
半红树植物苦郎树光响应模型拟合及光合特性研究
2022-09-19 06:49白艳斌刘佳哲余小红樊东函
湖北林业科技 2022年4期
白艳斌 刘佳哲 余小红 樊东函
(1.广西壮族自治区国有高峰林场 南宁 530002; 2.钦州市林业科学研究所 钦州 535099)
半红树植物是能在潮间带和陆地非盐渍土生长的两栖木本植物,具有独特的生境适应性和生态价值,也是丰富物种多样性和湿地植被恢复的重要对象[1]。苦郎树Volkameriainermis为唇形科Lamiaceae苦郎树属Volkameria的攀援状灌木,在中国福建、台湾以及北部湾地区的海岸潮滩均有分布,是一种沿海防沙造林的树种[2]。在红树林群落中苦郎树常以伴生树种的形式生长,或是生在红树林群里的林缘边[3],不仅在水土保持、红树林生态系统上有着重要价值,还具有一定的园林绿化功能[4]。此外,苦郎树提取的化合物在医药和农业有多种用途,对治疗皮肤病、风湿疼痛等疾病有一定作用,试验发现苦郎树提取物对多种植物病原真菌有抗菌活性[5]。面对沿海生态依然存在的突出问题,提高沿海地区植被覆盖率、建设沿海防护林对防风减灾有重要意义,提升红树林生态稳定性和多样性是海防林研究的重点。目前国内外对苦郎树的研究较少,仅限于扦插或组培等育苗技术研究、解剖学研究和提取物抑菌活性的研究[5-8],对光合特性的研究还未有报道。
光合作用是植物通过一系列复杂的生理进程将光能转化为化学能合成有机物的过程,为植物生长提供了基础,因此探究植物光合特性是研究植物最基础且不可或缺的一部分[9]。可通过光响应模型拟合光响应曲线,同时计算出光饱和点、光补偿点、最大净光合速率、暗呼吸速率和表观量子效率等光合参数[10]。由于不同模型拟合结果存在差异,因此通过研究常用的直角双曲线模型、非直角双曲线模型、指数模型和直角双曲线修正模型的拟合结果,选择最适合的拟合模型对苦郎树光响应曲线的拟合尤为重要。
本研究对广西北部湾地区苦郎树的光响应曲线测定,使用直角双曲线模型、非直角双曲线模型、指数模型和直角双曲线修正模型共4种光响应拟合模型拟合,评估4种光响应模型的拟合精度后找出最适模型,并研究苦郎树的光合特性,以期为日后利用苦郎树恢复红树林生态、提升物种多样性、防风固堤以及园林绿化提供参考依据。
1 材料与方法
1.1 样地及试验材料
研究样地位于广西钦州市康熙岭镇的广西茅尾海红树林自治区级自然保护区内(21°52′N,108°30′E)。该地区属于亚热带季风气候,夏季高温多雨,冬季温暖。年均气温22.2 ℃,其中最高月均温28.1 ℃,最低月均温 12.8 ℃。日照时长1 800 h,平均年降雨量2 150 mm,年平均相对湿度82 %,无霜期354 d。样地生境内海水盐度为25.6 ‰,土壤pH值8.3。
在广西茅尾海红树林自治区级自然保护区中潮滩带的苦郎树自然生长群落内选取3个5 m×5 m样地,样地之间间隔大于20 m,每个样地内选择3株成熟苦郎树植株,每株植株选取自上而下第二片健康、成熟且向阳的叶片进行光响应曲线测量。
1.2 光响应曲线测定方法
在晴朗无云的上午9∶00~11∶30时间段内,采用LI-6400XT便携式光合仪(LICOR,Lincoln,Nebraska,USA)的LED红蓝光源测定光响应曲线。使用开放式气路,在测定前使用1 200 μmol·m-2·s-1光强诱导20 min后选择LightCurve2预设程序测定,光强设置1 800、1 500、1 200、800、500、200、150、100、75、50、25、10、0 μmol·m-2·s-1共13个梯度。设置每次改变光强后最小等待时间120 s,最大等待时间200 s,直至所有光强梯度均测量结束后更换下一片叶子继续测量。
1.3 光响应模型
采用4种光响应模型对苦郎树光响应曲线进行拟合,并计算其光合参数。
1.3.1 直角双曲线模型
直角双曲线模型(RHM)[11]表达式为:
式中,I为光合有效辐射(μmol·m-2·s-1),Pn为净光合速率(μmol·m-2·s-1),Pmax为最大净光合速率(μmol·m-2·s-1),α为初始表观量子效应,Rd为暗呼吸速率(μmol·m-2·s-1)。
1.3.2 非直角双曲线模型
非直角双曲线模型(NHM)[12]表达式为:
式中,θ为反映非直角双曲线弯曲程度的曲角参数,取值范围为0≤θ≤1,其他参数意义同上。
1.3.3 指数模型
指数模型(EM)[13]表达式为:
式中,e为自然对数的底,其他参数意义同上。
1.3.4 直角双曲线修正模型
直角双曲线修正模型(MRHM)[14]表达式为:
式中,β和γ为系数,其他参数意义同上。
以上四种模型中通过模型计算得出相应系数后,令Pn=0可解出各模型拟合后光补偿点LCP的值。直角双曲线修正模型的光饱和点LSP可通过光合计算模型直接计算得出,直角双曲线和非直角双曲线需要通过解I≤200μmol·m-2·s-1时拟合直线方程Pmax=AQE×I-Rd得出,式中AQE为表观量子效率,即拟合直线方程的斜率。指数模型通常是假设Pn=0.9Pmax或0.99Pmax时的有效光合辐射为LSP的值。
1.4 模型评价
使用均方根误差(RMSE)[10]和决定系数(R2)对4个模型拟合结果的精确度进行评价,RMSE越小的同时R2越大则说明拟合优度越好。R2通过光合计算模型计算,RMSE公式如下:
式中,yo,i为净光合速率实测值,ym,i为净光合速率模型拟合值,n为观测个数。
1.5 水分利用效率的估算
水分利用效率(WUE)表达式为:WUE=Pn/Tr,式中Pn为净光合速率(μmol·m-2·s-1),Tr为蒸腾速率μmol·m-2·s-1。
1.6 数据处理与分析
在Excel2016中进行光合模型拟合和RMSE模型评价,单因素方差分析和多重比较使用SPSS 26处理和分析,使用R(v4.1.0)和Excel 2016分别绘制箱形图和折线图。
2 结果与分析
2.1 不同光响应模型拟合结果
4种光响应模型对苦郎树光响应曲线拟合的决定系数均接近0.999水平,指数函数模型相较其他三种模型决定系数较低(表1,图1)。实测光响应曲线与不同模型拟合后的光响应曲线如图3-A所示。
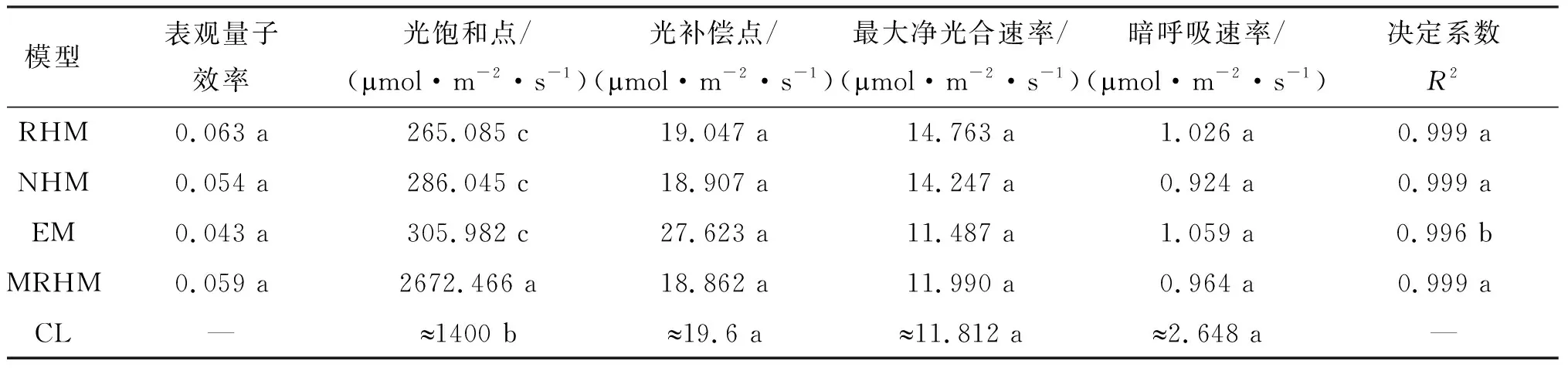
表1 4种光响应模型对苦郎树光合参数的拟合
不同模型的表观量子效应AQE无显著差异(P>0.05),其中直角双曲线模型AQE最大,指数模型AQE最小;在LSP拟合中4个光合拟合模型与实测值存在显著差异(P<0.05),直角双曲线修正模型拟合的LSP异常过大。直角双曲线模型、非直角双曲线模型和指数模型的LSP数值大小较为接近但与实测值依然存在显著差异(P<0.05)(图1-A);LCP拟合中,4个模型拟合值与实测值无显著差异(P>0.05),其中指数模型LCP最大,直角双曲线模型LCP最小,指数函数模型LCP大于实测值(图1-B);Pmax拟合中4个模型拟合值与实测值无显著差异(P>0.05),其中直角双曲线模型Pmax最大,指数模型Pmax最小,除指数模型以外三种模型均大于实测值(图1-C);Rd拟合中4个模型拟合值与实测值无显著差异(P>0.05),其中实测值Rd均大于4个拟合模型的拟合值,最小为非直角双曲线模型拟合值(图1-D)。

图1 4种光响应模型对苦郎树的光合参数拟合值与实测值对比
2.2 光响应模型的拟合优度
本研究采用RMSE和R2进一步评估4种不同光响应模型对苦郎树光响应曲线拟合的优度和精度。RMSE越低且R2越高则表示该模型的拟合精度较高,反之较差。通过计算RMSE和R2并进行方差分析,结果表明指数模型与直角双曲线模型RMSE与其它两种模型存在显著差异(P<0.05);指数模型R2与其他三种模型存在显著差异(P<0.05)。由图2可知,4种拟合模型的精度由高到低依次为:非直角双曲线模型、直角双曲线修正模型、直角双曲线模型和指数模型。非直角双曲线模型和直角双曲线模型的拟合精度都较高且较为接近,但直角双曲线修正模型LSP拟合值异常偏大,4种模型LSP拟合值均与实际值有显著差异,因此选用非直角双曲线模型对苦郎树光响应曲线拟合效果相对,LSP的拟合还需进一步的研究。
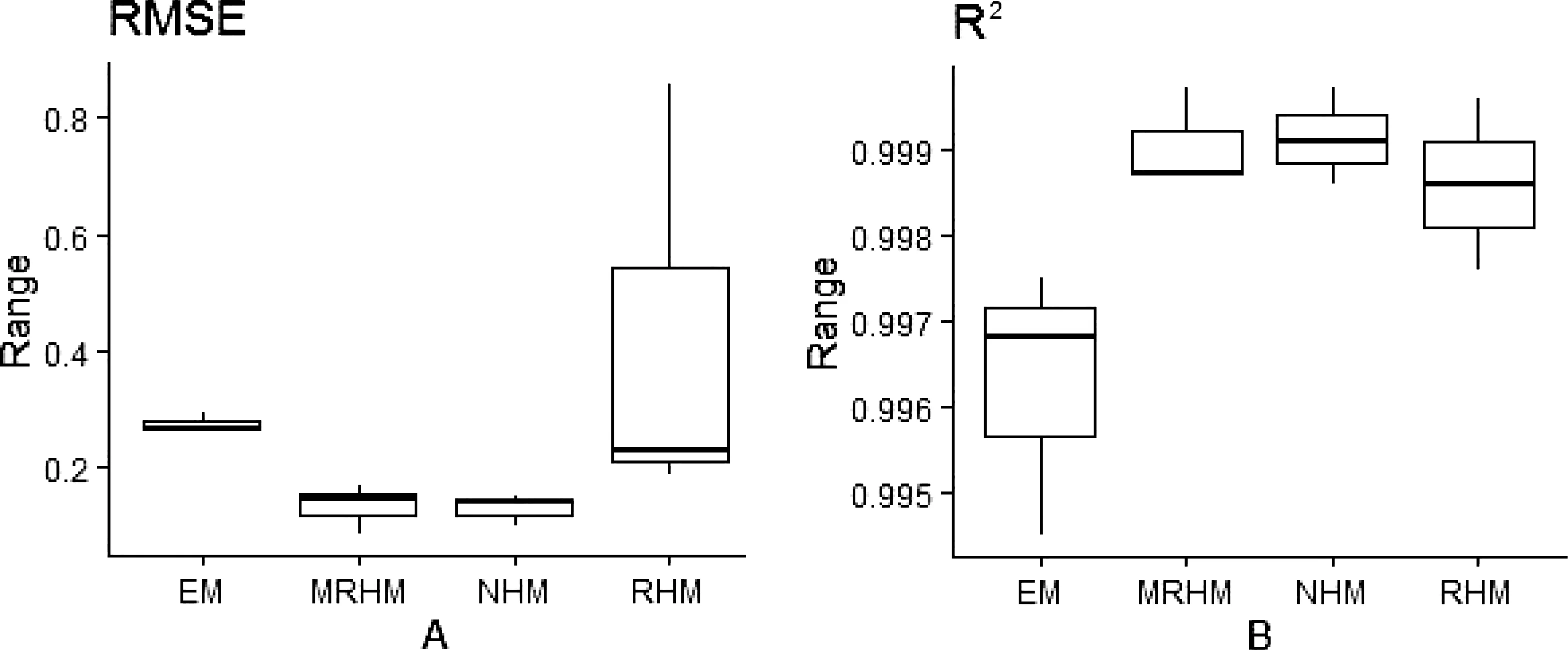
图2 4种光响应模型对苦郎树的拟合优度比较
2.3 气孔导度、胞间CO2浓度和水分利用效率光响应曲线
气孔是植物光合作用中CO2吸收和水分蒸腾的通道,气孔导度(Gs)描述植物气孔张开度,一般情况下随着光合速率的提升Gs也随之增大[15]。苦郎树Gs光响应曲线如图3-B所示,随着有效光合辐射的增加Gs也随之上升,但随着光饱和点的到来Gs持续上升并未随Pn下降而减小。
胞间CO2浓度(Ci)与植物的光合速率密切相关,光合作用消耗CO2和H2O为植物提供能量释放氧气。苦郎树Ci浓度光响应曲线如图3-C所示,随着有效光合辐射的增加Ci浓度持续下降,在光合辐射0~500 μmol·m-2·s-1区间内Ci浓度下降趋势较快,当光合辐射大于500 μmol·m-2·s-1后Ci浓度呈平缓趋势,当植物光饱和后Ci浓度略有回升。
水分利用效率(WUE)是植物蒸腾速率与净光合速率的比值,水也是光合作用的重要底物之一,WUE反映植物对水分的利用效率。苦郎树WUE光响应曲线如图3-D所示,WUE呈先升高再降低的趋势,当光合辐射小于500 μmol·m-2·s-1时WUE快速上升,达到最高值后在光合辐射大于500 μmol·m-2·s-1时缓慢下降。
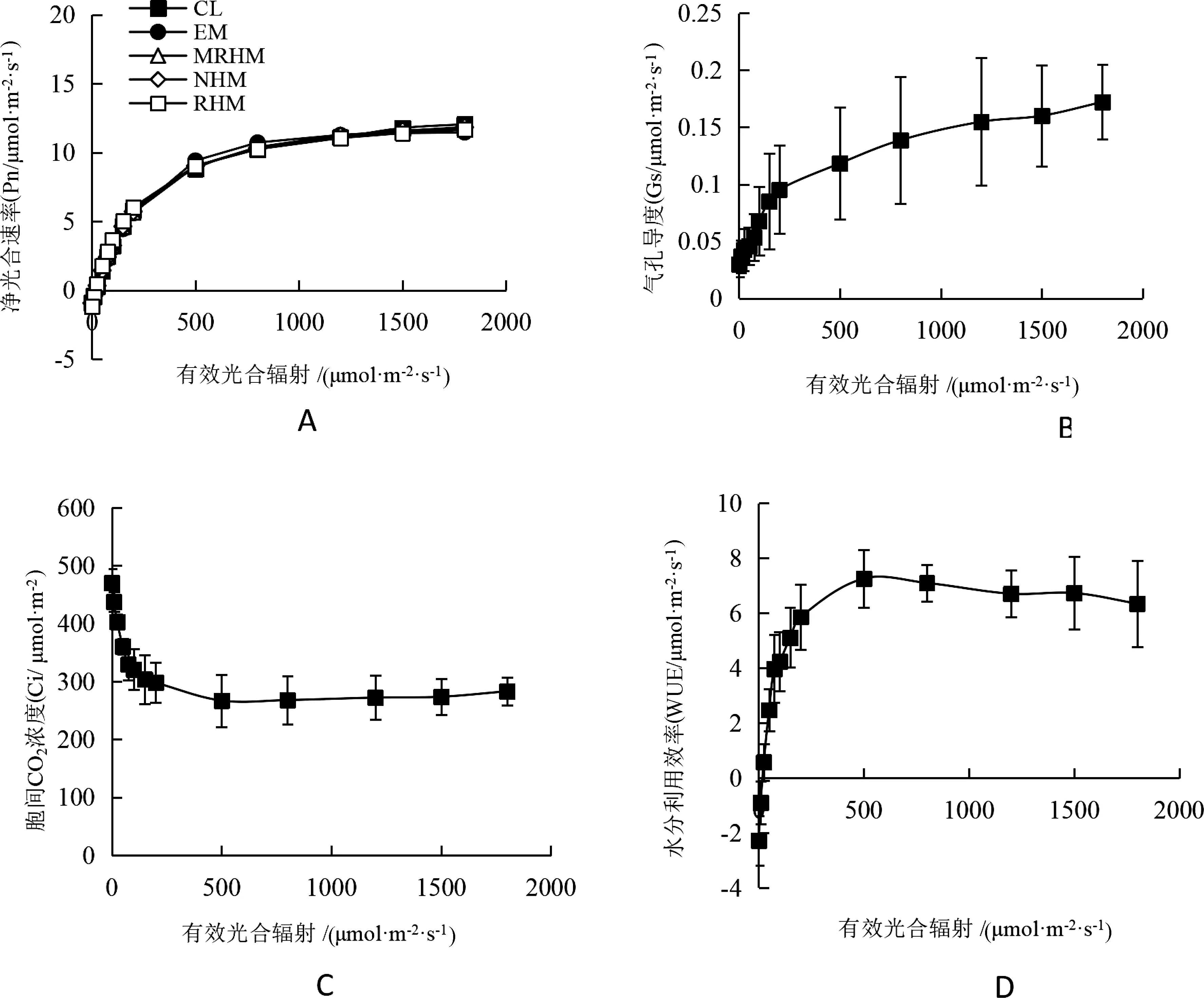
图3 苦郎树净光合速率、气孔导度、胞间CO2浓度和水分利用效率光响应曲线注:误差线为标准误差。
3 结论与讨论
3.1 不同模型对苦郎树光响应曲线的拟合效果
本研究使用4种光响应模型对苦郎树光响应参数拟合,并进行对比分析,结果表明不同模型对苦郎树光响应拟合存在差异,非直角双曲线模型对苦郎树拟合效果最好。非直角双曲线模型能够比较准确地拟合出苦郎树在不同光合辐射下的净光合速率的变化,同时在LCP、Rd和Pmax等光合参数的拟合上与实测值较为接近,但出现LSP偏小的现象。近年来国内学者使用的最多的光拟合模型是直角双曲线修正模型[16],因为该模型既能准确的计算出LSP,也能模拟出光抑制下的LCP,如王海珍等[17]对胡杨、刘阳阳等[18]对山葡萄以及刘强等[19]对长白落叶松等研究验证了直角双曲线修正模型的有效性。但在本研究种直角双曲线修正模型对LSP的拟合结果异常偏高,与实际情况明显不符,是由于植物的特性不同,不同植物适用的光拟合模型也不用,为了得到准确的光合参数可能需要使用不同模型进行拟合比较。如郎莹等[20]对山杏的光响应模拟结果发现在非直角双曲线模型适用于正常情况下,不能用于受到胁迫下的山杏光响应曲线模拟,因此应需要根据实际情况选用最适宜的光响应拟合模型。
通过比较R2水平可以发现4种模型中除指数模型外均达到0.999水平,RMSE结果表明直角双曲线最高,非直角双曲线最低。使用R2和RMSE评估拟合模型的适用性是通过数学统计的方式对模型的评估,有可能出现过拟合的现象,即决定系数较高但实际拟合后的光合参数与实际值相差甚远[21]。本研究中,4种模型的LSP拟合值存在显著差异,LCP、Rd和Pmax拟合值无显著差异。指数模型、非直角双曲线模型和直角双曲线模型LSP明显低于实际值,而非直角双曲线修正模型明显偏高,造成这一结果可能是强光光强设置梯度太大,LSP没有被测量到导致拟合结果的异常。因此评估模型的适用性不仅需要依据R2和RMSE判断,也要根据实际的拟合效果综合分析。综上,使用非直角双曲线模型可对苦郎树光响应曲线拟合,并计算LCP、Rd和Pmax等光合参数,但苦郎树LSP的精确计算方式还需进一步研究。
3.2 苦郎树的光合特性
植物光响应曲线能够直接的反映植物对不同光合辐射下的光合特性,本研究通过4种模型拟合了苦郎树光合-光响应曲线,同时计算气孔导度、胞间CO2浓度和水分利用效率光响应曲线。结果表明苦郎树在光抑制阶段(0~500 μmol·m-2·s-1)中净光合速率随光合辐射的增加快速上升,高光强下净光合速率增长缓慢,甚至下降。这一结果与大部分植物的光合特性相同。气孔导度-光响应曲线和胞间二氧化碳-光响应曲线结果表明苦郎树气孔导度随光合辐射增加而增加,相反胞间CO2浓度随光合辐射的升高而降低。气孔导度与胞间CO2均反映的是植物光合作用消耗气体的过程,通常植物在到达光饱和点后气体的交换速率变缓,同时净光合速率下降,如申文辉等[15]对格木、李佳等[22]对米槁的光合特性研究结果与本研究结果基本一致。苦郎树水分利用效率-光响应曲线呈先升高再降低的趋势,在光合辐射小于500 μmol·m-2·s-1时水分利用效率呈升高趋势,光合辐射大于500 μmol·m-2·s-1时呈下降趋势。造成水分利用效率这一趋势的原因很大程度上与植物光饱和点有关,当植物光饱和点达到,过高光强抑制净光合速率,净光合速率的降低影响对水分的利用效率。汪凤林等[23]对杉木和邓云鹏等[24]对栓皮栎的水分利用效率研究结果表明,在弱光强下杉木和栓皮栎水分利用效率升高迅速,在高光强下有下降的趋势,与本研究结果基本一致。
综上所述,在4种不同光响应拟合模型对苦郎树光合参数拟合对比中发现,4种模型拟合决定系数均在0.996以上,其中非直角双曲线模型拟合效果最佳,但4种不同模型对LSP拟合均存在一定差异,如何精确拟合LSP还需进一步研究,应根据实际情况选用最适宜的光响应拟合模型。苦郎树气孔导度随光合辐射增加而增加;胞间二氧化碳浓度随光合辐射的升高而降低;苦郎树水分利用效率呈先升高再降低的趋势,与大部分植物的光合特性相似。因此,在海堤植被恢复、红树林生态保护、湿地公园建设、园林绿化中种植苦郎树应充分考虑其光合特性,构建合理的群落结构,最大程度满足苦郎树对光照的需求,以发挥其最大的生态和景观价值。
猜你喜欢
灌溉排水学报(2022年9期)2022-09-28
探测与控制学报(2022年4期)2022-08-30
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
宁夏大学学报(自然科学版)(2022年2期)2022-07-18
农业工程学报(2022年4期)2022-04-24
农业与技术(2020年12期)2020-07-04
中国房地产业·下旬(2020年12期)2020-01-11
新课程·下旬(2017年7期)2017-08-14
大陆桥视野·下(2016年11期)2017-02-28
