
牧草混播生态系统功能研究进展
2022-09-16 03:43:26杨策张玉雪张鹤郑春燕朱峰
草业学报 2022年9期
杨策,张玉雪,张鹤,郑春燕,朱峰*
(1.中国科学院遗传与发育生物学研究所农业资源研究中心,河北省土壤生态学重点实验室,中国科学院农业水资源重点实验室,河北 石家庄 050022;2.中国科学院大学,北京 100049;3.河北农业大学园艺学院,河北 保定 071000)
近年来,随着社会经济的快速发展,居民对营养健康的要求不断提升,我国牛羊肉及奶制品的需求量不断增加,畜牧业所需饲草料缺口巨大[1]。尽管我国拥有大面积的天然草地,但由于气候、土壤等自然因素和过度放牧、管理粗放等人为因素影响,90%的天然草地发生了不同程度的退化,其生产力低下,生态功能也未得到有效发挥[2-3]。另外,目前我国的人工草地多是单一化种植,结构简单,严重依赖自然因素,产量稳定性差,牧草品质较低。因此,建设高产优质的人工草地,缓解由于天然草地过度利用所造成的生态威胁,是我国经济发展与农业结构大调整的一项重要任务[4]。
众所周知,生物多样性是群落和生态系统过程和功能的重要决定因素。大量生态学研究表明,物种多样性增加对于生态系统生产力、稳定性、抵抗生物入侵以及养分循环均有积极的影响[5-7]。与单播草地相比,混播草地不仅在提高饲草产量[8]、改善品质[9]方面具有优势,还在改善土壤肥力、调控土壤微生物群落结构[10-11]、增加生态系统稳定性[12],实现系统可持续生产方面具有明显作用。因此,本研究针对混播草地种间互作机理及其生态系统功能和服务的研究现状,重点对混播草地增产提质、提高稳定性和土壤健康等生态系统功能和服务等方面进行分析和总结,并对今后的研究方向和生产实践中需要注意的问题进行了展望,以期为我国生态草牧业的发展和科学研究提供思路。
1 混播草地种间互作机理
物种之间存在复杂的相互作用,生物多样性的变化可以通过物种性状改变个体生理生态特性(生物途径)来直接调节生态系统过程。互补和竞争是种内和种间关系的两个方面,同时存在且同时发生[13]。互补效应体现在物种间的生态位分化和相互促进作用[14]。物种多样性的增加通常导致群落整体资源利用率更高,从而获得更高的生产力[15]。物种之间的生态位互补效应起到很大作用,即多样性的增加使群落中物种的功能特性如空间分布、根系深度、养分利用偏向性等的多样化增加,从而提升资源利用率,进而促进系统功能水平的提高[16]。例如,多样性高的系统中植物能更有效地利用空间和光照;深根系物种和浅根系物种组合,能较大限度地利用不同土层的水分和养分。然而物种间也存在着对光、水分、养分等的竞争,多维的环境因子(包括地上和地下)引起多维的竞争过程。同种间互补一样,物种间的竞争平衡对生态系统功能也起到同样重要的作用[17]。例如,在混播植物的整个生长季内,当竞争作用强度大于互利时,混播群落对资源的整体利用能力下降,反之,混播群落对资源的整体利用能力增加,具有混播优势[18]。因此,混播草地种间互作机理是决定混播草地发挥优势的科学前提,可为人工草地的管理措施提供科学依据,对于维持混播草地的持续稳定生产具有重要意义。
1.1 种间互补
地上部空间和光照的互补性被认为是植物物种多样性对植物生物量产生积极影响的作用机制之一。物种多样性的增加促使植物结构的互补以及群落中单个植物物种的地上空间利用的改变,进而影响群落冠层结构以及对空间和光的利用[19]。当植物物种数从1、2、4增加到32时,地上生物量和叶面积指数随植物物种数的对数呈线性增长,包含32种植物的群落较单一物种单位面积的光吸收率增加了44%[20]。黑燕麦(Avena strigosa)较一年生黑麦草(Lolium multiflorum)植株更大,且分蘖密度低,混播使黑燕麦植株之间的空隙被一年生黑麦草植株填补。这种群体互补性可能对叶片生长有积极的影响,因为每个物种较低的分蘖密度及其不同的结构使这种混播草地能够占据和利用不同的冠层空间,并有效地捕获入射光,进而提高了总叶片量和总生物量[21]。
研究表明,根系生长物候[22]、氮素偏好[23]和土壤生物特定相互作用[24-25]的物种差异都可能有助于混播系统的互补性和高产,通常认为物种根系深度的差异促进了地下垂直生态位互补,一方面可以减少对于同一土层养分的激烈竞争[26],另一方面能够更充分地利用土壤资源[27],例如,浅根和深根植物吸收水分和养分的主要深度存在显著差异[28]。全球气候变化背景下,干旱对草地生态系统功能和服务产生严重的负面影响受到极大关注[29-30]。Hoekstra等[31]通过同位素示踪发现,在干旱条件下,混播物种从深层土壤吸收示踪剂的比例都有所增加,但深根物种的增加幅度更大。在浅层土壤资源有限的情况下,深根植物有更好的能力进入更深的土壤层,从而获得水分和养分。另外,深根系植物的提水作用为浅根系植物扩展了可利用水资源,为表层土壤养分的溶解吸收创造了适宜的水分条件[32-33]。这表明,由于水分和营养吸收的灵活性,在混播系统中包含深根物种是应对土壤水分条件变化的重要策略。
氮素是草地农业系统植物生长的主要限制性养分。豆-禾混播草地,以其较高的生产力、营养价值往往成为许多地区人工草地建植的主要方式之一[34-35]。豆科植物的共生固氮替代无机氮肥的投入对草地农业系统减少污染和降低成本起到重要作用[36]。豆科植物通过共生固氮,一方面满足自身需求,另一方面可以通过根系接触、丛枝菌根真菌或者凋落物分解等方式将氮素转移给相邻的禾草[37-39](图1)。豆-禾混播提高草地生产力的同时减少了对氮肥的需求。豆-禾混播系统中,由于禾草比豆科植物对于土壤中的无机氮有更强的竞争能力,迫使豆科植物更多进行共生固氮[8]。例如,与禾草混播时,紫花苜蓿(Medicago sativa)约80%的氮来自生物固氮,高于紫花苜蓿单播[40]。混播中苜蓿为禾草提供氮[37],在不同的豆-禾混播组合中,从豆科植物向禾本科植物转移的氮量每年为10~75 kg·hm-2[41]。Malhi等[42]发现,在保证牧草产量、品质的前提下,紫花苜蓿-无芒雀麦(Bromus inermis)混播相比无芒雀麦单播可节约100 kg N·hm-2。Nyfeler等[35]研究表明,豆-禾混播对产量的积极影响并不完全依赖于共生固氮的直接作用(仅含66%和37%豆科植物的豆-禾混播系统可获得与纯豆科植物相同的共生固氮量),而是豆科植物的共生固氮以及豆-禾之间的氮素转移这两个过程共同作用的结果。
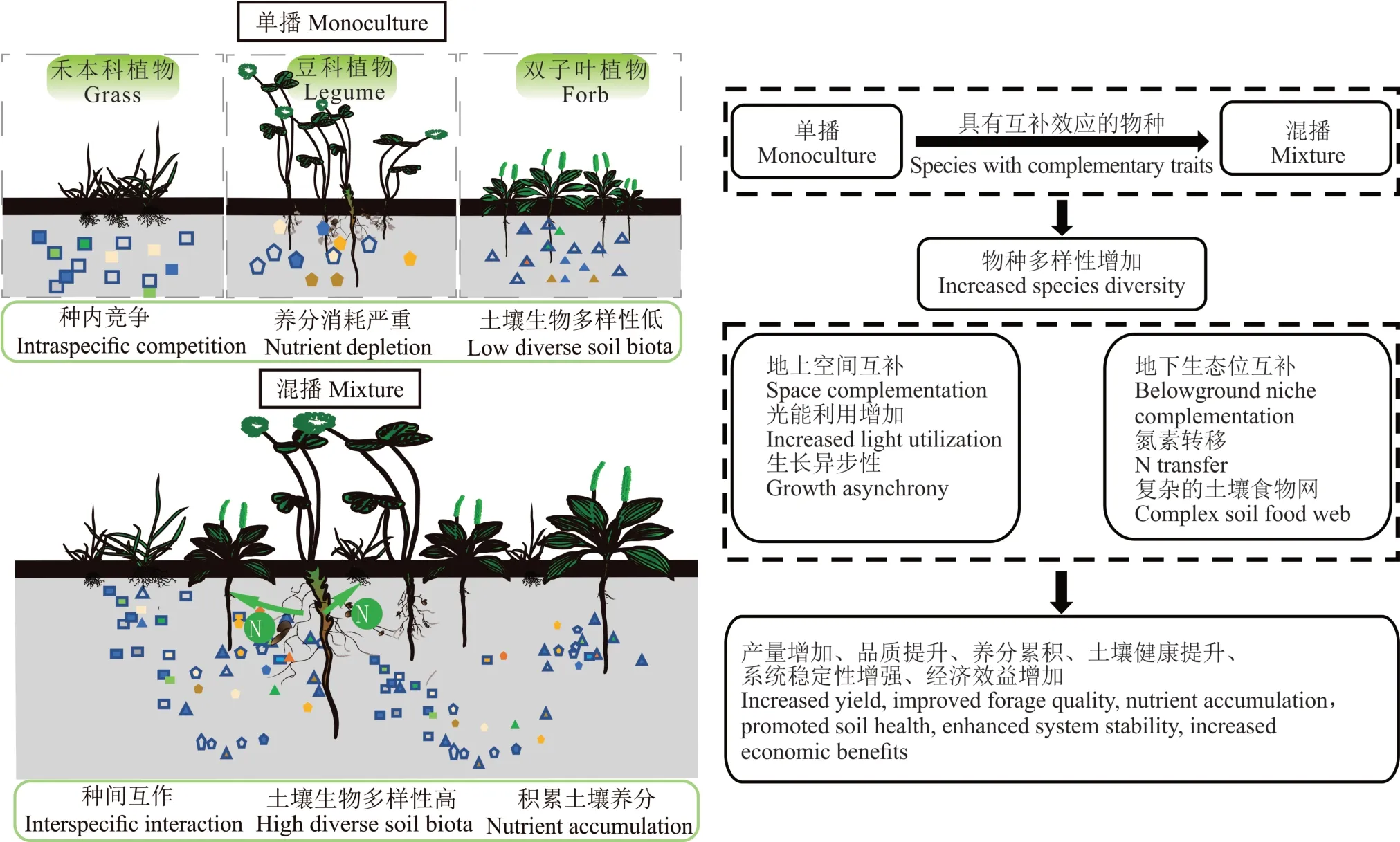
图1 牧草混播提高生态系统功能Fig.1 Forage mixture improves ecosystem functioning
物种生长的异步性促进了种间资源利用的时间互补性,例如在混合种植草地中,种间叶片生长的异步性可能会促使光截获的时间生态位分化[43],作物需水特征的差异产生整体系统水分需求的时间生态位分异,从而降低水分竞争,提高作物体系获取水分的能力[44]。群落中物种生产力峰值的不同步性,促使群落全年保持稳定的生产力水平[45]。Husse等[46]研究发现,与单一栽培相比,4种植物混播的草地,3年生长期内增产23%,混播草地产量与各物种间地上部生长的季节不同步性呈正相关关系,表明地上部季节性生长的不同步有利于高产和稳产。
1.2 相互竞争
物种间竞争也是调控群落结构[47]和生态系统功能的重要因素[48]。地上竞争控制光和空间的可用性,而地下竞争主要影响水和养分的供应。尽管目前还有许多争论,但物种之间的功能相似性和环境资源的可利用性常被认为是调节植物竞争强度、信号和结果的关键因素[49-50]。植株大小、生长速率和化感分泌物的产生等性状是影响植物竞争能力的重要因素[51]。植物间的竞争能力存在显著差异,可能导致明确的竞争层次和高度的竞争不对称(即抑制作用),最终导致竞争力差的物种一直处于弱势甚至被淘汰。无芒雀麦根茎型须根系可以保证其在浅层土壤中有效地吸收养分和水,刈割和放牧条件下,其主要依靠无性繁殖来扩张,增加其与周围植物的竞争力。分别与红三叶(Trifolium pratense)和红豆草(Onobrychis viciifolia)混播时,无芒雀麦的干物质产量所占总干物质产量的比例随着年限的增加而显著增加,相反,红三叶和红豆草干物质产量所占比例显著下降,特别是红三叶在建植后的第4年从混播中消失[52]。
禾草和豆科牧草在混播种植过程中往往出现一方逐渐消退,另一方逐渐占据优势的现象。生态位理论认为生态位分离是物种实现共存的一条途径。由于物种间的生态位差异,混合种植的个体在资源利用方面生态位重叠平均少于相应的单一种植。这种生态位重叠的减少使资源利用更有效。通常,禾本科植物对有效氮的竞争胜过豆科植物[53]。对于禾草与豆科牧草间对氮素的竞争权衡,只有当土壤氮含量通过氮循环达到禾草与豆科牧草各自竞争优势平衡的水平时,两者才可以共存。研究表明[54]无芒雀麦和紫花苜蓿混播,当不施加氮素时,紫花苜蓿通过生物固氮满足自身需求,且将氮素转移给无芒雀麦(地上部转移的氮素占无芒雀麦氮素含量的15.26%~29.92%),两者之间无明显的竞争效应,且相对总生物量显著大于1;当施加75和150 kg N·hm-2氮素时,紫花苜蓿的生物固氮比例被抑制,混播中无芒雀麦和紫花苜蓿同时竞争土壤氮素和肥料氮,且总生物量无显著的混播效应[54]。因此,不仅是种间互补,物种间的竞争平衡在增产等生态系统功能方面也具有重要作用。
2 混播草地的生态系统功能和生态系统服务
生态系统功能是构建系统内生物有机体生理功能的过程,侧重于反映生态系统的自然属性;生态系统服务是由生态系统功能产生的,基于人类的需要、利用和偏好,反映了人类对生态系统功能的利用[55]。由于物种之间存在复杂的相互作用,生物多样性的变化可以通过物种性状改变个体生理生态特性(生物途径)来直接调节生态系统过程,也可以通过改变有限资源的可获得性、微生境小气候及干扰机制等(非生物途径)来调控生态系统功能[56]。
草地生态系统不仅能够提供饲料,作为陆地生态系统一个重要的子系统在全球变化及生态系统功能发挥等方面也起着非常关键的作用[57-58]。由于植物与植物以及植物与土壤之间的相互作用关系,物种多样性和组成已被认为是影响草地生态系统功能和服务的重要因素[14,59-61]。对已有研究的分析发现,生物多样性总体上对生态系统功能和服务有积极影响[62]。另外,现代草地管理在追求增产提质的基础上也寻求提供多种生态系统功能和服务[63-64],牧草混播使物种多样性增加,有助于提高集约化种植草地的生态系统功能和服务[65]。
2.1 提高产量
生产力是栽培草地建植的主要目的和价值的直接体现。自然生态系统中生产力与物种多样性关系的试验最早被开展。大量生态学试验表明,生物多样性影响着生态系统的初级生产力及其他生态系统功能[59,66-67]。生态位互补效应(niche complementarity effect)和选择效应(selection effect)被认为是物种多样性影响生产力的两个主要机制[68-70]。生态位互补效应认为各物种获取不同资源的能力或者其他多种生态位上存在差异,物种多样性的增加导致群落整体资源利用率更高,从而获得更高的生产力[15]。选择效应即群落物种数的增加将增大从物种库中选择高产物种的可能性,高生产力物种的局部优势促进了整体初级生产力的提升[16,68,71]。
Tilman等[16]认为,物种间的差异和相互作用,使系统整体对环境的响应不同于各物种单独响应,例如,多物种资源竞争的情况下,多样性越大,物种特性差异越大,资源利用率越高,进而减少生态系统养分损失,导致土壤碳和其他养分储存的长期增加,这也进一步促进了随后生产力的提高。在为期7年的试验中,植物多样性和生态位互补性对生态系统功能的影响逐渐增强,16个物种混播草地的生物量是单作的2.7倍,而且生物量和土壤碳储量也高于单一栽培的最大值[59]。不仅在自然和半自然草地上如此,农业人工草地上的研究也得到一致的结果,Nyfeler等[8]在集约化经营草场上种植多年生黑麦草(Lolium perenne)、鸭茅(Dactylis glomerata)、红三叶、白三叶(Trifolium repens),比较单播和不同比例混播的产量发现,4种植物混播的产量比各物种单播的平均产量提高了106%,混播种植的产量比最高产的单一种植高出57%(超高产)。并且,多样性-生产力效应出现在多种不同物种比例和较低的氮肥用量下。近年来国内不同地区开展的多项不同物种组合的混播试验也表明,混播草地产量较单一栽培有很大提升[72-74]。这种生态位互补和种间正向互作等过程导致的物种多样性效应,在集约化饲草生产中具有重要意义和广泛适用性。
2.2 提高牧草品质
牧草品质一般包括营养价值、消化率及适口性等几个方面。其中,粗蛋白和粗纤维的含量是两项重要的营养指标,消化率高低影响家畜对营养物质的吸收,提高牧草粗蛋白含量、降低纤维素含量是提高牧草营养价值、改善牧草品质的重要内容[75]。
豆科植物和禾本科植物的营养成分有显著差异,豆科植物的粗蛋白、钙和磷含量比禾本科植物高,而纤维含量较低。一方面豆科牧草本身蛋白含量较高,另一方面豆科牧草在混播条件下能起到固氮作用,因此豆禾混播草地牧草具有较高的蛋白含量[9]。紫花苜蓿分别和垂穗披碱草(Elymus nutans)、无芒雀麦混播后,牧草产量和品质较两种禾草单播都有所提高[72]。研究表明[76],黑麦草、高羊茅(Festuca elata)、白三叶和红三叶混播使牧草整体酸性洗涤纤维浓度升高10%,尽管水溶性碳水化合物浓度降低20%~22%,但混播对净泌乳能量和粗蛋白产量仍有很强的正效应。与单一苜蓿干草相比,添加禾草后可提高混合饲料的纤维消化率,Veira等[77]研究表明,与苜蓿干草相比,将苜蓿-鸭茅混合物饲喂反刍动物时,腹胀明显减少,这可能是由于鸭茅中的中性洗涤纤维的消化率高于苜蓿,有助于调节瘤胃pH。
物种多样性的增加,不仅使混播牧草比单播含有较完全的营养成分,并且在整个收获期内,混播牧草营养质量也更稳定[78]。因为刈割后禾草较豆科植物的再生长过程更快,并且多数牧草生殖生长过程中由于茎和木质化纤维素的比例增加,纤维浓度增加,粗蛋白和易消化碳水化合物减少,消化率降低,但豆科植物的消化率下降率往往低于禾本科植物。因此,豆科作物的加入在生长后期改善了产量和饲料品质的季节分布[79]。
2.3 提高养分利用效率,改善土壤理化性质
单播与混播系统相比,往往会出现土壤养分消耗严重的现象。已有研究表明单播禾本科牧草的草地土壤表层碱解氮含量明显下降,单播豆科牧草则速效磷含量明显下降[80]。苜蓿比禾本科植物对磷的需求量更高,苜蓿单一种植会过度消耗土壤中的有效磷[81]。而豆-禾牧草混播有利于改善土壤速效氮、磷养分的供应。苜蓿与高羊茅混播后,与苜蓿单一种植相比,其对土壤无机磷的吸收减少[10]。苜蓿与鸭茅和无芒雀麦混播后土壤有效磷较单播有所增加[82]。红豆草与无芒雀麦混播与豆科和禾本科单播相比,其土壤容重降低,孔隙度和团聚体含量增加,土壤有机质、速效氮、速效磷和速效钾含量均有所增加[83]。
实际生产中,单播禾草需要施加大量的无机氮来维持产量,豆科牧草的加入,可通过共生固氮作用减少氮肥的使用,降低生产成本,减少温室气体排放和土壤中氮素淋溶[36]。另外,豆-禾混播通过地上生物量的积极影响进一步改变土壤的碳、氮收支[84],并且不同豆-禾比例导致土壤C∶N化学计量发生变化[85]。混播草地中不同植物功能类群互补效应促使了根系生物量的增加[59],例如,两种禾本科和双子叶植物混播第一年,根系生物量较单一种植增加了40%[86]。考虑到根系的高周转率,较高的根系产量将增加土壤中有机质输入[87],并进一步提高氮的矿化率和可利用性[88],土壤碳、氮的增加反过来又将对植物群落的高生产力产生积极的影响。多花黑麦草生长在前茬为豆科植物的土壤中较非豆科植物土壤中,可收获更高的氮产量和生物量,并且在多次刈割后仍然有效[89]。这种土壤遗留效应对不同草地和草-粮轮作可能会起到十分重要的作用。
2.4 调节土壤食物网,提高土壤健康
土壤生物群落是调节生态系统功能的关键因素,土壤生物多样性的丧失和土壤生物群落组成的简化将会影响植物多样性、凋落物分解、养分保持和养分循环等多种生态系统过程和功能[90-92]。现代农业大面积种植单一作物的做法降低了农田生态系统的生物多样性,导致土壤微生物数量下降、微生物多样性发生改变以及群落结构失衡等问题,并使土壤养分过度利用及严重流失[93]。近年来研究表明,植物个体对土壤微生物群落有着强大的影响,由于植物根系形态、根系分泌物[94]、土壤养分输入[95]等因素的差异,导致土壤生物与特定的植物物种有密切的联系[96],例如,由于禾本科和双子叶植物产生不同的根系分泌物,因而它们对土壤微生物群落有着显著不同的影响[97]。同样,土壤生物群落组成可能也受植物群落组成的调控。近年来在草地生态系统中的一些研究表明,植物多样性可通过驱动有机质的输入和调节土壤水分增加土壤微生物的生物量[98-99]。此外,长期草原生物多样性试验(the Jena experiment)[100]的结果表明,土壤细菌和真菌多样性随着植物多样性的增加而增加,并且植物群落组成对根系共生生物的群落组成有很大的影响。植物群落多样性对土壤生物的调节,可能有助于提升土壤健康水平,进一步促进地上植物的健康生长(图1)。
Lamb等[11]采用结构方程模型指出植物群落对土壤多样性和潜在功能的直接和间接影响,结果表明植物群落物种丰富度对古细菌有直接的正向影响,而植物均匀度的增加提高了细菌的丰度。在我国西北半干旱区的灰钙土中,紫花苜蓿和小黑麦(Triticale wittmack)间作可降低根际土壤pH值,明显改善小黑麦的根际土壤营养状况,进而调节了细菌群落结构[101]。有研究表明,尽管植物群落多样性促进了病原菌群落多样性,但同时降低了病原菌对植物个体的侵染水平[102]。混播种植系统中的丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF)孢子比单一种植的生存能力和与多种植物相互作用的能力更强,且更能显著提高白三叶的生物量。多样的植物群落可以维持土壤中AMF繁殖体的活力,这对土壤健康和生产力至关重要[103]。植物多样性控制着节肢动物的生物量和时间稳定性[104],可通过改变植物-食草动物和捕食者-食草动物的相互作用来限制草食动物。在欧洲和北美洲的两个长期草原生物多样性试验中,试验者收集了487个节肢动物食物网,研究其是否增加植物多样性以及如何减少食草动物对植物的影响。结果表明,在高多样性的混播种植中食草动物造成的损失不足单一种植的一半,并且揭示了植物多样性在有益于捕食者和降低平均食草动物食物质量的同时,来降低食草动物对植物的影响[105]。
2.5 提高系统稳定性
物种多样性对于维持生态系统在不断变化的环境中的稳定是必不可少的。从农业生产的角度来说,一味追求最高产量可能不是最优的选择,年际间产量稳定性以及群落对于环境变化的响应(包括抵抗力和恢复力)也很重要[79,106]。由于不同的植物物种对环境波动和干扰的反应不同,混播草地往往比单一种植草地的季节生长更均匀[7,107],总的群落或生态系统将更加稳定[108]。多样性对于在恒定或良好的环境条件下维持生态系统过程和功能可能显得并不那么重要,但在不断变化的条件下却起到了关键作用[15,109]。因此,选择具有互补功能性状的品种或基因型已被认为是提高产量和产量稳定性的有效策略[36]。
有研究[45,110]发现随着物种多样性的增加,植物群落生产力在不同年份和季节间变化均减小。生物多样性通过高产、物种异步性和保险效应增加时间稳定性[45,111],物种间的相互作用促进了生物多样性、生产力和时间稳定性。另外,持续时间较长的草地节约了重新播种费用,是一种经济可行的选择[112]。
深根系植物比浅根系植物更耐旱,通常豆科植物比非豆科植物更耐旱,而非豆科植物在干旱结束后恢复力更高。研究表明[12],豆科和非豆科植物间的协同作用提高了系统对于干旱胁迫的抵抗力以及干旱胁迫结束后的恢复力,这对于集约化管理草地在干旱环境下减少损失和确保长期产量稳定至关重要。冯鹏等[113]将4个不同苜蓿品种与无芒雀麦混播,对越冬率及根系各形态学指标的测定分析表明,混播处理越冬率均高于其单播处理;随着外界低温环境的胁迫,同一苜蓿品种混播处理根系直径、主根直径及侧根数均高于单播处理。
植物多样性与增加施肥和刈割频率等管理措施类似,可作为草地生态系统重要的生产因子[114]。混播草地由于物种多样性的增加,生物量、牧草品质、产量稳定性等生态系统功能的提升为农民增加了收入[115],改善土壤理化性质,提升土壤健康等多项生态系统功能和服务,对草地生态系统可持续发展起到了关键作用。
3 展望
草地生态系统经常受到粮食生产的农业扩展、放牧压力和气候变化等多种威胁。并且,由于近年来居民饮食结构的改变,草地生产需要跟上更高的肉和奶产量的要求,并应对和适应不断变化的气候[116]。因此迫切需要了解和管理草地生产性能和其他多种生态系统功能和服务。草地生态系统是研究生物多样性的理想生态系统,以往在自然草地生态系统的大量研究证明了物种多样性与多种生态系统功能和服务的正相关关系[12,59,117-118]。近年来,世界各地在集约化种植草地上的多项研究也表明,混播草地比其组成物种的单一种植的平均产量更高,牧草品质更好,经济收入增加,系统稳定性和土壤健康等多项生态系统功能均有所提升[46,114-115,119]。然而,驱动这些积极的混播生态系统功能关系的机制还不清楚[120],另外从天然或半天然草地研究中得出的生态学原理是否能转化到集约经营的人工草地还需要进一步验证。因此,本研究对我国未来混播草地生态系统功能研究提出以下展望:
混播草地涉及不同物种或功能类群之间的相互作用关系,其直接影响系统生产力和稳定性,例如竞争和互利在豆-禾混播群落中可同时存在,两者的相对大小因物种组成和建植年限而改变,并进一步影响群落结构,进而影响混播优势。互补和竞争是同一种间关系的两个方面,两种相互作用在空间和时间上都不是恒定的,并且在很大程度上受到播种比例、刈割制度和水分状况等管理措施的影响[34]。因此,了解影响种间互补和竞争强度的因素,明确混播系统对土壤资源的互补和竞争效果,对于理解混播系统的组合,维持混播草地的持续稳定生产至关重要[121]。
已有的研究对种间相互作用的地上部关注较多,迄今为止,还不清楚地上高产和互补性是否反映在地下部[86]。地上地下互作影响植物生长和群落结构,影响植物地上竞争能力的一些特征可能会影响到其地下竞争能力,例如,地下竞争能力与传向根部的碳供应能力有关,植物对光的竞争(获取碳)将会影响其向根部的碳供应能力,进而影响植物间地下竞争。草地植物物种与土壤群落成员之间存在着特定的联系是决定草地土壤生物多样性格局的关键因素。因此,进一步了解混播草地种间互作对土壤养分和土壤碳、根系养分竞争、根系与地上部分耦联机制的影响[122],以及与土壤群落的调控[123-125],对混播草地调节生态系统功能的机制至关重要。
与单一种植相比,优化混播生态系统功能需要选择合适的物种,确定植物种类特征的有效组合[112]。国内开展的相关研究主要集中于豆-禾混播,涉及种类较少,且缺少同功能类群和不同功能类群多物种的混播。目前尚不清楚的是,在豆-禾混播系统中,是否可以通过增加物种的功能群(如双子叶植物)来进一步提高生产力。已有研究人员提出,由于矿物质高,适口性好,许多双子叶草本植物,例如长叶车前(Plantago lanceolata)具有提高混播草地产量和饲料质量的潜力[41]。因此,为了增强多物种群落的生态系统功能,应该有针对性地选择物种以包含特定的特征或最大限度地提高性状多样性[126],将基于植物功能性状多样性的方法与植物物种多样性的方法结合起来,在草地农业生产中选择对产量和生态功能起到关键作用的物种性状,例如:1)高产潜力;2)在氮素获取和利用方面实现互补性的能力[35];3)物种发育的时间差异,以改善早期建立和保持种间相互作用,进一步增加对混播草地植物多样性-生态系统功能关系的认识[127]。
建植人工草地是为了加快退化草地的恢复以及发展草地畜牧业。放牧也是草地利用和管理的最常用方式。以往在天然草地上的研究发现,放牧对草地生态系统的影响体现在诸多方面,例如,由于植物适口性、耐牧性的差异,放牧制度、放牧强度和放牧频率都能影响植物群落结构组成[128-130],动物通过踩踏、排泄等行为改变土壤物理、化学性状[131-132]。草-畜互作对于混播人工草地生产、稳定性及生态系统可持续利用同样起到极其重要的作用。明确放牧对混播人工草地生态系统中“土壤-植物-动物”之间互作和耦合效应的影响,为促进牧草人工混播在种养结合新的畜牧业经营模式中的发展提供理论基础。
另外,国外牧草产业的发展和多地联网[133],大时间尺度[134]得到较为可靠普适的研究结果有关。国内多数研究是在较短的时间尺度和某一特定区域开展,缺少多地的联网合作,多样性效应在大的时间和空间尺度上是否一致还不确定。因此,未来草地农业生态系统的研究还应在较大时间和空间尺度上、高物种、高功能类群多样性梯度上开展工作,进一步探究草地生态系统如何提供持久和一致的高产和生态系统功能,促进我国集约经营的草地与可持续集约化农业的发展。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
现代畜牧科技(2021年11期)2021-12-21 06:11:24
食品安全导刊(2021年20期)2021-08-30 06:40:50
现代畜牧科技(2021年7期)2021-07-28 06:40:30
今日农业(2021年10期)2021-07-28 06:28:00
西藏农业科技(2019年1期)2019-07-25 00:37:00
——黔南扁穗雀麦
贵州农业科学(2019年1期)2019-02-20 02:09:54
杂草学报(2015年2期)2016-01-04 14:57:54
鹿鸣(2015年7期)2015-05-30 10:48:04
水生生物学报(2015年1期)2015-02-28 16:01:05
