
大口黑鲈抗缪勒氏管激素(AMH)原核表达及多克隆抗体制备
2022-09-19 08:08王向斌梁子怡王心雨刘慧芬
河南水产 2022年4期
郭 琦, 王向斌, 黄 茜, 梁子怡, 王心雨, 刘慧芬*
(河南师范大学水产学院, 河南新乡 453007)
抗缪勒氏管激素(Anti-Müllerian hormone,Amh)又称缪勒氏管抑制物, 是转化生长因子-β(Transforming growth factor-β,TGF-β) 超家族中的一员[1]。在哺乳动物中,Amh表达于精巢的支持细胞,可引起缪勒氏管的退化,维持雄性特征[2]。 雌性个体中,Amh在原始卵泡的募集以及优势卵泡选择中发挥重要作用[3]。 虽然绝大多数鱼类没有缪勒氏管,但是却保留了Amh基因。 2002年在日本鳗鲡中发现Amh基因,主要在支持细胞中表达,与精子发生有关[4]。 硬骨鱼类中,虽然Amh主要在雄性性别分化中发挥关键作用,但其在雌性卵巢中也有表达。 在斑马鱼中敲除Amh基因会导致卵巢肥大,初级卵母细胞数量显著增多,卵母细胞发育进程受阻[5]。 使用花鳍骨AMH重组蛋白对卵母细胞进行体外孵育, 结果发现AMH重组蛋白可以促进花鳍骨卵母细胞成熟[6]。这说明Amh在鱼类性腺发育过程中发挥一定作用。
大口黑鲈(Micropterus salmoides),又名加州鲈,隶属于鲈形目 (Perciformes)、 太阳鱼科(Cehtrachidae)、黑鲈属(Micropterus),具有肉质鲜美、抗病力强、 生长迅速、 适温范围较广等优点。 大口黑鲈自1983年被引入内地以来, 在全国多地区实现了人工养殖[7]。目前关于大口黑鲈品种开发、营养品质、养殖模式分析等方面已有较多研究, 但是有关其性腺发育及相关分子调控机制的研究鲜有报道。
本研究利用已经获得的大口黑鲈Amh基因cDNA全长序列,构建原核表达载体,制备多克隆抗体,以期为研究Amh基因的功能及其在大口黑鲈性腺发育中的分子机制提供参考。
1 材料与方法
1.1 试验材料与主要试剂
大口黑鲈购自于新乡市洪门镇鲈鱼养殖基地,暂养于河南师范大学水产养殖基地。 用于多克隆抗体制备的小鼠购自华兰生物工程股份有限公司。
感受态细胞、限制性内切酶、pET-32a(+)载体等购自宝日医生物技术(北京)有限公司。 彩虹180广谱蛋白Marker、 弗氏佐剂等购自北京索莱宝科技有限公司。His Tag HP镍柱购自上海生工生物工程股份有限公司。 实验所用引物均由生工生物工程(上海)股份有限公司合成。
1.2 原核表达载体构建
根据Amh基因序列, 选择合适的区域设计特异性引物, 同时根据原核表达载体pET32a的图谱选取酶切位点,具体信息见表1:

表1 重组载体引物
以大口黑鲈卵巢cDNA为模板,通过PCR扩增序列、 琼脂糖凝胶电泳回收PCR产物。 将回收产物和pET-32a质粒用BamH I 和Xho I限制性内切酶双酶切(37℃,过夜),酶切产物纯化后,构建重组表达载体pET32a-AMH。 将重组质粒转化至BL21感受态细胞中,挑选阳性克隆送至生工生物工程(上海)股份有限公司测序, 采用DNAMAN软件对测序结果进行比对分析,以判断目的基因是否正确插入载体。
1.3 融合蛋白诱导表达
将阳性表达菌接入LB液体培养基中进行扩大培养(取部分菌液作为对照),37℃震荡培养至OD6000.5左右, 向其中加入IPTG (Isopropyl β-D- Thiogalactopyranoside,终浓度1 mmol/L)进行诱导表达,37℃震荡培养,在培养6 h、8 h、10 h及12 h后取材,12000 rpm离心20 min,收集沉淀。菌体沉淀以4×loading buffer重悬裂解,随后使用SDS-PAGE对总菌体、沉淀和上清进行检测。
1.4 融合蛋白纯化及抗体制备
用上述条件大量诱导蛋白, 在室温下8000 rpm离心5 min收集菌体, 加入适量变性结合缓冲液,超声波破碎 (破3 s, 停3 s, 功率50%, 破碎总时间30 min),获得融合蛋白,用His Tag HP柱纯化蛋白。蛋白纯化所需试剂配制及操作方法参照冯军厂的方法[8]。 用纯化透析后的融合蛋白免疫小鼠,每隔一周加强免疫一次, 首次将融合蛋白与弗氏完全佐剂等比混合后对小鼠进行皮下注射, 之后每次将融合蛋白与弗氏不完全佐剂等比混合后注射,4周后采血获得抗血清。
1.5 抗体效价检测
利用酶联免疫 (enzyme linked immunosorbent assay,ELISA)法检测制备抗体的效价,测定时以未免疫的小鼠血清作为阴性对照。
1.6 数据分析
采用SPSS 22.0统计分析, 不同组之间的差异显著性(P<0.05)采用单因素方差分析(ANOVA)进行检验,Duncan氏方法多重比较, 实验数据均使用平均值±标准误(mean ± SE)表示。
2 结果与分析
2.1 重组表达载体构建
以大口黑鲈卵巢cDNA为模板进行扩增,获得的目的片段与预期大小一致 (图1 a)。 重组质粒pET32a-AMH经单双酶切验证, 检测目的片段已成功导入表达载体(图1 b)。 随后,将产物进行纯化、测序,结果表明构建的pET32a-AMH正确。
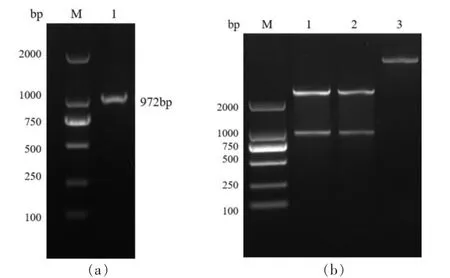
图1 Amh目的片段扩增及验证
2.2 重组蛋白诱导表达及可溶性检测
构建好的重组质粒pET32a-AMH转入大肠杆菌BL21。 随后采用IPTG在16℃下进行诱导,SDS-PAGE检测发现,在52.3 kDa左右出现目的条带,而对照在同一区域未出现特异条带, 表明重组表达载体pET32a-AMH构建成功。 诱导后菌体经超声破碎、离心、SDSPAGE电泳分析表明重组蛋白主要存在于沉淀中。
2.3 重组蛋白纯化
对诱导菌液进行收集、破碎,利用亲和纯化的方法,对AMH融合蛋白进行纯化。使用不同浓度咪唑洗脱液洗脱蛋白,SDS-PAGE 电泳检测结果显示,100 mM咪唑洗脱时开始检测到目的蛋白;150 mM和200 mM咪唑洗脱时,检测到大量的目的蛋白,但是200 mM 咪唑浓度洗脱时,获得的目的蛋白纯度更高(图2)。经透析去除杂质,SDS-PAGE电泳检测结果显示,纯化后的杂蛋白明显减少,条带较为单一,可用于后续实验(图3)。
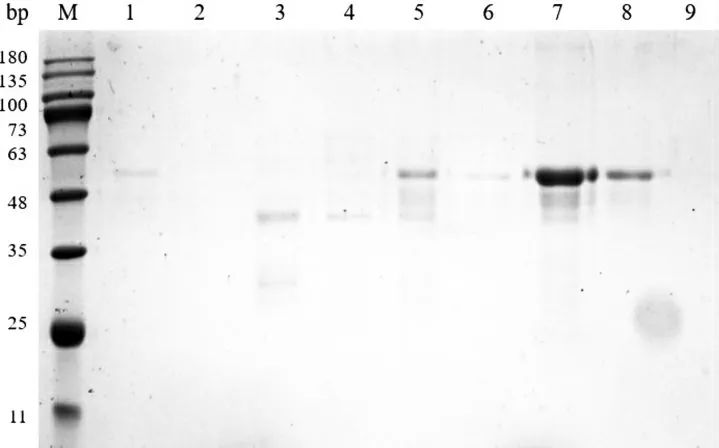
图3 AMH重组蛋白不同浓度咪唑洗脱结果

图4 AMH重组蛋白纯化
2.4 AMH多克隆抗体制备及效价检测
经过4次免疫,获得鼠源AMH抗血清。 使用酶联免疫吸附方法对获得的多克隆抗体血清进行效价测定,结果显示多克隆抗体的效价为2.7×105,可为后续实验提供原材料。
3 讨论
抗缪勒氏管激素是转化生长因子家族成员之一,在哺乳动物中具有调节细胞发育及分化,诱导缪勒氏管退化,维持雄性特征的作用。当前已有较多研究表明,Amh在哺乳动物卵巢发育过程中发挥一定作用。 在硬骨鱼类中,Amh多被当作雄性性别分化关键基因。 尼罗罗非鱼XX个体中,amhy的过表达导致了雌性到雄性的性逆转[9]。 也有研究表明,Amh在雌性个体卵巢发育过程中发挥一定作用。 在黑鲷研究中发现,Amh在卵巢发育早期不表达,在卵黄蛋白原合成期表达量升高[10]。 斑马鱼中的研究发现,Amh缺失会导致雌鱼性腺发育不良,卵泡发生受阻[11]。 这说明,Amh基因在鱼类性腺发育过程中发挥重要作用。
原核表达是外源基因融合表达的方式之一, 具有易培养、生长快、操作简单等优点[12]。 多克隆抗体具有稳定性较好、制备技术要求不高、周期短、成本低等优点,广泛用于蛋白亚细胞定位、蛋白互作、细胞信号通路等研究领域[13]。 本研究利用获得的大口黑鲈Amh基因序列,成功构建pET32a-AMH原核表达载体,诱导出大小为52.3kDa的AMH重组蛋白, 与奥利亚罗非鱼49kDa的AMH重组蛋白[14]及双须骨舌鱼48kDa的AMH重组蛋白[15]大小相近。随后通过免疫小鼠,获得AMH多克隆抗体,利用ELISA对抗体血清进行效价检测,结果显示多克隆抗体的效价为2.7×105,可用于后续研究。
本研究通过构建大口黑鲈AMH原核表达载体,获得AMH重组蛋白,利用重组蛋白对小鼠进行免疫,获得AMH多克隆抗体。 获得的大口黑鲈AMH重组蛋白和多克隆抗体可为后续研究Amh在大口黑鲈中的功能和作用机制提供基础。
猜你喜欢
环球时报(2022-09-20)2022-09-20
宝藏(2022年1期)2022-08-01
当代水产(2022年6期)2022-06-29
心理学报(2022年2期)2022-02-17
延安大学学报(医学科学版)(2021年3期)2021-10-25
现代检验医学杂志(2021年4期)2021-08-10
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
时代英语·高二(2015年2期)2015-05-18
青年文摘·上半月(1984年1期)1984-11-01
