
水稻抽穗期调控基因Hd6的PARMS标记开发与利用
2022-09-19 04:28:40彭永彬杜晨阳郑崇珂周晋军孙伟和亚男谢先芝
山东农业科学 2022年8期
彭永彬,杜晨阳,郑崇珂,周晋军,孙伟,和亚男,谢先芝
(1.山东省农业科学院湿地农业与生态研究所,山东 济南 250100;2.山东农业大学生命科学学院,山东 泰安 271018)
抽穗期反映了水稻品种对种植区域气候和生态环境的适应性,是水稻重要的农艺性状之一[1]。抽穗过早会导致水稻因对光、热资源利用不足而减产,而抽穗过晚易遭遇低温等灾害天气,同样会导致减产[2]。水稻抽穗期受多种外界因素如光周期、温度、土壤肥力等的影响,其中光周期是核心因素。水稻属于短日照作物,短日照条件下促进开花,长日照条件下抑制开花[3]。基于水稻对光周期的敏感性,其生长阶段可分为基本营养阶段(BVP)和光周期敏感阶段[4],其中,BVP的长短和对光周期的敏感性是由遗传因素决定的,不同栽培品种由于具有不同的抽穗期基因型组合而表现出不尽相同的抽穗期,因此,研究水稻抽穗期及其相关基因,鉴定可适应不同地区的最佳抽穗期基因组合对培育地区适应性较强的品种具有重要意义。挖掘水稻抽穗期遗传位点,通过分子育种将抽穗期位点导入性状优良但熟期不适的品种中,或是培育优良水稻品种的关键。
水稻的抽穗期受多基因组成的复杂数量性状基因座(QTLs)所调控。利用正、反向遗传学和群体遗传学,大量与抽穗期有关的基因被克隆,其中,有多个涉及水稻抽穗期感光性的主效基因,如Hd1、Hd6、Ghd7及DTH8等[5-11]。克隆的抽穗基因多数作用于成花基因的上游,可大致归为两种相互串联的调控通路:OsGI-Hd1-Hd3a通路和以Ehd1为核心的调控通路。Hd1作为栽培稻驯化过程中受选择的基因之一,其功能具有双重性:在长日照条件下,Hd1促进Ghd7表达,并与之形成抑制复合体,抑制Ehd1-Hd3a开花通路,导致不同程度的延迟抽穗甚至不抽穗;而在短日照条件下,Ghd7表达下调,Hd1与抑制复合体形成竞争关系,促进Hd3a的表达,从而促进水稻抽穗[2,12]。Hd6是一个主效光周期响应位点基因[8],编码酪蛋白激酶CK2的α亚基,可通过磷酸化DTH7蛋白间接调控Hd1的表达,从而在长日照条件下抑制抽穗;此外,Hd6可磷酸化昼夜节律调节因子OsLHY[13],其不同自然变异类型能精调水稻抽穗期的昼夜临界时长[14]。但Hd6延迟抽穗的作用仅仅是在其有功能的前提下才能发挥,‘日本晴’和‘Kasalath’的Hd6等位基因(Hd6Nip和Hd6Kasa)在编码区存在一个核苷酸的替换,致使Hd6Nip编码提前终止,而Hd6Kasa正常行使调控抽穗的功能[8,13]。Hd6基因的功能多样性有助于水稻栽培面积进一步向北扩大,在水稻育种中具有重要的应用价值。分子标记是指以核苷酸序列变异为基础的遗传标记,能够直接反映个体基因组DNA的差异。经典的分子标记主要有RFLP(restriction fragment length polymorphism,限制性片段长度多态性)、SSR(simple sequence repeat,简单重复序列)、RAPD(random amplified polymorphic DNA,随机扩增多态性DNA)、SNP(single nucleotide polymorphisms,单核苷酸多态性)以 及InDel(insert and deletion,插 入 缺失)[15-17]等。利用分子标记育种技术,可实现对水稻产量、品质以及抗性等方面的改良[18]。PARMS技术是基于扩增受阻突变的体系PCR(ARMS-PCR)开发的一种SNP基因分型技术[19],增加了两条不同荧光标记引物,可通过荧光信号的差异检测基因的多态性。与传统的需要电泳分型的标记相比,PARMS标记具有操作简便、耗时短、成本低等优点,而且可基于CRISPR/Cas9基因编辑过程中产生的单碱基插入缺失和小片段缺失对基因编辑后代进行基因分型鉴定。
本研究根据水稻抽穗期基因Hd6Kasa的SNP位点开发了一个PARMS分子标记Hd6Kasa-PARMS,并利用该标记对以‘蜀恢498’为供体亲本、‘苏秀867’为轮回亲本构建的BC2F2群体进行基因型鉴定,可以准确高效地鉴定出Hd6Kasa基因型,为培育具有优良生育期和适应性的栽培水稻品种提供了有力工具。
1 材料与方法
1.1 供试材料
‘蜀恢498’是具有高配合力优质的水稻恢复系材料,作为重穗型骨干亲本,基因资源丰富,熟期适当,适宜于我国籼稻区种植。‘日本晴’是典型的粳稻材料,常作为基础研究的对照和受体。‘苏秀867’是黄淮稻区粳型常规水稻,在产量品质及抗病性等方面表现比较均衡,适合种植于山东南部、沿淮及淮北地区。‘9311’是籼稻代表性品种,具有较为完善的基因组数据库,常作为籼稻基础研究对照。供试水稻材料均种植于济南市历城区饮马泉实验站,常规大田水肥管理。
1.2 水稻叶片DNA的提取
取幼嫩的水稻叶片于2 mL离心管中,参考凌飞等[20]的CTAB法提取叶片DNA。
1.3 引物设计与PCR反应
根据目的SNP位点开发了一个分子标记Hd6Kasa-PARMS,根据目的SNP位点上下游共129 bp的DNA序列设计了3条标记引物,包括两条正向特异性引物Hd6Kasa-PARMS-FAM(5′-GAAGGTGACCAAGTTCATGCTTGGAGGTCCAAA CATTGTGT-3′)和Hd6Kasa-PARMS-HEX(5′-GAAGGTCGGAGTCAACGGATTTGGAGGTCCAAA CATTGTGA-3′)及一条反向通用引物Hd6Kasa-PARMS-common(5′-TCATAATCTGTCAACGTGGGGTAC-3′),其中FAM的标签序列为GACCAAGTTCATGCT,HEX的标签序列为CGGAGTCAACGGATT。引物由擎科生物科技有限公司合成。
PARMS-PCR扩增体系(10μL):94℃预变性15 min;94℃变性20 s,65℃(每循环下降0.8℃)退火及延伸1 min,扩增10个循环;94℃变性20 s,57℃退火及延伸1 min,扩增28~30个循环。产物用QuantStudio 5荧光定量PCR仪的Genotyping模式采集FAM和HEX荧光信号值并自动进行基因分型。
常规PCR扩增体系(20μL):94℃预变性5 min;94℃变性10 s,55℃退火30 s,72℃延伸1 min,扩增30个循环;72℃延伸2 min,12℃保存。产物用1%琼脂糖凝胶电泳检测。切胶回收,纯化后送至擎科生物科技公司测序。引物为Hd6-F(5′-ATGGGGTGAGCAGGATGACT-3′)和Hd6-R(5′-CGTGGGGTACAGCACTTTGA-3′)。
2 结果与分析
2.1 PARMS标记的开发与应用结果
已知‘Kasalath’与‘日本晴(Nipponbare)’的Hd6等位基因存在一个碱基替换(A436-T436),而正是这个碱基的改变,导致两个基因型的功能发生质的改变[8,13]。通过测序比对发现,‘蜀恢498’与‘Kasalath’Hd6基因序列的碱基序列一致,SNP位置均为A;而‘苏秀867’与‘日本晴’的一致,均为T(图1A)。利用‘蜀恢498’为供体亲本、‘苏秀867’为轮回亲本构建BC2F2群体,为了筛选出该BC2F2群体中携带抽穗期基因Hd6Kasa的品系,在该SNP位点处设计了一个PARMS分子标记Hd6Kasa-PARMS(图1B)。从BC2F2群体中选取50个单株,利用Hd6Kasa-PARMS标记进行基因分型,结果(图1C)显示,利用该标记可很好地区分开Hd6基因分型,相同基因分型的聚为一类,其中,17个单株基因型与供体亲本相同,即图中的蓝点样本;22个为杂合子,即图中的绿点样本;11个与轮回亲本基因型相同,即图中的红点样本。上述结果表明筛选出的39个携带QTL位点Hd6Kasa的单株可作为育种中间材料,继续扩大种植和筛选。

图1 分子标记Hd6Kasa-PARMS的开发与利用
2.2 群体基因型的测序验证
为了进一步验证PARMS标记的特异性,对50份BC2F2样品进行PCR扩增和Sanger测序。序列比对结果(表1)发现,有17份样品SNP位点处碱基为A且为单峰,与‘Kasalath’的基因型一致;有11份样品SNP位点处碱基为T且为单峰,与‘日本晴’的基因型一致;22份样品SNP位点处碱基显示为A/T且双峰,为杂合基因型。这与利用PARMS标记得到的基因分型结果一致(图1、表2)。说明Hd6Kasa-PARMS标记能非常精准地从群体中筛选出携带抽穗期基因Hd6Kasa的个体。

表1 BC2F2群体SNP位点的测序分型结果
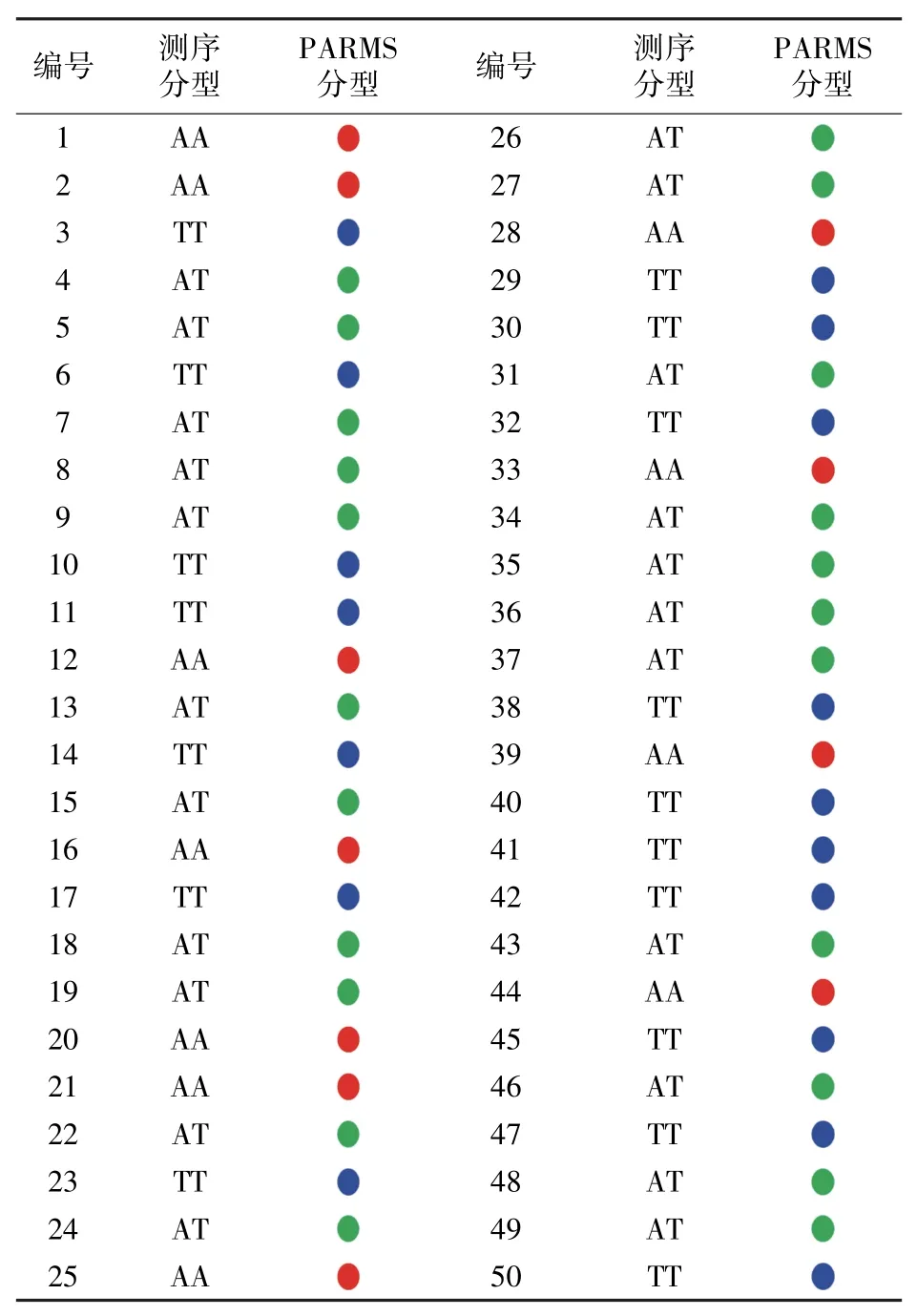
表2 Hd6Kasa-PARMS与DNA测序基因分型结果对应
2.3 群体基因型和抽穗期表型的相关性分析
为了阐释Hd6基因的功能和开发标记的必要性,对BC2F2群体各单株的抽穗期进行统计,结果(表1)显示,‘Kasalath’基因型的17个单株平均抽穗时长为155 d左右,杂合型的22个单株平均抽穗时长为143 d左右,‘日本晴’基因型的11个单株平均抽穗期为145 d左右。利用SPSS 26.0软件对抽穗期表型与Hd6基因分型的量化数据进行处理,将Hd6Kasa基因型设置为数字0,Hd6Nip基因型以及杂合型设置为数字1,结合50份单株的抽穗期调查,对基因型与表型进行相关性分析,结果显示,两者相关系数为0.858(P<0.01),达到极显著水平,说明水稻抽穗期表型与Hd6基因型密切相关。长日照条件下,Hd6Kasa基因型的导入能有效改变水稻的抽穗期。
3 讨论与结论
选育具有理想抽穗期的水稻品种是实现特定产区增产的关键。在常规育种工作中,育种家们会尽量选择生育期合适的株系;而在杂交育种过程中,各组合抽穗期的测定耗时耗力,尤其由籼粳交组合中选育的品种抽穗期普遍较晚,无法准确选育出抽穗期适合的品种,很大程度上限制了水稻亚种间杂种优势的利用。在过去的几十年里,大量的抽穗期基因及其等位基因型被挖掘分析,为选育理想抽穗期的品种提供了有力条件。当前分子标记辅助选择、基因工程调控以及基因编辑技术均可有效利用抽穗期相关的基因资源,培育熟期合适的栽培品种。其中,分子标记辅助育种相对于传统的育种方法,具有高效便捷的优势,且不受外界环境条件的干扰,是目前应用最广泛的育种方法之一[18]。Hd6基因编码区存在一个碱基替换,引起的自然变异导致水稻抽穗期调控功能上的籼粳分化,同时Hd6能精调水稻抽穗期的昼夜临界时长,其功能多样性使其具有重要的育种应用价值。
与早期传统的分子标记相比,PARMS标记更加高效、准确、快速及高通量,只需进行PCR扩增即可完成检测,且试验成本与传统方法差别不大。本研究即基于高通量的PARMS标记检测技术,以Hd6Kasa和Hd6Nip之间的SNP位点开发了Hd6Kasa-PARMS标记,利用该标记可以准确区分出Hd6基因分型,能从以‘蜀恢498’为供体亲本、以‘苏秀867’为轮回亲本获得的BC2F2群体中筛选出携带抽穗期基因Hd6Kasa的单株,经Sanger测序验证,准确率高达100%。综上所述,利用Hd6Kasa-PARMS标记可快速准确筛选出携带Hd6Kasa基因的株系,这有利于充分发挥抽穗期调控基因Hd6在水稻育种中的应用价值,提高育种效率,为高效培育抽穗期理想的水稻品种提供了方法。
猜你喜欢
安徽农学通报(2022年6期)2022-04-07 21:30:29
教学考试(高考生物)(2020年6期)2020-11-23 05:25:56
基层中医药(2020年5期)2020-09-11 06:32:00
食品与生物技术学报(2020年8期)2020-01-06 08:00:56
科学24小时(2019年5期)2019-06-11 08:39:38
发明与创新(2019年9期)2019-03-26 02:22:48
基层中医药(2018年5期)2018-08-31 02:35:42
畜牧与饲料科学(2018年3期)2018-05-08 02:48:35
湖南林业科技(2017年1期)2017-02-06 05:28:55
制造技术与机床(2015年10期)2015-04-09 07:06:14
