉与数据处理软件。
实验采用的无线脑肌电同步采集系统由脑电采集帽、肌电采集电极、无线放大器和同步核组成,脑电帽的电极位置按照10-20国际标准分布。LiveAmp放大器是针对运动EEG信号采集设计的,LiveAmp的记录特性可与高端实验室脑电图系统相比,24位A/D转换,可同时对EEG和sEMG信号进行采集,内置三轴加速度计,可用于后续对运动伪迹的去除。实验开始前,被试者头戴脑电帽,脑电帽的绝缘电缆线一端与脑电帽相连,另一端与信号放大器相连,放大器与计算机USB接口相连,并通过其供电和传输数据,不需要任何的特殊接口或特殊电源供应。计算机上的Recorder采集系统实时采集并保存被试24导EEG信号(Fp1,Fp2,F3,F4,C3,C4,P3,P4,O1,O2,F7,F8,T7,T8,P7,P8,Fz,Cz,Pz,Oz,M1,M2,FPz和VEOG,其中VEOG为水平眼电信号)和左右腿共8个肌肉通道的sEMG信号,采样频率为500 Hz。分别将Vicon反光球贴在左和右下肢的髂前上棘、髂后上棘、大腿、膝关节、胫骨(小腿)、踝关节、脚后跟、脚趾共16个位置上,采用10个摄像头捕捉反光球的运动轨迹,采样频率为100 Hz。
这天下午,易非陪着妈带着李倩倩去产检,李倩倩刚进到B超室,妈就在外面唠叨:“这孩子是怀上了,不知道能接进来不?”她指的还是房子。
2.3 sEMG和EEG信号预处理
由于实验中采用的LiveAmp无线放大器和三维步态分析系统Vicon T40S的采样频率不一致,同时考虑到增加序列长度(单个步态阶段的数据点数)可以丰富脑电信号或表面肌电信号的特征信息,因此将重采样频率设为1 000 Hz。实验中采用的电极帽,其参考电极位于额叶区和中央区之间,不符合参考电极的标准:参考电极的活动尽量降到最低,以获得最真实的基线(近似零)信号。因此,FCz通道不适合作为参考信号,本文选择双侧耳后乳突M1和M2的平均值作为新参考。眼球运动会改变眼睛周围电场分布,而眼睛又十分靠近头皮,所以对头皮表面电场分布影响很大。眼动伪迹频率范围与EEG频率重叠,且幅值较大,是EEG采集中的一种主要干扰。独立成分分析(independent component analysis,ICA)分析方法可用于从多个源信号线性混合的信号中分离出源信号,算法首先会找到一个映射,使混合信号通过该映射后投影分量在时间上重叠的最小,因此引入眼电相关ICA对初始信号中的眼电进行分离。下肢运动带来的运动伪迹,对EEG的影响较大,可利用EEGLAB工具箱中的伪迹子空间重构(artifact subspace reconstruction,ASR)算法进行处理。ASR将主成分分析应用于EEG数据,识别出与包含最小运动伪影的基线数据显著偏离的EEG通道,再与基线数据相比,方差高于预定义阈值的通道被识别为受干扰的通道,并使用从基线数据计算的混合矩阵进行重构
。经过以上分析,可分别去除和眼电、运动相关性最高的成分
。
2.4 sEMG和EEG信号运动相关特征提取

运动相关的频段主要分布在低频部分。Presacco等的研究表明
,运动学信号频率小于3 Hz部分占总功率谱密度的90%,因此对sEMG、EEG和运动信号进行1~3 Hz的带通滤波。事件相关谱扰动(event related spectral perturbation,ERSP)
可对EEG信号进行时频分析,利用ERSP对行走时EEG信号的能量谱进行分析,验证EEG用于步态解码的可行性。如图5所示,图5(a)为一个步态周期C3通道EEG信号ERSP分析结果,图5(b)为滤波后EEG、sEMG信号和踝关节运动轨迹。从图5(a)中可观察到,一个步态周期EEG信号的1~3 Hz频段(红框部分)包含充分的信息,比如在步态周期前50%,有两个时段大脑活动受到抑制,说明在这两个时段大脑参与行走运动
。从图5(b)中可观察到,滤波后sEMG信号和EEG信号中出现部分与踝关节运动相关的波形,特别是sEMG信号的波形与踝关节的运动轨迹相关的波形更多。

sEMG、EEG和运动信号信号特征提取和解码器训练过程如图6所示。首先对右腿4个肌肉通道的sEMG信号、21个通道的EEG信号和右脚踝、膝关节的运动轨迹进行带通滤波,滤波频率为1~3 Hz,然后将sEMG和EEG信号和关节运动数据分别分为训练集、测试集和验证集,3个数据集的比例为6∶2∶2。由于下肢运动是由拮抗肌肉的激活产生的,因此需要提取能够代表这种拮抗作用的特征,以提高步态的连续解码精度。为了解决这一问题,采用BiLSTM神经网络自动提取sEMG和EEG信号的运动相关特征。在所有的深度学习算法中,BiLSTM由于增加了遗忘门,可以对记忆的信息进行选择性遗忘,从而对长时依赖信号的分析方面有显著优势。为了验证该算法在利用EEG和sEMG信号对步态解码的优势,本文将对采用BiLSTM神经网络和支持向量机神经网络(support vector machine,SVM)
的步态解码结果进行对比分析,SVM分类算法可实现对离散动作的识别,回归算法可实现对连续轨迹的拟合,是一种常用的生物电信号分析机器学习算法。本文采用BiLSTM (200-100)对非平稳sEMG和EEG信号进行特征提取,数据的标签设置为下肢关节运动位移数据,在误差反向传播阶段,需要根据预测结果和标签的数据之间的误差对训练过程进行调节。
3 步态时空参数解码结果
3.1 基于sEMG和EEG的步态时空参数解码
对于多种模式信息的融合,主要有数据层、特征层和决策层3种方式。数据层的融合方式可以最大程度的保留各种融合信号的信息;特征层融合方式是在提取每种融合信号的特征后,将融合后的特征输入判别器进行分析;决策层的融合方式是对每种信号的判别结果采用特定规则进行融合。由于BiLSTM神经网络模型的特征提取由网络自动进行的,不需要采用特征层融合方式,但为了和数据层融合策略的解码结果进行对比,提取sEMG和EEG信号的平均绝对值(mean absolute value,MAV)特征并进行融合作为BiLSTM神经网络输入信号,决策层融合方式丢失信息过多,因此本次研究不进行分析。本文采用数据层的sEMG和EEG融合方式对步态时空参数进行解码,将右腿4个肌肉通道的sEMG信号和21个通道的EEG信号,组成数据矩阵输入BiLSTM神经网络模型中,在模型中自动进行特征提取和回归分析。
sEMG和EEG融合信号的膝关节和踝关节运动轨迹解码结果和模型训练过程中误差的变化如图7所示,随着模型迭代次数的增加,模型误差RMSE逐渐减小,最终趋于稳定,说明该模型对步态解码效果较好,迭代次数选择较合理。从解码的轨迹和实测轨迹的对比中,可看到sEMG和EEG融合信号解码的轨迹与实测信号轨迹虽然形状较相似,但解码出的轨迹幅值存在一定误差。可能的原因是sEMG和EEG信号幅值的差异较大,导致解码出的运动轨迹形状相似性较高,但幅值在波峰和波谷处存在误差,后续研究可在通过对信号进行归一化处理或者针对不同类型信号引入调节参数进行处理。sEMG-EEG融合信号解码轨迹和实际轨迹的Pearson相关系数
和均方根误差RMSE如表2所示。从表中可看到,sEMG-EEG融合信号的解码轨迹和实际轨迹的Pearson相关系数
超过0.9,说明他们形状相似度非常高,本文所提的步态时空参数解码策略可成功解码出下肢膝关节和踝关节的运动参数。

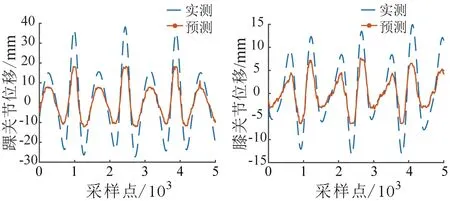
为了证明本文中所提sEMG-EEG融合信号对步态时空参数解码的效果,分别对单独的sEMG信号、EEG信号和sEMG-EEG特征融合信号的步态轨迹解码进行分析,对踝关节轨迹解码结果如图8(a)、(b)和(c)所示。此外,对sEMG-EEG融合信号在分别采用BiLSTM神经网络和SVM神经网络的步态时空参数解码进行分析,结果如图8(d)所示。图8中4种方法解码轨迹的Pearson相关系数
和均方根误差RMSE如表3所示。图8中的结果表明,采用单独的sEMG信号可解码出波峰处的轨迹(比如采样点1 000左右),但对次波峰处的解码轨迹误差较大(比如采样点300左右)。采样单独的EEG信号和sEMG-EEG特征融合信号的步态轨迹解码形状误差较大,从表3中可看到,二者解码出的轨迹
值都低于0.5。采用SVM回归算法从sEMG-EEG融合信号中解码出的踝关节轨迹在波峰和波谷处出现高频振荡,形状相似性
值为0.85±0.02,与BiLSTM神经网络的解码结果相比,解码轨迹的形状相似性较低。综上所述,本文所提出的基于sEMG-EEG融合信号的BiLSTM神经网络步态时空参数解码策略效果最好。
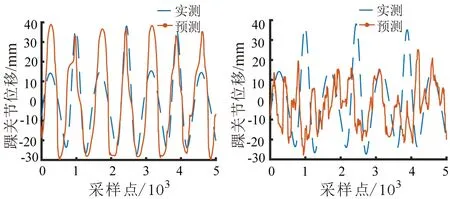

3.2 基于EEG的行走时sEMG信号解码
行走时大脑皮层和下肢肌肉之间生理层面的相关性,为从EEG信号中解码sEMG信号提供了理论基础。BiLSTM神经网络模型结构及参数设计与上述研究中的相同,将21个EEG通道信号作为BiLSTM神经网络模型输入,左腿VM肌肉通道的左腿VM肌肉通道的sEMG信号作为标签数据,对模型进行训练,以被试1为例对解码结果进行分析,其解码结果如图9所示。图中实测的曲线为实测sEMG信号经过1~3 Hz滤波后的波形图,可观察到在前2 000个采样点中,实测与解码的波形图形状相似性较高,2 000~2 500这段解码波形出现两个波峰,实际波形只有一个波峰,其他时间段也存在类似的情况,整段解码波形和实际波形存在一定误差。解码波形与实际波形的平均Pearson相关系数
=0.45±0.02,证明了形状相关性较低,可能是因为EEG信号中包含的步态信息不足以解码出sEMG信号中的步态成分(参考图5(b))。后续可以通过设计新的实验方案以增加EEG信号中步态信息,比如在被试行走过程中增加行走运动想象。解码波形的平均RMSE为2.03±0.32,和解码关节运动轨迹时的RMSE值相比没有增加很多。这可能是因为sEMG信号的幅值相对关节运动的位移较小,因此尽管预测波形的误差较大,但其RMSE值依然较小。

4 总 结
本文通过构建基于BiLSTM神经网络模型的下肢膝关节和踝关节的运动轨迹连续解码模型,将sEMG-EEG融合信号作为神经网络的输入信号,解码轨迹与实测轨迹形状相似性较高(膝关节
=0.94±0.04,踝关节
=0.97±0.02),可实现对下肢膝踝关节运动参数的解码。与传统的SVM模型相比,对踝关节解码轨迹与实测轨迹形状相似性Pearson相关系数提高了0.12;所提出的脑肌电融合方法比单独的脑电、表面肌电信号和脑肌电信号平均绝对值特征融合信号相比,对踝关节解码轨迹与实测轨迹形状相似性Pearson相关系数分别提高了0.81、0.19和0.63。
明朝继承元朝的东北疆域后,于明成祖永乐七年(1409)置奴儿干都指挥使司(简称奴儿干都司),统辖包括今库页岛、北海道在内的黑龙江流域。
在采用BiLSTM神经网络模型,从EEG信号中连续解码左腿VM肌肉通道的sEMG波形的研究中,虽然可解码出部分波形,但平均的形状相关性较低,解码波形的Pearson相关系数
接近0.5,说明EEG信号中存在和sEMG相似的运动模式。综上,研究证明从EEG信号中可解码出肌肉通道的sEMG信号波形,但还需进行深入研究相关的解码算法和增强EEG中运动信息,以改善解码精度。
:
[1] YOUNG A J,FERRIS D P.State of the art and future directions for lower limb robotic exoskeletons [J].IEEE Transactions on Neural Systems and Rehabilitation Engineering,2017,25(2):171-182.
[2] KAPSALYAMOV A,JAMWAL P K,HUSSAIN S,et al.State of the art lower limb robotic exoskeletons for elderly assistance [J].IEEE Access,2019,7:95075-95086.
[3] TARIQ M,TRIVAILO P M,SIMIC M.EEG-based BCI control schemes for lower-limb assistive-robots [J].Frontiers in Human Neuroscience,2018,12:312.
[4] 陈朝峰.穿戴式下肢外骨骼系统设计与交互控制策略研究 [D].哈尔滨:哈尔滨工业大学,2020.
[5] GORDLEEVA S Y,LOBOV S A,GRIGOREV N A,et al.Real-time EEG-EMG human-machine interface-based control system for a lower-limb exoskeleton [J].IEEE Access,2020,8:84070-84081.
[6] 杨灿军,彭桢哲,徐铃辉,等.柔性膝关节保护外骨骼及其行走助力方法设计 [J].浙江大学学报(工学版),2021,55(2):213-221.
YANG Canjun,PENG Zhenzhe,XU Linghui,et al.Design of flexible knee-joint protection exoskeleton and walking assistance method [J].Journal of Zhejiang University (Engineering Science),2021,55(2):213-221.
[7] WINTER C,KERN F,GALL D,et al.Immersive virtual reality during gait rehabilitation increases walking speed and motivation:a usability evaluation with healthy participants and patients with multiple sclerosis and stroke [J].Journal of NeuroEngineering and Rehabilitation,2021,18(1):68.
[8] 陈超强,蒋磊,王恒.基于SAE和LSTM的下肢外骨骼步态预测方法 [J].计算机工程与应用,2019,55(12):110-116,154.
CHEN Chaoqiang,JIANG Lei,WANG Heng.Gait prediction method of lower extremity exoskeleton based on SAE and LSTM neural network [J].Computer Engineering and Applications,2019,55(12):110-116,154.

[10] 郝静涵.基于SEMG信号及加速度信号的步态辨识研究 [D].天津:河北工业大学,2019.
[11] 郭浩.基于EMG的人体下肢运动意图感知与预测研究 [D].哈尔滨:哈尔滨工业大学,2019.
[12] 李梦瑶.基于多源神经信号融合的人体运动意图识别研究 [D].北京:中国科学院大学,2021.
[13] 刘笃信.下肢外骨骼机器人多模融合控制策略研究 [D].深圳:中国科学院大学(中国科学院深圳先进技术研究院),2018.
[14] NAKAGOME S,LUU T P,HE Yongtian,et al.An empirical comparison of neural networks and machine learning algorithms for EEG gait decoding [J].Scientific Reports,2020,10(1):4372.
[15] FITZSIMMONS N A,LEBEDEV M A,PEIKON I D,et al.Extracting kinematic parameters for monkey bipedal walking from cortical neuronal ensemble activity [J].Frontiers in Integrative Neuroscience,2009,3:3.
[16] YI Chunzhi,JIANG Feng,BHUIYAN M Z A,et al.Smart healthcare-oriented online prediction of lower-limb kinematics and kinetics based on data-driven neural signal decoding [J].Future Generation Computer Systems,2021,114:96-105.
[17] JIA LU,AI Qingsong,MENG Wei,et al.Individualized gait trajectory prediction based on fusion LSTM networks for robotic rehabilitation training [C]∥2021 IEEE/ASME International Conference on Advanced Intelligent Mechatronics (AIM).Piscataway,NJ,USA:IEEE,2021:988-993.
[18] DELORME A,MAKEIG S.EEGLAB:an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis [J].Journal of Neuroscience Methods,2004,134(1):9-21.
[19] CHOI J,KIM H.Real-time decoding of EEG gait intention for controlling a lower-limb exoskeleton system [C]∥2019 7th International Winter Conference on Brain-Computer Interface (BCI).Piscataway,NJ,USA:IEEE,2019:1-3.
[20] GWIN J T,GRAMANN K,MAKEIG S,et al.Electrocortical activity is coupled to gait cycle phase during treadmill walking [J].NeuroImage,2011,54(2):1289-1296.
[21] WEI Pengna,ZHANG Jinhua,WANG Baozeng,et al.Surface electromyography and electroencephalogram-based gait phase recognition and correlations between cortical and locomotor muscle in the seven gait phases [J].Frontiers in Neuroscience,2021,15:607905.
[22] PRESACCO A,GOODMAN R,FORRESTER L,et al.Neural decoding of treadmill walking from noninvasive electroencephalographic signals [J].Journal of Neurophysiology,2011,106(4):1875-1887.
[23] SAXENA M,GUPTA A.Exploration of temporal and spectral features of EEG signals in motor imagery tasks [C]∥2021 International Conference on Communication Systems &Networks (COMSNETS).Piscataway,NJ,USA:IEEE,2021:736-740.
[24] SORIANO-SEGURA P,IANEZ E,ORTIZ M,et al.Detection of the intention of direction changes during gait through EEG signals [J].International Journal of Neural Systems,2021,31(11):2150015.
[25] WEI Pengna,ZHANG Jinhua,TIAN Feifei,et al.A comparison of neural networks algorithms for EEG and sEMG features based gait phases recognition [J].Biomedical Signal Processing and Control,2021,68:102587.
猜你喜欢
现代仪器与医疗(2022年4期)2022-10-08
包装工程(2022年10期)2022-05-27
中学生数理化(高中版.高考数学)(2022年4期)2022-05-25
读友·少年文学(清雅版)(2020年4期)2020-08-24
读友·少年文学(清雅版)(2020年3期)2020-07-24
南方周末(2019-12-19)2019-12-19
中国外汇(2019年19期)2019-11-26
南方周末(2019-07-18)2019-07-18
南方周末(2019-05-09)2019-05-09
科学之谜(2018年4期)2018-09-17
