
谷子NPR1基因特征及响应禾生指梗霉侵染的表达分析
2022-09-19 12:12刘旭张诺徐林孙卓楠张宝俊
山西农业科学 2022年9期
刘旭,张诺,徐林,孙卓楠,张宝俊
(山西农业大学 植物保护学院,山西 太谷 030801)
植物与病原菌在长期进化的过程中,形成了高效的免疫防御系统。由病原菌保守相关分子模式触发的免疫反应(PAMPs-Triggered Immunity,PTI)和效应子触发的免疫反应(Effector-Triggered Immunity,ETI)在寄主抵御外来病原菌侵染中发挥重要作用。由水杨酸(Salicylic acid,SA)的产生及积累诱发的植物系统获得性抗性(Systemic acquire resistance,SAR)也是寄主受病菌侵染后激活防卫反应的重要组成部分,并伴随病程相关蛋白(Pathogenesis-related proteins,PRs)的表达[1]。病程相关基因1的非表达子(NPR1)是水杨酸信号通路中核心调控元件,是植物病程相关蛋白基因表达及SAR的激活子[2]。当病菌侵入寄主植物后,NPR1寡聚体在水杨酸的积累下,被氧化还原为单体,并与转录因子TGA互作,从而诱导PR基因的表达[3]。
NPR1基因主 要包含2个 结 构域:BTB/POZ和Ankyrin-Repeat结构域,且与蛋白间的互作紧密相关[4-5]。研究人员首先在拟南芥中鉴定到6个NPR1基因[6-9],CAO等[7]研 究 发 现,AtNPR1通过启动 子区W-box序列与转录因子WRKY相结合,激活下游PR基因的表达,诱导寄主获得系统抗病性,此外,AtNPR5、AtNPR6还被证实参与调控叶片及花器官的生长发育[8-9]。随着生物信息学及测序技术的发展,NPR1基因相继在多种作物中被鉴定,主要在植物抗病性方面发挥重要作用。SILVA等[10]在二倍体草莓中过表达拟南芥AtNPR1,发现AtNPR1通过诱导防御基因FvPR1和FvPR5转录本的积累,增强草莓对白粉病、角斑病及炭疽病的抗性;YUAN等[11]在水稻中过表达OsNPR1,显著增强水稻对白叶枯病的抗性以及草食性生物对水稻的敏感性;WANG等[12]在本氏烟和油菜中过表达BnaNPR1,激活了与SA防御反应相关基因的表达,而抑制了与JA信号相关基因的表达,增强其对核盘菌的抗性。可见,NPR1基因在植物响应病菌侵染的过程中,通过激活相关抗病基因及SAR信号途径来抵御病原菌侵染。
谷子白发病是由禾生指梗霉(Sclerospora graminicola)引起的一种系统性侵染卵菌病害,严重影响着谷子的产量。禾生指梗霉是一种典型的活体营养型病菌,研究禾生指梗霉侵染过程中SA的响应及积累对谷子抗白发病机制的研究具有重要意义,而有关谷子SiNPR1家族基因的研究鲜有报道。
为了挖掘谷子中NPR1基因的相关功能,本研究通过生物信息学方法筛选及鉴定谷子全基因组中的NPR1家族基因,对其进行基因定位、蛋白特性分析、进化分析、motif分析、启动子顺式作用元件分析,并分析了谷子在响应禾生指梗霉侵染条件下NPR1基因的表达模式,深入解析NPR1家族基因在谷子生长发育及抗病代谢通路中发挥的作用,旨在为谷子抗病育种奠定基础。
1 材料和方法
1.1 谷子NPR1家族基因的获取及基因鉴定
从Multi-omics Database forSetaria italica数据库(http://foxtail-millet.biocloud.net/home)中获取xiaomi基因组序列,以从国家水稻数据中心(https://www.ricedata.cn/)下载 到 的5条水稻NPR1蛋白序列及TAIR数据库(https://www.arabidopsis.org/)获取的6条拟南芥NPR1蛋白序列作为搜索谷子同源基因的探针序列。在TBtools工具中利用BLAST GUI Wrapper程序,在谷子全基因组蛋白序列中搜索同源序列。选取比对值Evalue<0.01。
1.2 基因定位及蛋白特性分析
利用ExPASY-ProtParam在线工具(https://web.expasy.org/protparam/)预测谷子NPR1家族蛋白的理化性质,包括基因组位置、氨基酸数目、分子量、等电点、亲水性均值;利用PSORT Prediction在线工具(http://psort1.hgc.jp/form.html)进行亚细胞定位预测分析。
1.3 谷子NPR1家族基因系统发育树构建
使用MEGA 7.0对筛选的拟南芥、水稻和谷子的NPR1家族蛋白序列进行序列比对,采用邻接法(Neighbor Joining Algorithm)构建系统发育树,其中BootStrap参数设置为1 000,其他参数选择默认。借助在线工具iTOL(https://itol.embl.de/)进一步优化进化树。
1.4 谷子NPR1家族基因motif、基因结构、顺式作用元件预测分析
从谷子基因组gff3文件中筛选出NPR1基因结构信息,利用MEME网站(http://meme-suite.org/tools/meme)对谷子NPR1蛋白进行保守结构域预测,并在TBtools中可视化。在Pfam(http://pfam.xfam.org/)数据库进行保守结构域分析。将谷子NPR1家族基因的启动子序列在Plantcare(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)网站进行在线顺式作用元件分析,预测该家族基因可能存在的功能。
1.5 谷子NPR1基因表达差异分析
从Multi-omics Database forSetaria italica数据库(http://foxtail-millet.biocloud.net/home)中获得谷子种子、根、茎、穗、叶不同组织的基因表达量数据。基于各基因在植物组织各器官的表达差异构建表达热图,并利用可视化软件TBtools展示。
依据实验室前期转录组数据,以晋谷20为植物材料,将上一年从病残体中收集的卵孢子与种子按1∶15的比例进行拌种,种植于山西农业大学杂粮试验基地,以甲霜灵拌种作为对照(甲霜灵∶种子=1∶500)。在营养生长期、生长锥伸长期、枝梗分化期、小穗和刚毛分化期、雌雄蕊分化期(时期记为T1、T2、T3、T4、T5)取叶片及幼穗,对照健康叶片组记为Leaf_CK1-Leaf_CK5,处理组记为Leaf_TG1-Leaf_TG5;对照健康幼穗组记为Panicle_CK1-Panicle_CK5,处理组记为Panicle_TG1-Panicle_TG5,分析谷子NPR1基因在禾生指梗霉侵染下不同时期的表达情况,利用RStudio对各组FPKM值进行pheatmap图绘制。
2 结果与分析
2.1 谷子NPR1基因筛选、基因定位和蛋白特性分析
利用BLASTP在xiaomi基因组中鉴定到4个NPR1基 因,命 名 为SiNPR1~SiNPR4,均 包 含BTB/POZ和Ank结构域(图1-A)。利用TBtools对谷子NPR1基因进行染色体定位,SiNPR1~SiNPR4基因分布在5、8、9号染色体上(图1-B)。
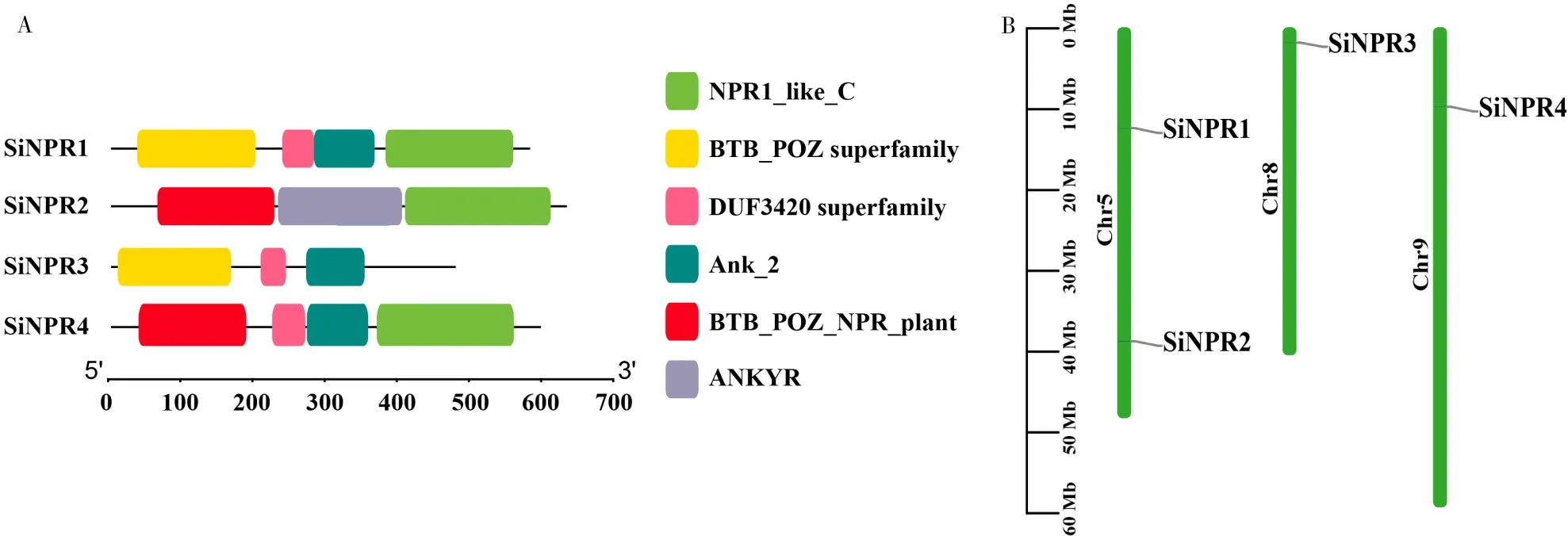
图1 谷子NPR1基因结构域及基因定位Fig.1 NPR1 gene domain and gene location in foxtail millet
理化性质分析表明,谷子NPR1家族基因氨基酸数目分布在475~629个,平均数目为569个;蛋白分子质量介于50.29~68.96 ku;等电点位于5.52~6.11;除SiNPR3表现为疏水性外,其他均表现为亲水蛋白;亚细胞定位预测显示,SiNPR1定位于细胞核中,SiNPR3定位于质膜,SiNPR2和SiNPR4均定位于叶绿体中(表1)。
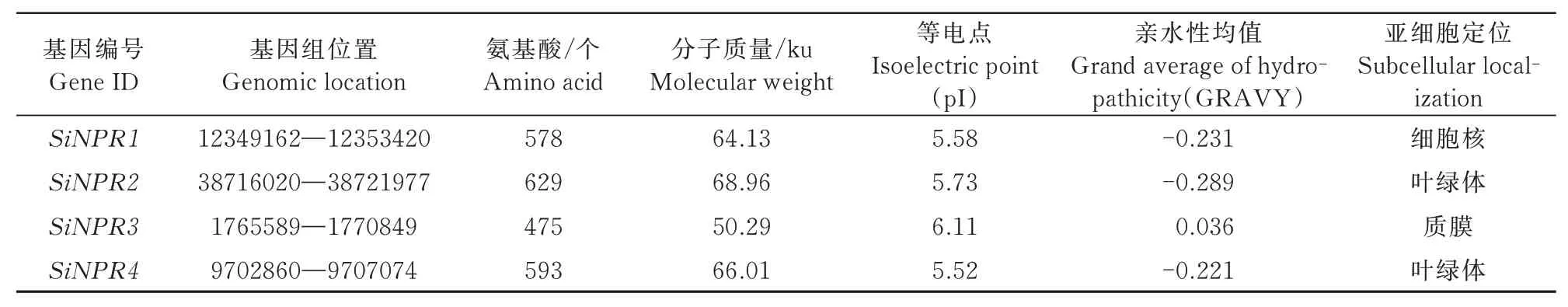
表1 谷子NPR1基因理化性质分析Tab.1 Analysis of physicochemical properties of NPR1 genes in foxtail millet
2.2 谷子NPR1家族成员系统进化分析
谷子NPR1家族基因系统进化分析结果如图2所示。

图2 谷子NPR1家族基因系统进化分析Fig.2 Phylogenetic analysis of NPR1 family genes in foxtail millet
谷子NPR1家族基因系统进化树共分为3类(图2),其中Class1中SiNPR2与OsNPR2的亲缘关系最近,SiNPR4与OsNPR3的亲缘关系最近,但均与AtNPR3和AtNPR4同源;Class2表明,SiNPR1与OsNPR1的同源关系最近,且与AtNPR1和AtNPR2同源;Class3中SiNPR3与OsNPR5、AtNPR5、AtNPR6、OsNPR4同源,其 中AtNPR5和AtNPR6的同源关系最近。
2.3 谷子NPR1家族基因保守结构域及基因结构分析
对谷子NPR1家族基因进行保守结构域和基因结构分析发现,谷子NPR1家族基因中含有8个基序(motif1~motif8),长度分布在24~50个氨基酸(表2),其中SiNPR1、SiNPR2、SiNPR4均包含8个motif基序,且基序排列顺序均为motif6、motif2、motif7、motif3、motif4、motif5、motif1、motif8。SiNPR3仅含有5个保守基序,不包含motif1、motif5、motif8(图3-A)。基因结构分析表明,所有家族成员在上下游均含有调控区(UTR)、外显子,但其具体的位置和长度存在差异(图3-B)。

表2 谷子NPR1家族基因蛋白基序信息Tab.2 Protein sequence information of NPR1 family genes in foxtail millet
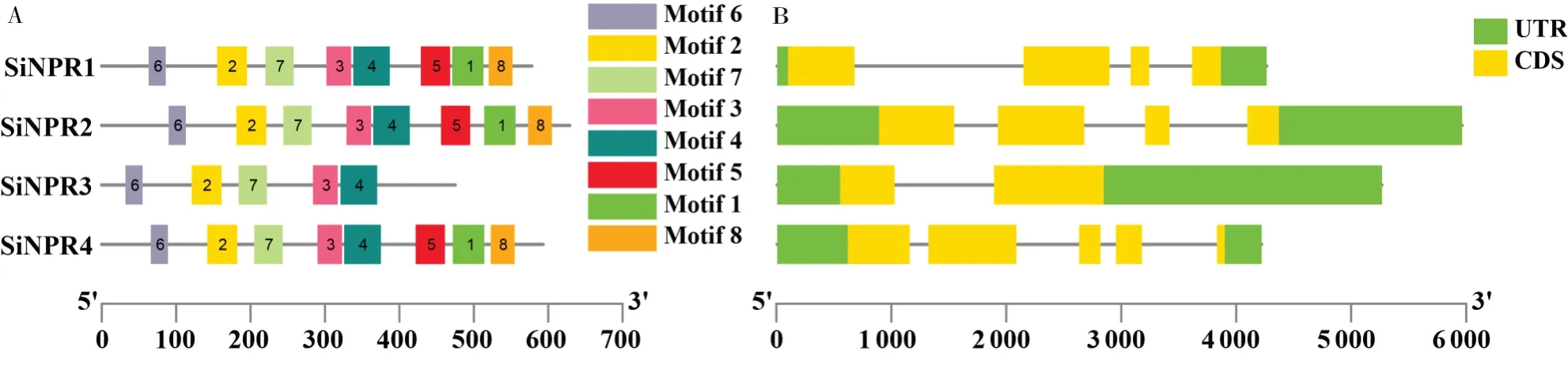
图3 谷子NPR1家族基因保守基序分析Fig.3 Conservative motif analysis of NPR1 family genes in foxtail millet
2.4 谷子NPR1家族基因顺式作用元件
顺式作用元件分析结果显示,SiNPR1~SiNPR4基因包含的元件分别有16、27、18、41个(图4),均包含脱落酸反应元件(abscisic acid responsive element)、防御和压力反应元件(defense and stress responsive element)和光响应元件(light responsive element)。其 中,仅SiNPR1和SiNPR4含 有 水 杨酸反应元件(salicylic acid responsive element),推测其可能通过调控水杨酸生物合成来调控谷子对病菌侵染的响应;SiNPR3和SiNPR4均含有赤霉素反应元件(gibberellin responsive element)、生长素响应元件(auxin responsive element)、茉莉酸甲酯响应元件(MeJA responsive element)及低温响应元件(low temperature responsive element),推测其可能参与调控谷子的生长发育。
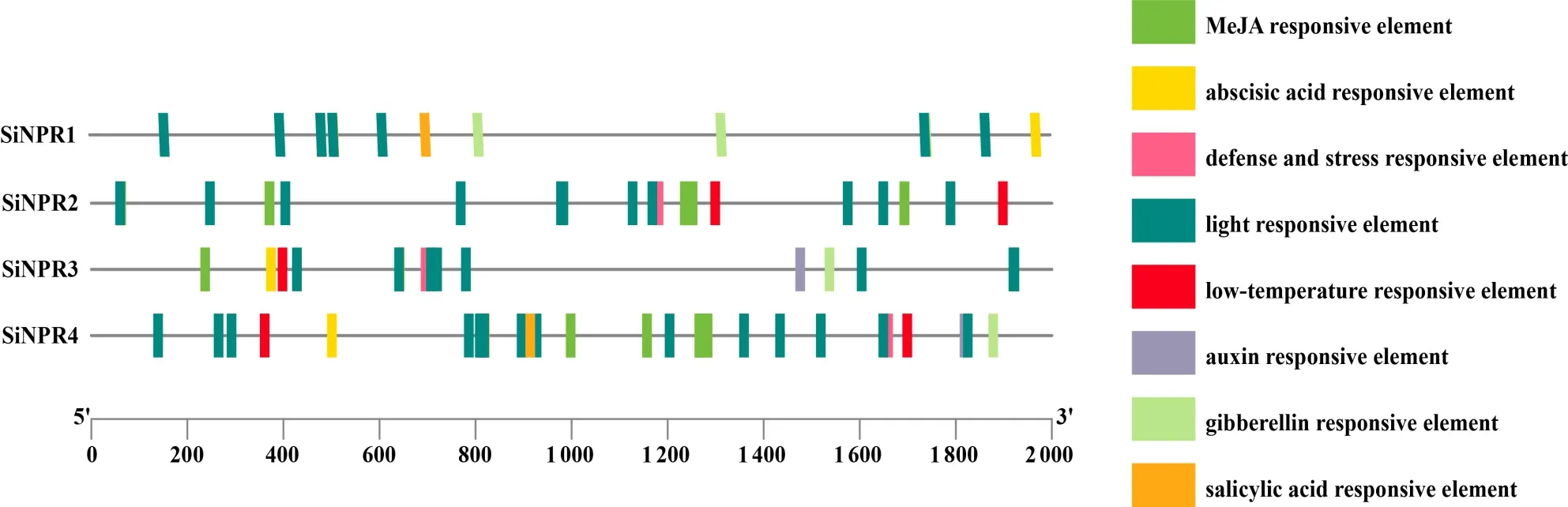
图4 谷子NPR1家族基因顺式作用元件分析Fig.4 Analysis of cis-acting elements of NPR1 family genes in foxtail millet
2.5 谷子NPR1家族基因表达差异分析
谷子NPR1家族基因组织特异性表达结果表明,NPR1家族基因在谷子不同部位的FPKM值分布在0~82.2,且在不同组织中均存在表达特异性(图5)。
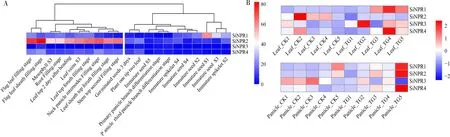
图5 NPR1家族基因表达热图Fig.5 Gene expression heat map of NPR1 family genes
图5-A从左到右依次为灌浆期旗叶(Flag leaf filling stage);旗叶鞘充盈期(Flag leaf sheath filling stage);叶肉S3时期(Mesophyll S3);根填充阶段(Root filling stage);抽穗后2 d的旗叶(Leaf top 2 days after heading);叶脉S3时期(Leaf veins S3);灌浆期旗叶(Leaf top filling stage);颈穗节间充盈期(Neck panicle internodes filling stage);灌浆期叶鞘(Leaf sheath top filling stage);茎顶第2填充阶段(Stem top second filling stage);3 d发 芽 的 种 子(Germinated seeds 3 d);一尖两叶期(Plant one tip two leaf);未成熟种子S5时期(Immature seed S5);初生穗分枝分化阶段(Primary panicle branch differentiation stage);穗部第3穗分枝分化期(Panicle third panicle branch differentiation stage);未成熟小穗S4时期(Immature spikelet S4);未成熟种子S4时期(Immature seed S4);未成熟种子S2时 期(Immature seed S2);未 成 熟 种 子S1时 期(Immature seed S1);未成熟种子S3时期(Immature seed S3);未成熟小穗S2时期(Immature spikelet S2)。由图5可知,SiNPR2在旗叶的不同时期及其他叶肉细胞中特异高表达,其中旗叶鞘充盈期FPKM值高达82.2,其次是灌浆期旗叶和颈穗节间充盈期,而在种子和谷穗中低表达或不表达;SiNPR1在未成熟种子S1时期表达量最高,为59.58,而在穗部各时期均不表达;SiNPR3和SiNPR4均在灌浆期旗叶中高表达,而在未成熟种子的各时期低表达或不表达。表明NPR1家族基因在不同组织中存在不同的表达特性,预示其在谷子的生长发育及抗逆反应中的复杂功能。对NPR1家族基因在不同时期组织进行聚类分析,发现不同时期种子聚为一类,不同时期叶片聚为一类,表明NPR1家族基因在特定组织的不同时期的表达具有稳定性。
转录组分析结果表明,在禾生指梗霉胁迫下,SiNPR1、SiNPR3、SiNPR4在叶片中呈上调趋势,SiNPR2表达量下调(图5-B),其中,SiNPR1在T3、T4、T5时期明显上调,以T4时期差异最为显著,该时期是禾生指梗霉病菌有性态卵孢子形成关键期,会导致谷子叶肉细胞破坏,叶片发生纵裂。SiNPR1、SiNPR2、SiNPR4在穗部转录组中也表现出上调表达趋势,其中,SiNPR1在T5时期差异最为显著,此时是病菌侵染穗部的关键时期。
3 结论与讨论
SA是植物体内的小分子酚类物质,参与活体或半活体营养型病原菌引发的ETI免疫反应并诱发植物的系统获得抗性。NPR1是水杨酸信号通路中一类关键因子,作为SA的受体能与SA分子结合,激活TGAs等转录因子的活性,启动PR等抗病基因的表达,从而诱导产生SAR。NPR1赋予小麦、苹果、烟草等作物抗性的研究相继报道[13-15]。研究表明,在芥菜、柑橘中过表达NPR1基因可提高寄主对病原菌的抗性[16-17]。此外,NPR1家族基因在植物非生物胁迫方面也发挥作用,如AtNPR1与HSFA1相互作用,提高拟南芥的抗寒性[18]。
本研究在谷子中共鉴定到4个NPR1家族基因,理化性质分析表明,谷子NPR1家族基因氨基酸数目分布在475~629个,蛋白分子质量介于50.29~68.96 ku,等电点位于5.52~6.11,与香蕉、油桐中报道的较为相似[19-20]。谷子NPR1家族基因分布在5、8、9号染色体上,且均包含BTB/POZ和Ankyrin结构域,为与TGA蛋白互作提供了条件[21]。motif分析中共鉴定到8个motif基序,除SiNPR3以外,其他家族基因基序排列顺序一致,推测其可能与遗传进化中的变异有关;基因结构分析发现,所有家族成员在上下游均含有调控区(UTR)、内含子、外显子。谷子NPR1家族基因在各个组织部位均有表达,但不同组织间的表达存在差异,与大白菜NPR1家族基因的组织表达模式相似[22]。
YUAN等[11]研究发现,过表达OsNPR1被证实可提高水稻对白叶枯病的抗性;JIANG等[23]研究发现,OsNPR3与TGAL11互作能提高水稻对白叶枯病的抗性;HEPWORTH等[9]研究表明,AtNPR5、AtNPR6参与调控拟南芥叶片和花器官的发育。进化分析表明,SiNPR1、SiNPR4分别与OsNPR1、OsNPR3同源,SiNPR2与OsNPR2同源,结合转录组结果分析发现,受侵染叶片的SiNPR1在T3、T4、T5时期的表达显著上调,分别是对照的6.57倍、10.25倍、9.91倍,此外,T4、T5时期SiNPR3、SiNPR4表达上调,而SiNPR2表达下调;谷穗中SiNPR1、SiNPR2、SiNPR4在T5时期表达量显著上调,SiNPR3显著下调,表明SiNPR1、SiNPR4可能在谷子抗禾生指梗霉侵染过程中起正调控作用;SiNPR2在叶中特异高表达,推测其可能参与谷子的生长发育与形态建成。尽管SiNPR3与AtNPR5、AtNPR6同源,但在各组织中均不表达,其功能有待进一步确定。
本研究通过BLASTP从xiaomi数据库中共鉴定到4个谷子NPR1基因,命名为SiNPR1~SiNPR4,生物信息学分析表明,SiNPR1定位于细胞核内,SiNPR4定位于叶绿体中,均含有SA响应元件。经进化分析及转录组分析可知,SiNPR1与OsNPR1的同源性最高,在禾生指梗霉侵染后T5时期的叶、穗均显著上调,推测SiNPR1参与激活谷子SA介导的SAR表现。此外,SiNPR2、SiNPR4与AtNPR3、AtNPR4同源,且均定位于叶绿体,其中,SiNPR2在禾生指梗霉侵染谷子后的叶T1~T5时期均下调表达,推测在谷子抗禾生指梗霉反应中起负调控作用;SiNPR4在禾生指梗霉侵染后T5时期的叶、穗均显著上调,推测其可能抵御卵孢子形成。本研究结果可为进一步探究谷子的生长发育及抗病机制提供一定的参考依据。
猜你喜欢
传感器世界(2022年6期)2022-11-25
当代水产(2022年1期)2022-04-26
当代陕西(2021年21期)2022-01-19
中国瓜菜(2019年8期)2019-09-19
科技与创新(2019年16期)2019-08-26
延河(2018年6期)2018-06-11
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2017年1期)2017-02-27
中学生数理化·中考版(2016年7期)2016-12-07
职业·下旬(2009年8期)2009-10-12
