
结实期低温胁迫对水稻强、弱势粒淀粉形成与积累的影响
2022-09-15 02:17:48陈红阳贾琰赵宏伟瞿炤珺王新鹏段雨阳杨蕊白旭王常丞
中国水稻科学 2022年5期
陈红阳 贾琰 赵宏伟 瞿炤珺 王新鹏 段雨阳 杨蕊 白旭 王常丞
结实期低温胁迫对水稻强、弱势粒淀粉形成与积累的影响
陈红阳 贾琰*赵宏伟*瞿炤珺 王新鹏 段雨阳 杨蕊 白旭 王常丞
(寒地粮食作物种质创新与生理生态教育部重点实验室/东北农业大学,哈尔滨 150030;*通信联系人,email:jiayan_cool@126.com, hongweizhao@163.com)
【目的】研究结实期低温胁迫对水稻强、弱势粒淀粉组分含量、积累速率及关键酶活性的影响,明确淀粉合成关键酶活性变化对淀粉积累速率的调控效应,探究强、弱势粒淀粉形成积累差异对水稻产量的影响。【方法】以耐冷型品种东农428和冷敏型品种松粳10为试验材料,设置1个常温处理(白天温度28℃,14h/夜间温度22℃,10 h,7 d)和4个低温处理(17℃,低温处理时间分别为1、3、5、7 d),分析了结实期低温胁迫对强、弱势粒淀粉组分积累、合成关键酶活性、水稻产量及构成因素的影响,并探讨了灌浆期各阶段淀粉积累差异与酶活性变化的关系。【结果】与对照相比,结实期低温胁迫降低了两个品种水稻强、弱势粒中腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)、可溶性淀粉合成酶(SSS)、淀粉分支酶(SBE)的峰值活性以及支链淀粉和总淀粉含量,提高了齐穗后28~38 d 低温处理3、5、7 d的颗粒结合型淀粉合成酶(GBSS)活性和直链淀粉含量。与对照相比,各低温处理酶活性最高和淀粉积累最快的时间均有不同程度的推迟,低温处理7 d的影响最大,强、弱势粒低温处理7 d的支链淀粉和总淀粉含量分别在齐穗后13、18 d降幅最大,直链淀粉含量在28 d增幅最大。相关分析表明,强、弱势粒直链淀粉含量及强势粒支链淀粉、总淀粉含量与其最大积累速率呈极显著正相关,弱势粒支链淀粉、总淀粉含量还受到其最大积累速率出现时间的影响。AGPase、GBSS、SSS、SBE活性变化与淀粉积累速率和积累时间早晚密切相关,对淀粉及淀粉组分含量的变化有着明显的影响。同时,结实期低温胁迫显著降低了水稻的千粒重、结实率和产量,且随低温处理天数的增加降幅逐渐增大。结实期低温胁迫对弱势粒中淀粉合成关键酶活性变化影响大于强势粒,弱势粒淀粉合成积累减慢,含量降低,导致水稻千粒重显著下降,产量降低。【结论】就品种而言,耐冷型东农428具有较高的淀粉合成关键酶活性,淀粉及其组分含量较高,在低温胁迫下产量能维持在较高的水平。因此,强、弱势粒淀粉合成关键酶活性对淀粉合成起着非常关键的调控作用,淀粉组分和含量及其变化对产量有着十分重要的影响。
水稻;结实期;低温;淀粉合成
黑龙江作为我国主要粳稻产区,在水稻种植面积、生产总量方面均居于我国首位,是我国发展水稻生产的优势区域[1]。21世纪以来,受环境、热量、气候等条件变化的影响,黑龙江省夏季低温冷害频繁发生,严重影响北方粳稻的高产稳产[2-4]。
结实期是影响水稻籽粒充实程度和粒重,决定水稻产量形成的关键时期[5]。水稻的灌浆过程主要是籽粒淀粉合成积累的过程,淀粉的含量与颖花在穗上的着生部位有密切关系[6]。一般情况下,稻穗中上部早开花的强势粒淀粉积累较快,充实好,粒重大;稻穗下部迟开花的弱势粒淀粉积累较慢,充实差,粒重小[7-9]。籽粒淀粉积累主要依赖于各种酶促反应的协同,腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)、颗粒结合型淀粉合成酶(GBSS)活性、可溶性淀粉合成酶(SSS)、淀粉分支酶(SBE)是淀粉代谢的关键酶,对淀粉合成起着重要的调节作用[10-12]。AGPase负责催化1-磷酸葡萄糖(G-1-P)形成腺苷二磷酸葡萄糖(ADPG),为合成淀粉提供原料,是决定淀粉合成速率和最终的淀粉合成量的限速酶[13]。由基因编码的GBSS在不影响支链淀粉合成的情况下延长直链淀粉聚合物,控制直链淀粉合成[14]。SSS和SBE通过催化葡聚糖产生新分支和延伸链长的方式共同完成支链淀粉的合成[15]。
温度对水稻籽粒淀粉合成积累的影响较大,特别是结实期温度是影响水稻产量和品质的首要环境因子[16]。有研究表明[17],籽粒自身生理活性,即淀粉合成关键酶是限制籽粒充实的重要因素之一。夏楠等[18]研究发现,结实期冷水胁迫显著降低了淀粉合成酶和淀粉分支酶活性,影响淀粉的前期合成及后期积累。沈鹏等[19]认为腺苷二磷酸葡萄糖焦磷酸化酶和可溶性淀粉合成酶对温度反应迟钝,淀粉分支酶对温度变化比较敏感,温度过高过低均影响该酶活性宋兴年[20]研究发现弱势粒酶活性随生育期延长呈先升高后下降趋势,夜间高温对酶活也有一定抑制作用。已有研究表明,低温会导致水稻籽粒淀粉含量降低,使籽粒中直链淀粉与支链淀粉的比例发生改变,中、低直链淀粉品种在低温胁迫下籽粒直链淀粉含量升高,结实期低温对直链淀粉积累有显著影响[21]。低温使水稻每穗实粒数、结实率、千粒重显著降低,影响水稻产量[22-24]。付景等[25]研究发现,灌浆期籽粒中淀粉合成相关酶活性与灌浆速率呈正相关,酶活性降低导致其弱势粒灌浆速率小、充实差、粒重低,不能形成有效的经济产量,严重限制了水稻产量潜力的发挥。
综上所述,国内外关于温度对水稻籽粒淀粉形成积累及产量影响的报道已有很多[26-28],前人关于水稻低温冷害的研究大多采用冷水灌溉的方式,分析对不同生育时期中部籽粒淀粉合成及产量的影响[29, 30],但针对结实期低温对强、弱势粒淀粉组分形成积累差异及合成关键酶的调控机理方面研究较少。因此,本研究以黑龙江省水稻耐冷品种东农428和冷敏感品种松粳10为试验材料,研究结实期低温胁迫对水稻强、弱势粒淀粉组分含量、积累速率及关键酶活性的影响,明确淀粉合成关键酶活性变化对淀粉积累速率的调控效应,探究强、弱势粒淀粉形成积累差异对水稻产量形成的影响,以期丰富水稻耐冷生理特性研究,为黑龙江省水稻高产及预防低温冷害提供理论依据。
1 材料与方法
1.1 试验材料
选择生育期相近,耐冷性存在显著差异的两个水稻品种东农428(耐冷型)和松粳10(冷敏感型)为试验材料。
1.2 试验设计
试验于2020和2021年在东北农业大学农学院人工气候室及盆栽场进行。4月21日播种,5月26日移栽至盆栽场。选择无渗漏的30 cm×28 cm塑料盆钵,每盆装过筛混匀黑土10 kg,盆栽密度设置为4穴/盆,每穴3株。人工气候室面积为50 m2,光照强度为30 000 lx,光照时间5:00-19:00,二氧化碳浓度为0.04%,相对湿度为75%。
试验采用随机排列,在齐穗期(水稻植株平均抽穗 80%左右)选取长势一致的植株挂牌标记,每个品种180盆,共计360盆。将每品种中的60盆移入常温人工气候室处理7 d,记为D0(对照),白天温度设置为28℃,夜间温度22℃;将每品种其余的120盆移入人工气候室,处理温度为全天17℃(接近寒地粳稻灌浆结实期冷害温度),处理天数为分别1 d、3 d、5 d、7 d,依次记为D1、D3、D5、D7,各低温处理10盆,3次重复。低温处理结束后立即移至常温人工气候室,全部处理结束后将所有水稻移至盆栽场生长。两个气候室除温度设置不同外,其余生长条件均相同。
1.3 试验方法
1.3.1 强、弱势粒直链、支链及总淀粉含量
待全部处理结束后第2天开始取样,之后每5 d取样一次,各取3穴,3次重复。将穗部籽粒按强、弱势粒分装。强势粒取最上部3个一次枝梗上的籽粒,弱势粒取穗最下部3个二次枝梗上的籽粒,剔除空粒。于105℃下杀青30 min,并于80℃恒温条件下烘干至恒重。粉碎,过100目筛,存放在干燥器内,等待测定。
参考《粮油籽粒品质及其分析技术》[31],采用双波长法测定淀粉含量。其中,测定支链淀粉的主波长为550 nm,参比波长为730 nm;测定直链淀粉的主波长为620 nm,参比波长为430 nm。
1.3.2 强、弱势粒淀粉合成关键酶活性
取样方法同1.3.1。用液氮冷冻处理并移至−80 ℃冰柜中备用。
参照李太贵等[32]方法测定淀粉分支酶(SBE)活性。参照Douglas等[33]、程方民等[34]方法,采用相应试剂盒测定腺苷二磷酸焦磷酸化酶(AGPase)、淀粉合成酶(SSS)、颗粒型淀粉合成酶(GBSS)活性,每个样本重复测定3次。
1.3.3 产量及产量构成因素
于成熟期各处理选取长势一致的植株3穴进行考种,测定有效穗数、每穗实粒数、千粒重、结实率并计算。同时,各处理取3盆,收获其全部稻穗,脱粒称量并测定实际产量。
1.4 计算方法
1.4.1 淀粉积累速率
淀粉积累速率(%/d)=(2−1)/(2−1)。其中,1为1时期淀粉含量,2为2时期淀粉含量。
1.4.2 低温反应指数
低温胁迫下的水稻淀粉合成关键酶活性变化用低温反应指数(CRI)[35]说明,计算公式如下:
低温反应指数(CRI)=低温处理区性状表型值/对照区性状表型值×100%。
1.5 数据分析
采用 Microsoft Excel 2019整理数据,SPSS 26 统计分析数据,Origin 2021作图软件绘图。
2 结果与分析
2.1 结实期低温胁迫对水稻强、弱势粒淀粉形成积累的影响
2.1.1 结实期空气低温对水稻强、弱势粒直链淀粉积累的影响
由图1可见,水稻强、弱势粒直链淀粉含量随生育进程的推进呈上升趋势。与对照(D0)相比,齐穗后8~23 d,低温处理下东农428和松粳10的强、弱势粒直链淀粉含量显著下降(除弱势粒第8天),且降幅随处理天数增加逐渐增大;齐穗后28~38 d,低温处理3、5、7 d(D3、D5、D7)两个品种的强、弱势粒中直链淀粉含量显著升高(除东农428的强势粒在齐穗33、38 d D3处理)。结合方差分析可知,齐穗后8~38 d强势粒直链淀粉含量处理间差异极显著,齐穗后13~38 d弱势粒直链淀粉含量处理间差异极显著。与D0相比,齐穗后13 d强势粒D7处理降幅最大,东农428和松粳10分别为46.76%和52.33%;齐穗后18 d弱势粒D7处理降幅最大,东农428和松粳10分别为38.67%和43.95%;齐穗后28 d强、弱势粒D7处理直链淀粉含量增幅最大,东农428强、弱势粒增幅分别为7.20%和6.63%,松粳10分别为8.54%和11.11%。
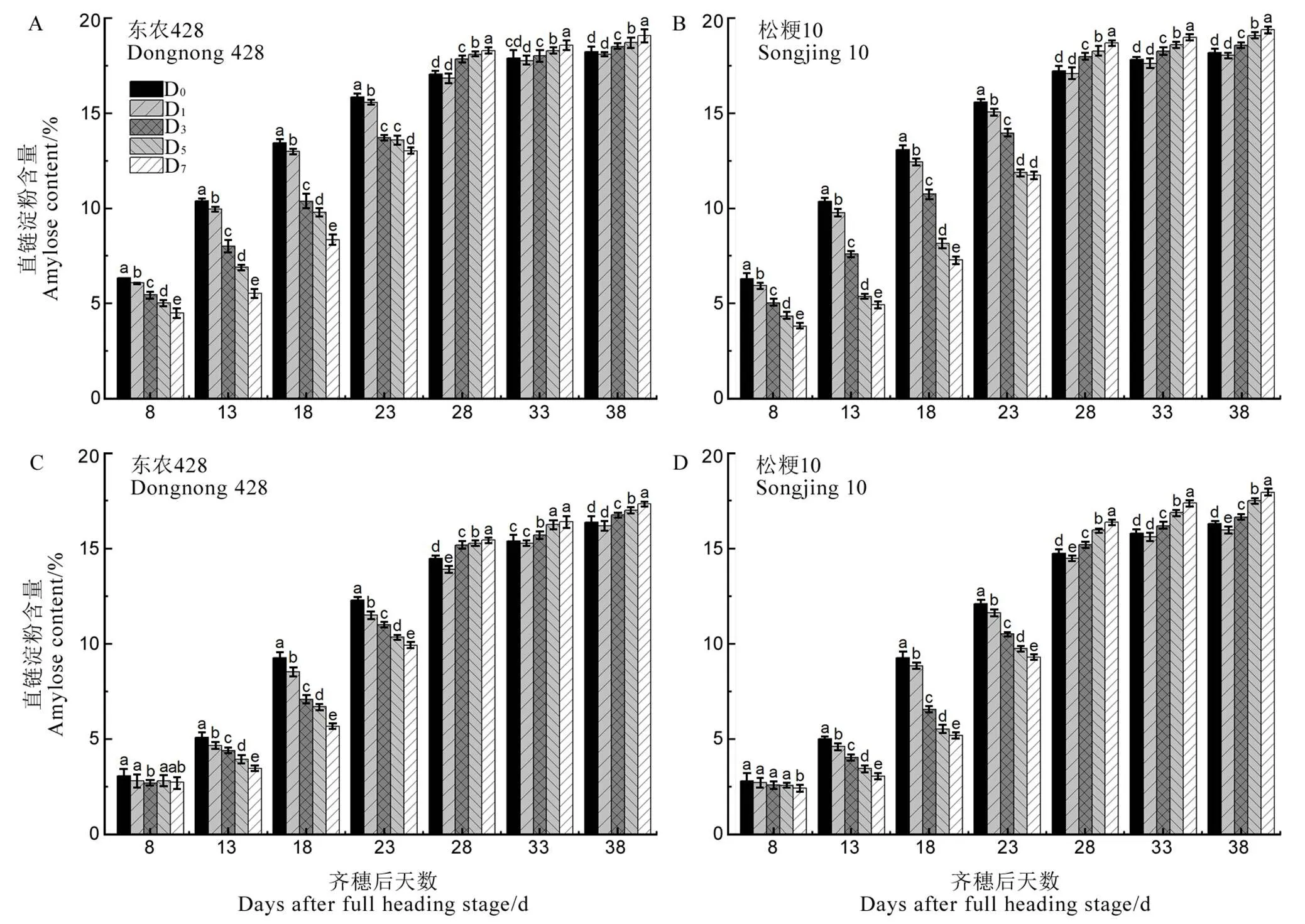
柱状图上不同小写字母表示处理间差异达5%显著水平。A、B-强势粒,C、D-弱势粒。D0、D1、D3、D5、D7分别代表低温(17℃)处理0(对照)、1、3、5、7 d。下同。
Fig. 1. Effect of low temperature stress on amylose content of superior and inferior grains during grain filling period.
由表1可知,强、弱势粒直链淀粉积累速度在处理和品种间存在极显著差异(除强势粒33~38 d)。齐穗后8~13 d,D0和D1处理强势粒直链淀粉快速积累,且D1积累速率显著慢于D0。齐穗后23~28 d,D3、D5、D7强势粒直链淀粉快速积累,积累速率显著高于D0,但较D0推迟15 d。齐穗后13~18 d,D0和D1处理弱势粒直链淀粉快速积累,两者间差异不显著。齐穗后23~28 d,D3、D5、D7弱势粒直链淀粉快速积累,积累速率显著高于D0,但较D0推迟10 d。以上结果表明,较长时间低温胁迫(D3、D5、D7)显著提高了强、弱势粒直链淀粉积累速率,使直链淀粉含量升高。
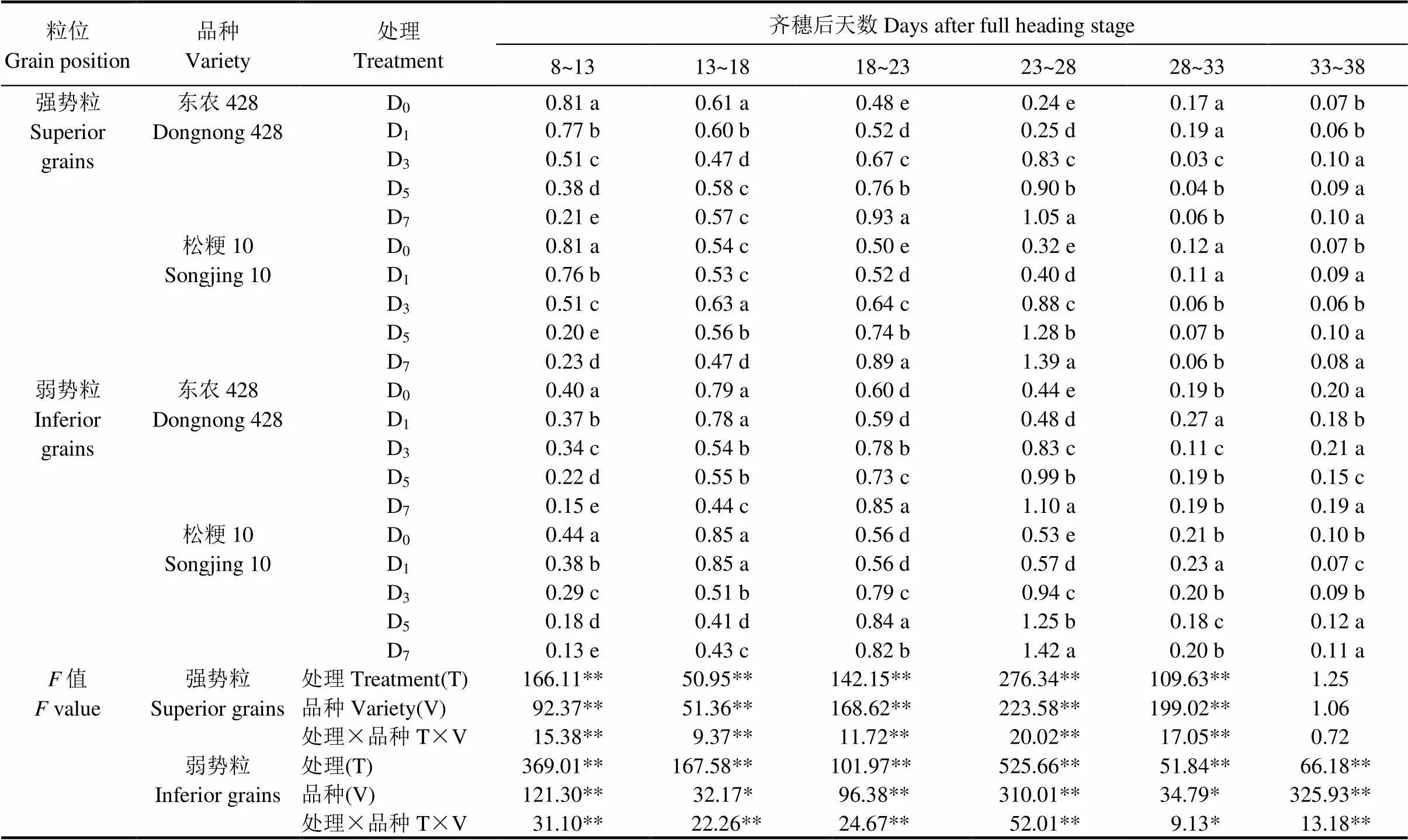
表1 结实期低温胁迫下直链淀粉积累速率
*表示0. 05显著水平;**表示0.01显著水平。
*represents significant difference at<0.05;** represents significant difference at<0.01.
2.1.2 结实期低温胁迫对水稻强、弱势粒支链淀粉积累的影响
由图2可见,水稻强、弱势粒支链淀粉含量随生育进程的推进呈增加趋势。与D0相比,低温处理下强、弱势粒支链淀粉含量显著下降(除弱势粒齐穗后8 d各处理,东农428弱势粒齐穗后38 d D1处理和强势粒齐穗后28、33、38 d D1处理),且降幅随处理天数增加逐渐增大。结合方差分析可知,强势粒支链淀粉含量处理间差异极显著。与D0相比,齐穗后13 d D7支链淀粉含量降幅最大,东农428和松粳10分别为14.06%和37.77%。弱势粒支链淀粉含量在齐穗后13~38 d处理间差异极显著,与D0相比,齐穗后18 d D7支链淀粉含量降幅最大,东农428和松粳10分别为28.73%和35.58%。
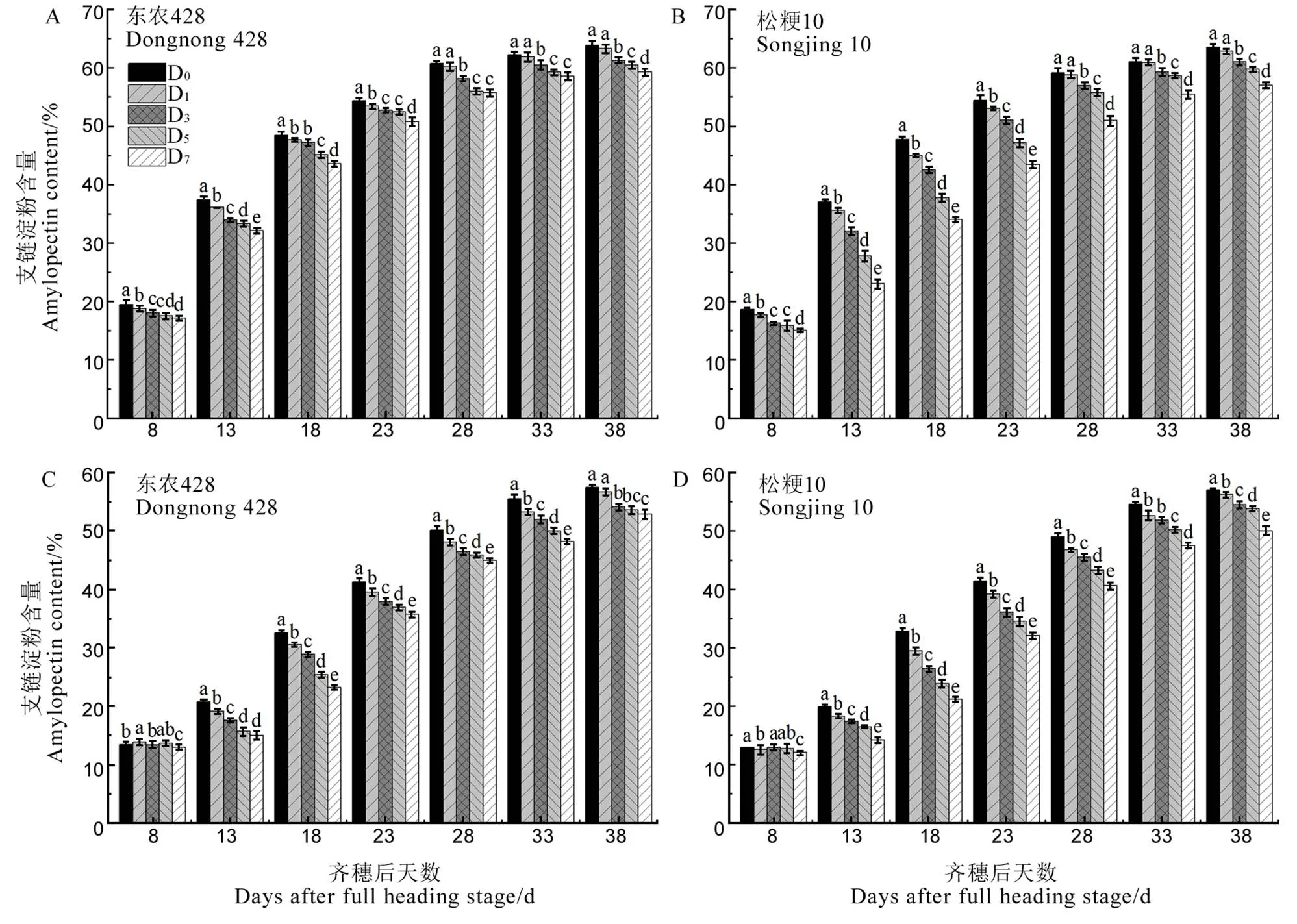
图2 结实期低温胁迫对强、弱势粒支链淀粉含量的影响
Fig. 2. Effect of low temperature stress on amylopectin content of superior and inferior grains during grain filling period.

表2 结实期低温胁迫下支链淀粉积累速率
由表2可知,强、弱势粒支链淀粉积累速率在处理和品种间存在极显著差异。除松粳10 D7处理外,其余各处理强势粒支链淀粉均在齐穗后8~13 d快速积累,且各低温积累速率显著低于D0,而松粳10 D7处理强势粒支链淀粉在齐穗后13~18 d快速积累,较D0推迟5 d。齐穗后13~18 d,D0、D1处理弱势粒支链淀粉快速积累,D1积累速率显著低于D0。齐穗后18~28 d,D5、D7弱势粒支链淀粉快速积累,较D0推迟5 d,而松粳10在处理3 d时就出现积累延后现象。以上结果表明,低温显著降低强势粒支链淀粉积累速率,导致最终支链淀粉含量下降;长时间低温胁迫下(D5、D7),弱势粒支链淀粉积累速率出现升高趋势,但淀粉积累时间后移,积累进程缩短,导致弱势粒支链淀粉含量降低。
2.1.3 结实期低温胁迫对水稻强、弱势粒总淀粉积累的影响
由图3可知,水稻强、弱势粒中总淀粉含量随生育进程的推进呈升高趋势。与D0相比,低温处理强、弱势粒总淀粉含量显著下降(除强势粒齐穗后33~38 d D1处理和弱势粒齐穗后8 d各低温处理),且随处理天数增加降幅逐渐增大。结合方差分析可知,强势粒总淀粉含量处理间差异极显著。齐穗后13 d D7处理总淀粉含量降幅最大,东农428和松粳10分别为21.17%和40.95%。弱势粒总淀粉含量在齐穗后13~38 d处理间差异极显著,齐穗后18 d D7处理总淀粉含量降幅最大,东农428和松粳10分别为30.93%和37.42%。
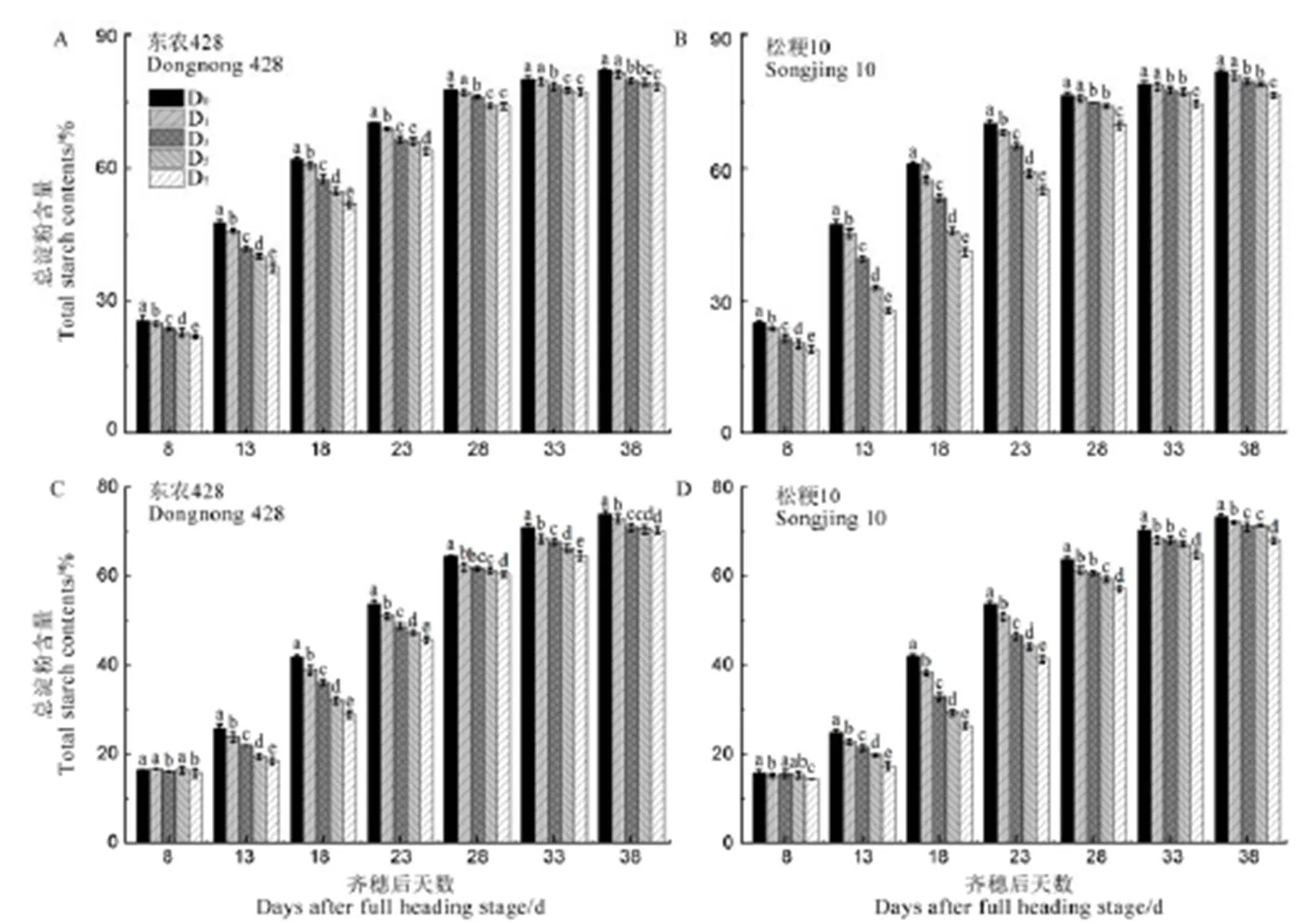
图3 结实期低温胁迫对强、弱势粒总淀粉含量的影响
Fig. 3. Effect of low temperature stress on total starch contents of superior and inferior grains during grain filling period.
表3 结实期低温处理下总淀粉积累速率
Table 3. Accumulation rate of total starch under low temperature treatment during grain-filling period. %/d
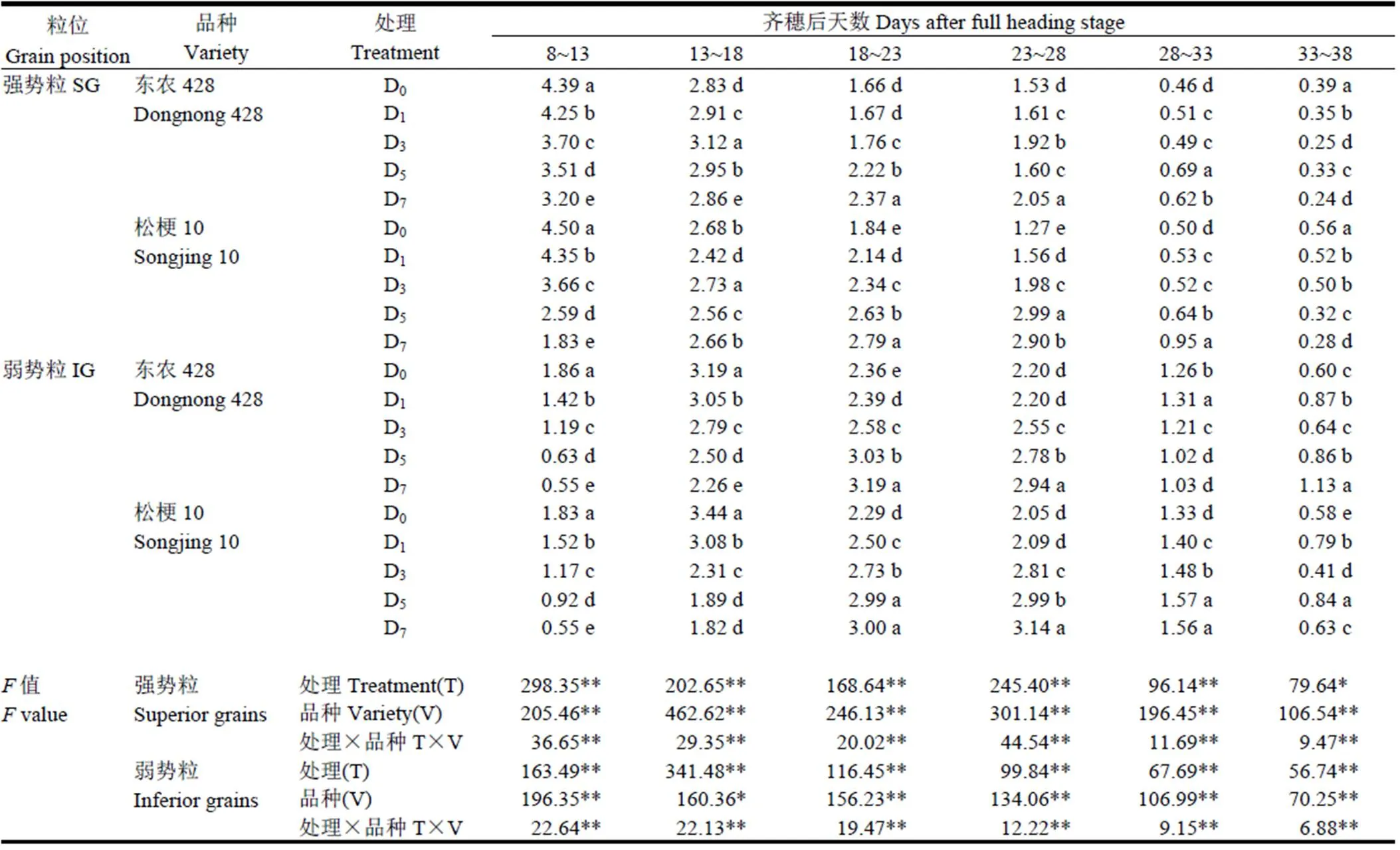
Fig. 4. Changes of AGPase activity of superior and inferior grains under low temperature treatment during grainfilling period.
由表3可知,强、弱势粒总淀粉积累速度在处理和品种间存在极显著差异。东农428强势粒各处理及松粳10强势粒D0、D1、D3处理总淀粉含量在齐穗后8~13 d快速积累,而松粳10强势粒D5、D7处理总淀粉含量则在23~28 d快速积累,较D0推迟15 d,且各低温处理总淀粉积累速率显著低于D0。东农428弱势粒D0、D1、D3及松粳10弱势粒D0、D1总淀粉含量在齐穗后13~18 d快速积累,东农428弱势粒D5、D7处理总淀粉含量则在18~23 d快速积累,较D0推迟5 d;松粳10弱势粒D3、D5、D7处理总淀粉含量在23~28 d快速积累,但较D0推迟10 d。
2.2 结实期低温胁迫对水稻强、弱粒淀粉合成关键酶活性的影响
2.2.1 腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)活性
由图4可见,水稻强、弱势粒中AGP焦磷酸化酶活性随生育进程推进呈先升高后下降的单峰曲线变化。与D0相比,各低温处理强、弱势粒AGPase峰值活性显著下降,且随处理天数增加降幅逐渐增大,各低温处理间差异显著。齐穗后13 d D0、D1、D3处理强势粒AGPase活性最高,D5、D7则在齐穗后18 d活性最高,较对照推迟5 d。齐穗后18 d东农428各处理和松粳10的D0、D1、D3处理弱势粒AGPase活性最高,松粳10的D5、D7处理则在齐穗后23 d活性最高,较D0推迟5 d。
AGPase峰值活性因品种耐冷性不同而存在差异,东农428强、弱势粒AGPase峰值活性低温反应指数(CRI)分别为85.21%、86.49%、88.44%、96.56%和75.82%、83.11%、87.49%、96.88%,松粳10强、弱势粒AGPase峰值活性CRI分别为72.33%、77.77%、81.30%、93.79%和68.29%、75.98%、81.53%、91.79%,AGPase峰值活性CRI小于东农428,说明松粳10对低温抵御能力较弱。
2.2.2 颗粒结合型淀粉合成酶(GBSS)活性
由图5可见,2个供试品种强、弱势粒中GBSS活性随生育进程推进呈先升高后下降的单峰曲线变化,与D0相比,强、弱势粒D1处理GBSS峰值活性显著下降,D3、D5、D7处理GBSS峰值活性显著升高且增幅随处理天数增加逐渐增大,强、弱势粒GBSS峰值活性处理间差异显著。齐穗后23 d D0、D1强势粒GBSS活性最高,D3、D5、D7处理则在齐穗后28 d活性最高,较D0推迟5 d。齐穗后28 d各处理弱势粒GBSS活性最高。

图5 结实期低温胁迫下强、弱势粒GBSS活性的变化
Fig. 5. Changes of GBSS activity of superior and inferior grains under low temperature treatment during grain filling period.
GBSS峰值活性因品种耐冷性不同而存在差异,东农428强、弱势粒GBSS峰值活性CRI分别为95.71%、105.23%、111.14%、114.98% 和93.76%、109.10%、115.06%、116.06%,松粳10强、弱势粒GBSS峰值活性CRI分别为94.01%、108.32%、113.63%、118.66%和91.95%、112.68%、122.73%、132.21%,D1处理GBSS峰值活性CRI小于东农428,D3、D5、D7处理GBSS峰值活性CRI大于东农428,说明松粳10对低温抵御能力较弱,受影响大。
2.2.3 可溶性淀粉合成酶(SSS)活性
由图6可见,2个供试品种强、弱势粒中SSS活性随生育进程推进呈先升高后下降的单峰曲线变化。强、弱势粒SSS峰值活性处理间差异显著(除东农428强势粒D7)。与D0相比,各低温处理强、弱势粒SSS峰值活性显著下降,且随胁迫天数增加降幅逐渐增大。齐穗后13 d D0、D1、D3强势粒SSS活性最高,D7处理则在齐穗后18 d活性最高,但较D0推迟5 d,而松粳10低温处理齐穗后5 d就出现峰值推迟现象。齐穗后18 d D0、D1弱势粒SSS活性最高,D5、D7处理则在齐穗后23 d活性最高,较对照推迟5 d,而松粳10强势粒胁迫3 d就出现峰值推迟现象。
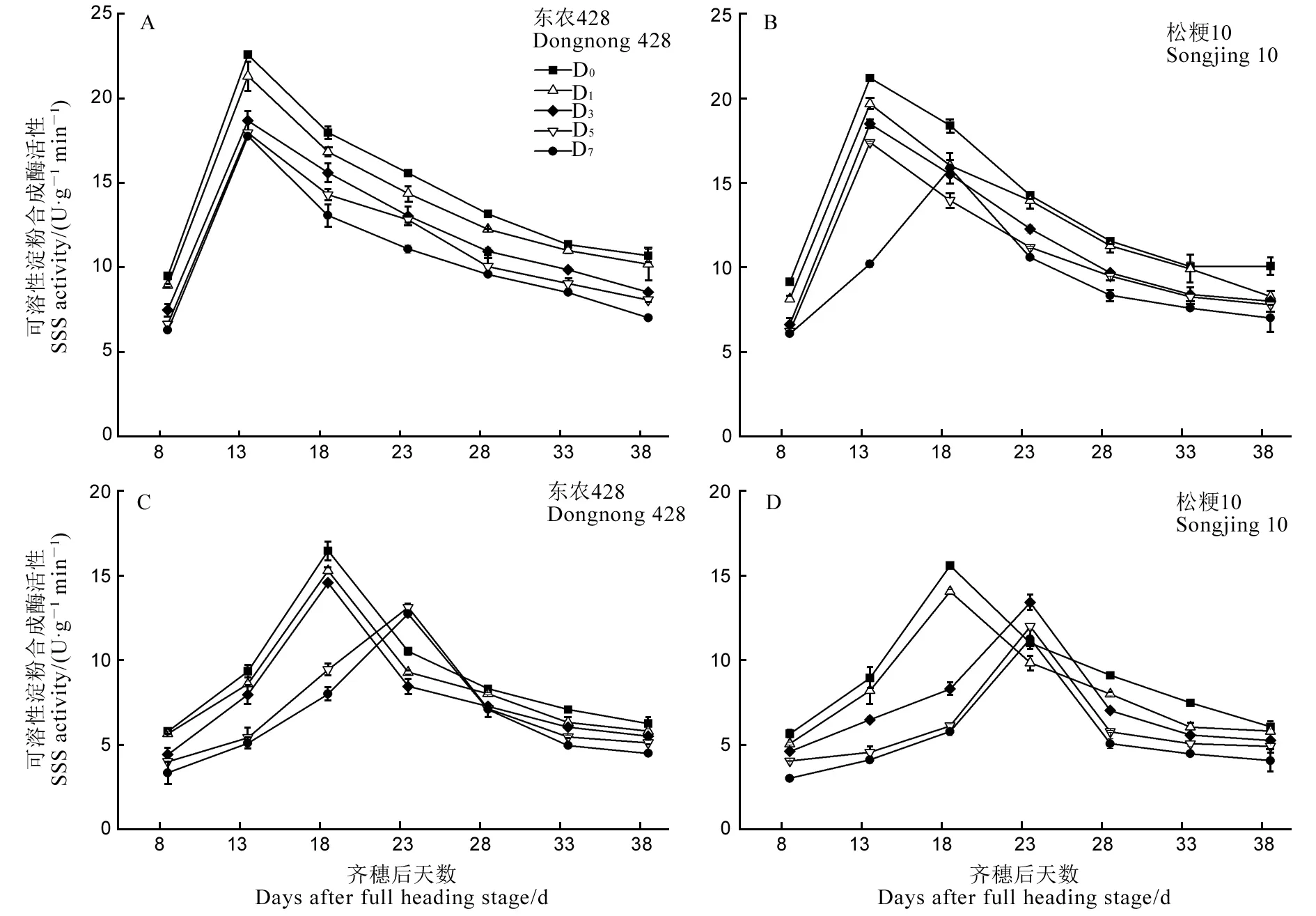
图6 结实期低温胁迫下强、弱势粒SSS活性的变化
Fig. 6. Changes of SSS activity of superior and inferior grains under low temperature treatment during grain filling period.
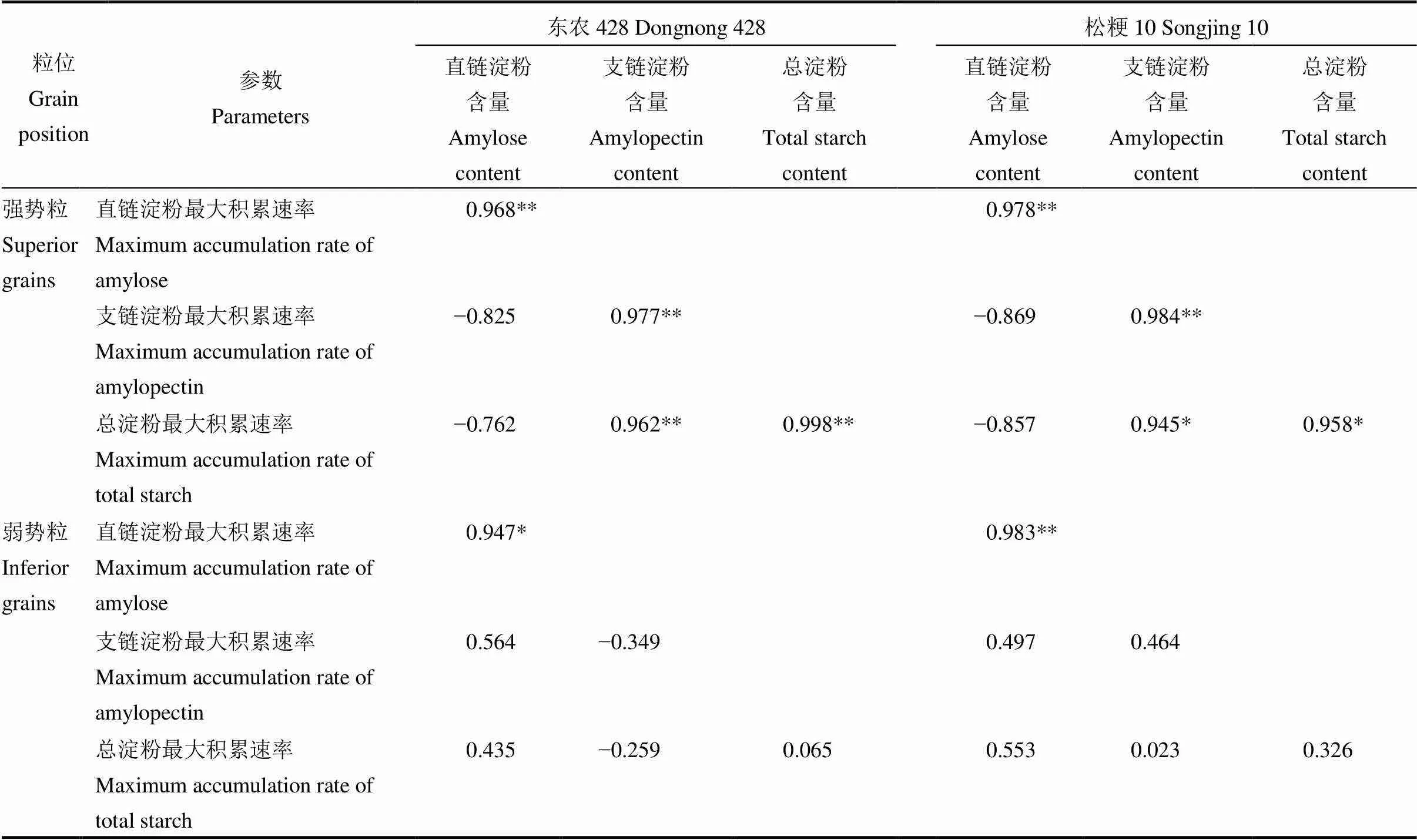
表4 淀粉含量与最大积累速率之间的相关分析
SSS峰值活性因品种耐冷性不同而存在差异,东农428强、弱势粒SSS峰值活性CRI分别为78.69%、79.56%、82.74%、94.39%和77.37%、79.68%、88.51%、92.88%;松粳10强、弱势粒SSS峰值活性CRI分别为74.88%、82.03%、87.25%、92.86%和72.26%、76.95%、86.03%、90.17%,SSS峰值活性CRI小于东农428,说明松粳10对低温抵御能力较弱,受影响大。
2.2.4 淀粉分支酶(SBE)活性
由图7可见,2个供试品种强、弱势粒 SBE活性随生育进程的推进呈先升高后下降的单峰曲线变化。与D0相比,各低温处理强、弱势粒SBE峰值活性显著下降(除东农428强、弱势粒D1处理),且随处理天数增加降幅逐渐增大,各低温处理间差异显著。D0、D1处理强、弱势粒SBE活性分别在齐穗后18、23 d最高,D5、D7处理则分别在齐穗后23、28 d活性最高,较D0推迟5 d,而松粳10低温处理3 d就出现峰值推迟现象。
SBE峰值活性因品种耐冷性不同而存在差异,东农428强、弱势粒SBE峰值活性CRI分别为86.87%、89.61%、96.17%、98.51%和82.87%、87.04%、94.02%、98.40%,受影响小;冷敏感型松粳10强、弱势粒SBE峰值活性CRI分别为74.05%、81.99%、86.13% 、93.76%和70.03%、79.69%、83.39%、92.36%,SBE峰值活性CRI小于东农428,说明松粳10对低温抵御能力较弱。
2.3 淀粉合成关键酶活性与淀粉组分积累的关系
由表4所示,强、弱势粒直链淀粉含量与直链淀粉最大积累速率呈显著或极显著正相关,强势粒支链淀粉和总淀粉含量与其对应的最大积累速率呈显著或极显著正相关,弱势粒支链淀粉和总淀粉含量与其对应的最大积累速率不相关或呈负相关。
如表5所示,齐穗后8~13 d,东农428强势粒和松粳10 强、弱势粒直链淀粉积累速率与AGPase呈显著或极显著正相关,东农428弱势粒和松粳10强、弱势粒支链淀粉积累速率与AGPase呈显著或极显著正相关,总淀粉积累速率与AGPase活性呈显著或极显著正相关。
齐穗后8~13 d、18~33 d强势粒直链淀粉积累速率与GBSS活性呈显著或极显著正相关,齐穗后8~38 d弱势粒直链淀粉积累速率与GBSS活性呈显著或极显著正相关,除齐穗后8~13 d强、弱势粒直链淀粉积累速率与AGPase、SSS活性变化呈显著或极显著正相关,齐穗后13~18 d东农428弱势粒直链淀粉积累速率与AGPase呈显著正相关、齐穗后13~18 d松粳10弱势粒直链淀粉积累速率与SSS、SBE活性变化呈显著或极显著正相关外,其余时期直链淀粉积累速率与AGPase、SSS、SBE不相关或负相关。

表5 淀粉积累速率与酶活性变化的相关分析
Q1、Q2、Q3、Q4、Q5、Q6分别表示强势粒齐穗后 8-13、13-18、18-23、23-28、28-33、33-38;d的淀粉积累速率;R表示弱势粒。
DAFH, Days after full heading.Q1, Q2, Q3, Q4, Q5, Q6represent the starch accumulation rate of superior grains at 8-13, 13-18, 18-23, 23-28, 28-33, 33-38 days after full heading; R means inferior grains.
除齐穗后8~13 d弱势粒和33~38 d松粳10弱势粒支链淀粉积累速率与SSS、SBE活性不相关,其余各生育时期支链淀粉积累速率都分别与SSS、SBE活性呈显著或极显著正相关。其中,齐穗后13~18 d强势粒和18~23 d弱势粒与SSS呈显著或极显著正相关,齐穗后18~23 d强势粒和23~28 d弱势粒与SBE呈显著或极显著正相关。说明SSS和SBE两者相互协同共同催化支链淀粉积累,但2种酶活性达到峰值的时间略有不同,SSS发挥作用的时间较 SBE 稍早,这可能是支链淀粉积累在达到最大速率后仍保持较高积累量的原因。
2.4 结实期低温胁迫对水稻产量及其构成因素的影响
由表6可见,低温对2个供试品种水稻的有效穗数和每穗粒数影响小,各处理间差异不显著,但品种间差异极显著。两品种水稻结实率、千粒重及产量在处理和品种间差异极显著。与D0相比,两品种各低温处理每穗粒数有所下降,但均未达到显著水平。就结实率和千粒重而言,除东农 428 D1处理千粒重与D0差异不显著,其余各低温处理结实率和千粒重均显著低于D0。东农 428 D1、D3、D5、D7处理的结实率较D0分别下降了1.32%、5.60%、9.65%、12.29% (2020),2.59%、9.11%、12.79%、15.13% (2021);松粳10 各低温处理下降3.75%、6.47%、12.36%、15.56%(2020)、3.84%、8.98%、13.90%、17.64% (2021)。东农 428各低温处理千粒重分别较D0下降了0.40%、4.34%、10.88%、16.28% (2020),1.44%、5.92%、9.68%、16.36% (2021);松粳10各低温处理下降5.22%、9.89%、16.34%、19.68% (2020),5.37%、12.14%、16.93%、21.35% (2021)。低温胁迫下水稻的最终产量均大幅度降低,东农 428 各低温处理的产量与对照相比降幅为8.67%、18.12%、24.80%、33.17%(2020)、7.65%、17.69%、23.24%、32.77% (2021),松粳10 降幅为 13.90%、23.07%、31.41%、39.58% (2020)、11.06%、22.80%、29.69%、38.90% (2021)。上述结果表明,结实期低温胁迫下水稻产量降低的主要原因是结实率和千粒重下降。
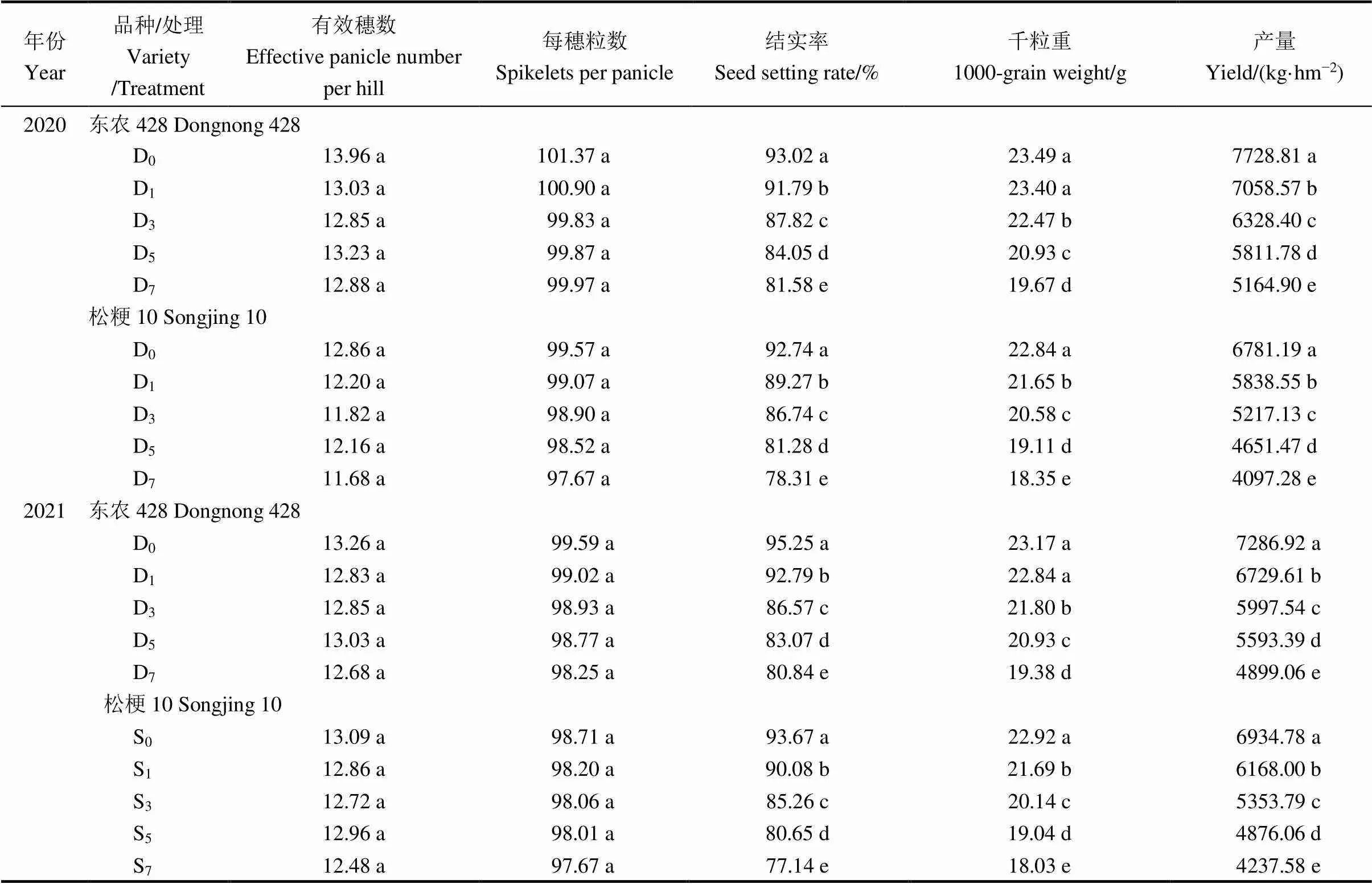
表6 结实期低温胁迫对水稻产量构成因素的影响
3 讨论
淀粉是水稻籽粒中的关键的藏物质,结实期温度与淀粉形成积累存在着密切的联系[36]。目前对低温胁迫下强、弱势粒淀粉积累差异的机理研究较多[37, 38],普遍认为结实期低温严重影响水稻籽粒淀粉积累进程,降低淀粉合成速度,引起籽粒淀粉含量下降,导致籽粒中直链淀粉与支链淀粉的比例发生改变[39]。本研究发现,随低温处理天数增加,支链淀粉和总淀粉含量降幅增大,弱势粒齐穗后8 d各低温处理和强势粒齐穗后28、33、38 d D1处理支链淀粉含量与对照差异不显著,强势粒齐穗后28~38 d D1处理总淀粉含量与对照差异不显著。分析原因可能是短时低温降低了籽粒前期的淀粉合成速率,但处于优势位置的强势粒组织形成快,代谢能力强,随着生育进程温度恢复,酶活性恢复较快,使强势粒支链淀粉和总淀粉积累增加,含量接近对照水平;而处于劣势位的弱势粒维管束小,生理结构薄弱,短时低温也可能对其灌浆造成不可逆的负面影响[40]。长时间低温胁迫对水稻组织结构及内含物造成一定程度损伤,争夺同化物的能力变差,没有充足的底物供应,后期温度回升也不能恢复灌浆,同时籽粒淀粉合成底物转化效率也因低温胁迫而受到限制[41, 42]。说明弱势粒对低温更为敏感,结实期低温胁迫对弱势粒淀粉合成影响明显,最终造成总淀粉含量下降,而强势粒在经历1 d低温胁迫情况下,随温度恢复在生育后期总淀粉含量恢复至对照水平,这是强、弱势粒对低温胁迫响应的差异所在。同时,低温胁迫下东农428强、弱势粒总淀粉含量显著高于松粳10,且与对照相比,强、弱势粒总淀粉含量降幅小于松粳10。这暗示了结实期低温胁迫下不同耐冷型品种间差异主要表现在强、弱势粒淀粉积累的差异。通过淀粉最大积累速率与淀粉含量的相关性分析可知,低温胁迫下水稻直链淀粉含量和强势粒支链淀粉、总淀粉与其最大积累速率呈极显著正相关,而弱势粒支链淀粉、总淀粉含量则与最大积累速率不相关或负相关,说明低温胁迫下强势粒淀粉含量下降主要由最大积累速率下降所引起,弱势粒支链淀粉和总淀粉含量降低很大程度上还受到最大积累速率时间推迟的影响,这与郭连安等[43]的研究结果一致。
淀粉的合成积累受到AGPase、SSS、SBE、GBSS活性变化的调控[44, 45]。低温导致淀粉合成关键酶活性不同程度下降,对淀粉合成的各个阶段影响显著[46-48]。本研究发现,低温显著降低了水稻强、弱势粒AGPase、SSS、SBE峰值酶活性,不同程度推迟了酶活性达到峰值的时间。齐穗后8~13 d AGPase活性变化与强、弱势粒总淀粉积累速率呈显著或极显著正相关,AGPase通过调控灌浆初期直链淀粉和支链淀粉积累进而决定总淀粉合成速率,为淀粉合成提供必要的原料,其活性对淀粉的积累速率有较大的影响。通过支链淀粉积累速率与酶活性变化的相关性分析可知,低温胁迫下SSS、SBE活性变化与支链淀粉最大积累速率出现的时间和积累速率一致,两者相互协同分别在生育期的不同阶段共同调控着支链淀粉积累。以上均与前人研究结果一致[49, 50]。GBSS是不同温度处理下调控籽粒直链淀粉占比的重要位点,灌浆中后期GBSS活性升高,催化直链淀粉大量合成[51, 52]。本研究发现,较长时间低温处理(D3、D5、D7)显著降低齐穗后8~23 d GBSS活性,而齐穗后28~38 d,GBSS活性显著上升,且增幅随低温处理天数的增加而增大,GBSS活性变化与直链淀粉最大积累速率变化趋势基本一致。由相关分析可知,齐穗后8~13 d、18~33 d强势粒直链淀粉积累速率与GBSS活性变化存在显著或极显著正相关,弱势粒在整个生育时期均与GBSS活性变化呈显著或极显著正相关。表明GBSS是水稻灌浆过程中对直链淀粉积累起决定性作用的关键酶,这也是长时间低温胁迫下(D3、D5、D7)直链淀粉含量在齐穗后8~23 d显著下降,齐穗后28~38 d直链淀粉含量逐渐升高并高于对照的主要原因。据此,笔者认为,这可能是水稻GBSS的一种低温响应机制,以弥补低温胁迫对淀粉合成造成的不利影响,但关于GBSS对直链淀粉在低温响应的调控机理的研究比较缺乏,值得进一步探讨。本研究结果表明,与对照相比,AGPase、SSS、SBE峰值酶活性显著下降,GBSS峰值酶活性显著上升,且随低温处理天数增加,酶活性变化幅度增大。弱势粒峰值酶活性变化幅度大于强势粒,可以推断低温胁迫对弱势粒酶活性影响较大,造成弱势粒淀粉积累时间缩短,积累速率降低,含量下降幅度增大,充实度变差,导致最终千粒重降低。因此,提高弱势粒AGPase、SSS、SBE活性有利于籽粒灌浆,增加粒重[53]。本研究还发现,在上述酶活性达到峰值之前,低温处理下东农428酶活性普遍高于松粳10(除GBSS强势粒),与对照相比,东农428强、弱势粒峰值酶活性降幅小于松粳10,这说明较高的酶活性有利于生育后期淀粉的合成积累,这可能东农428耐冷性强的原因之一。
以往研究普遍证实,在水稻在结实期遭遇低温冷害,会导致最大灌浆速率降低,延迟整个灌浆进程,进而引起结实率和千粒重的下降,最终造成减产[54-57]。本研究发现,结实期低温胁迫对水稻有效穗数和每穗粒数影响不显著,而结实率、千粒重、产量均显著降低(除东农428D1处理千粒重外),东农428结实率、千粒重、产量降幅小于松粳10,受影响小。综上,结实期低温胁迫造成水稻产量降低的原因主要包括两方面:一是淀粉合成关键酶活性受到抑制,淀粉含量减少,造成千粒重下降;二是低温会影响水稻开花授粉过程,使花药不开裂、散不出花粉难以授精;如果低温发生在花粉完全授精后,则会影响到子房体,使其不能伸长,造成空壳,这可能是水稻即使经历1 d低温也会发生结实不良,造成结实率降低,进而导致产量降低的原因[58, 59],这与本研究结果一致。因此,为减轻结实期低温对水稻造成的危害,可以在冷害发生后通过适当增施磷肥和实施灌水保温等措施来提高植株抗冷能力[60, 61]。同时,选育耐冷型品种,采用合理栽培措施,综合防治病虫草害亦是防御冷害、保证水稻优质高产的重要途经[62, 63]。
前人对不同生育时期水稻低温冷害做了大量研究,大多集中在冷水处理模拟自然冷害的方式[29, 30]。本研究是在人工气候室内对水稻进行空气低温处理,严格控制气候室内的光照、湿度、二氧化碳浓度等条件,排除处理之外的环境因素对结果的影响。由此可知,不同处理间强、弱势中淀粉含量、积累速率及淀粉合成关键酶活性的差异主要由低温所引起。此外,本研究针对某个时间段的淀粉含量的差值与酶活性差值(变化量)进行相关分析,与以往研究利用某一时间点的淀粉含量与对应的酶活性(固定值)之间分析的方法不同,并且对强、弱势粒淀粉积累及关键酶活性的动态分析较为翔实。本研究结果可对大田生产有一定理论指导,但具体应用还有待进一步验证。
[1] 周明旭. 黑龙江省水稻生产可持续发展研究[D]. 长春:吉林大学, 2014.
Zhou M X. Study on sustainable development of rice production in heilongjiang province[D].Changchun:Jilin University, 2014.(in Chinese with English abstract)
[2] Shimono H, Okada M, Kanda E, Arakawa I. Low temperature-induced sterility in rice: Evidence for the effects of temperature before panicle initiation [J]., 2006, 101(2): 221-231.
[3] 王主玉, 申双和. 水稻低温冷害研究进展[J]. 安徽农业科学, 2010, 22: 11971-11973.
Wang Z Y, Shen S H. Research progress on low temperature and chilling injury of rice[J]., 2010, 22: 11971-11973.(in Chinese with English abstract)
[4] Zhang J, Dong P, Zhang H Y, Meng C R, Zhang X J, Hou J W, Wei C Z. Low soil temperature reducing the yield of drip irrigated rice in arid area by influencing anther development and pollination[J]., 2019, 11(3): 103-114.
[5] 窦志. 灌浆期开放式增温对水稻籽粒灌浆和品质的影响及氮素粒肥的调控效应[D]. 南京: 南京农业大学, 2017.
Dou Z. Effects of Free-air during grain filling stage on grain filling and quality of rice and regulation effect of nitrogen spikelet fertilizer[D].Nanjing:Nanjing Agricultural University, 2017. (in Chinese with English abstract)
[6] 张荣萍, 马均, 王贺正, 李艳, 李旭毅, 汪仁全. 不同灌水方式对水稻籽粒灌浆特性的影响[J]. 西昌学院学报: 自然科学版, 2007, 21(4): 23-27.
Zhang R P, Ma J, Wang H Z, Li Y, Li X Y, Wang R Q. Effects of different irrigation methods on grain filling characteristics of rice[J]., 2007, 21(4): 23-27. (in Chinese with English abstract)
[7] 黄锦文, 梁义元, 梁康迳, 林文雄. 不同类型水稻籽粒灌浆的生理生化特性研究[J]. 中国生态农业学报, 2003, 11(1): 10-13.
Huang J W, Liang Y Y, Liang K J, Lin W X. Physiological and biochemical characteristics of grain filling in different types of rice[J]., 2003, 11(1): 10-13. (in Chinese with English abstract)
[8] 徐富贤, 郭晓艺, 张林, 熊洪, 朱永川, 刘茂, 周兴兵. 杂交中稻库源结构对弱势粒灌浆结实的影响[J]. 中国农业科技导报, 2013, 15(1): 96-101.
Xu F X, Guo X Y, Zhang L, Xiong H, Zhu Y C, Liu M, Zhou X B. Effects of sink-source structure of hybrid middle rice on grain filling of weak grains[J]., 2013, 15(1): 96-101. (in Chinese with English abstract)
[9] 谈桂露, 张耗, 付景, 王志琴, 刘立军, 杨建昌. 超级稻花后强、弱势粒多胺浓度变化及其与籽粒灌浆的关系[J]. 作物学报, 2009, 35(12): 2225-2233.
Tan G L, Zhang H, Fu J, Wang Z Q, Liu L J, Yang J C.Variation of polyamine concentration in strong and weak grains of super rice after flowering and its relationship with grain filling[J]., 2009, 35(12): 2225-2233. (in Chinese with English abstract)
[10] 杨建昌, 袁莉民, 唐成, 王志琴, 刘立军, 朱庆森. 结实期干湿交替灌溉对稻米品质及籽粒中一些酶活性的影响[J]. 作物学报, 2005, 31(8): 1052-1057.
Yang J C, Yuan L M, Tang C, Wang Z Q, Liu L J, Zhu Q S.Effects of alternate wet and dry irrigation on rice quality and some enzyme activities in grain[J]., 2005, 31(8): 1052-1057. (in Chinese with English abstract)
[11] Du X D, Zhao H W, Wang J G, Liu H L, Yang L, Xu J, Song J T. Changes in starch accumulation and activity of enzymes associated with starch synthesis under different nitrogen applications in Japonica rice in cold region[J]., 2012, 38(1): 159-167.
[12] Yan Z, Ding J, Song J, Humphreys G, Peng Y X, Li C Y, Zhu X K, Guo W S. Author correction: Grain yield, starch content and activities of key enzymes of waxy and non-waxy wheat (L.)[J]., 2018, 8(1): 12-16.
[13] 胡博文, 谷娇娇, 贾琰, 沙汉景, 张君颜, 黄书勤, 赵宏伟. 盐胁迫对寒地粳稻籽粒淀粉形成积累及产量的影响[J]. 华北农学报, 2019, 34(1): 119-127.
Hu B W, Gu J J, Jia Y, Sha H J, Zhang J Y, Huang S Q, Zhao H W.Effects of salt stress on grain starch formation, accumulation and yield of japonica rice in cold regions[J]., 2019, 34(1): 119-127. (in Chinese with English abstract)
[14] Maung T Z, Yoo J M, Chu S H, Kim K W, Chung I M, Park Y J. Haplotype variations and evolutionary analysis of the granule-bound starch synthase I gene in the Korean World Rice Collection[J]., 2021, 12(1708): 707237.
[15] 刘奇华, 蔡建, 李天. 水稻籽粒中的淀粉合成关键酶及其与籽粒灌浆和稻米品质的关系[J]. 植物生理学报, 2006, 42(6): 1211-1216.
Liu Q H, Cai J, Li T.Key enzymes of starch synthesis in rice grains and their relationship with grain filling and rice quality[J]., 2006, 42(6): 1211-1216. (in Chinese with English abstract)
[16] Liu J, Qian Z, Zhou L, Cao Z Z, Shi C H, Cheng F M. Influence of environmental temperature during grain filling period on granule size distribution of rice starch and its relation to gelatinization properties[J]., 2017, 76: 42-55
[17] Kato T, Shinmura D, Taniguchi A. Activities of enzymes for sucrose-starch conversion in developing endosperm of rice and their association with grain filling in extra-heavy panicle types [J]., 2007, 10(4): 442-450.
[18] 夏楠, 赵宏伟, 吕艳超, 赵振东, 邹德堂, 刘化龙, 王敬国, 贾琰.灌浆结实期冷水胁迫对寒地粳稻籽粒淀粉积累及相关酶活性的影响[J]. 中国水稻科学, 2016, 30(1): 62-74.
Xia N, Zhao H W, Lv Y C, Zhao Z D, Zou D T, Liu H L, Wang J G, Jia Y.Effects of cold water stress on grain starch accumulation and related enzyme activities of japonica rice in cold regions[J]., 2016, 30(1): 62-74. (in Chinese with English abstract)
[19] 沈鹏, 金正勋, 罗秋香, 金学泳, 孙艳丽. 水稻灌浆过程中籽粒淀粉合成关键酶活性与蒸煮食味品质的关系[J]. 中国水稻科学, 2006, 20(1): 58-64.
Shen P, Jin Z X, Luo Q X, Jin X Y, Sun Y L. The relationship between key enzyme activities of grain starch synthesis and cooking and eating quality during rice filling process[J]., 2006, 20(1): 58-64. (in Chinese with English abstract)
[20] 宋兴年. 夜间温度对水稻弱势籽粒灌浆充实的作用机制研究[D]. 郑州: 河南农业大学, 2011.
Song X N.Mechanism of nighttime temperature on grain filling and enrichment of weak grains in rice[D]. Zhengzhou:, 2011. (in Chinese with English abstract)
[21] 钟连进, 程方民. 水稻籽粒灌浆过程直链淀粉的积累及其相关酶的品种类型间差异[J]. 作物学报, 2003, 29(3): 452-456.
Zhong L J, Cheng F M. The accumulation of amylose during grain filling in rice and the differences in related enzymes among cultivars[J]., 2003, 29(3): 452-456. (in Chinese with English abstract)
[22] 王士强, 宋晓慧, 赵海红, 孙明明, 萧长亮, 顾春梅, 那永光, 解保胜, 曹立勇, 程式华. 孕穗期低温胁迫对寒地水稻产量和品质的影响[J]. 农业现代化研究, 2016, 37(3): 579-586.
Wang S Q, Song X H, Zhao H H, Sun M M, Xiao C L, Gu C M, Na Y G, Xie B S, Cao L Y, Cheng S H.Effects of low temperature stress at booting stage on yield and quality of rice in cold regions[J]., 2016, 37(3): 579-586. (in Chinese with English abstract)
[23] 王艳华. 持续低温对沈阳地区水稻的影响及品种搭配决策研究[D]. 沈阳: 沈阳农业大学, 2013.
Wang Y H.Effects of persistent low temperature on rice in Shenyang area and research on variety combination decision[D]. Shenyang:Shenyang Agricultural University, 2013. (in Chinese with English abstract)
[24] Ali I, Tang L, Dai J. Responses of grain yield and yield related parameters to post-heading low-temperature stress in Japonica rice[J]., 2021, 10(7): 1425.
[25] 付景, 徐云姬, 陈露, 袁莉民, 王志琴, 杨建昌. 超级稻花后强、弱势粒淀粉合成相关酶活性和激素含量变化及其与籽粒灌浆的关系[J].中国水稻科学, 2012, 26(3): 302-310.
Fu J, Xu Y J, Chen L, Yuan L M, Wang Z Q, Yang J C. Changes of enzyme activities and hormone contents related to starch synthesis in strong and weak grains of super rice after flowering and their relationship with grain filling[J]., 2012, 26(3): 302-310. (in Chinese with English abstract)
[26] 沈直, 唐设, 张海祥, 陈文珠, 丁艳锋, 王绍华. 灌浆期开放式增温对水稻强势粒和弱势粒淀粉代谢关键酶相关基因表达水平的影响[J]. 南京农业大学学报, 2016, 39: 906.
Shen Z, Tang S, Zhang H Y, Chen W Z, Ding Y F, Wang S H. Effects of open warming at grain filling stage on the expression levels of genes related to key enzymes in starch metabolism in superior and inferior grains of rice[J]., 2016, 39: 906. (in Chinese with English abstract)
[27] 李健陵, 霍治国, 吴丽姬, 朱庆华, 胡飞.孕穗期低温对水稻产量的影响及其生理机制[J]. 中国水稻科学, 2014, 28(3): 277-288.
Li J L, Huo Z G, Wu L J, Zhu Q H, Hu F. Effects of low temperature at booting stage on rice yield and its physiological mechanism[J]., 2014, 28(3): 277-288. (in Chinese with English abstract)
[28] Seyede S, Fallah A. Corrigendum to "temperature effect on yield and yield components of different rice cultivars in flowering stage"[J]., 2020, 2020: 1-1.
[29] 韩涛. 孕穗期冷水胁迫对水稻碳水化合物形成积累规律的影响[D]. 哈尔滨: 东北农业大学, 2014.
Han T.Booting stage cold water stress on rice carbohydrates form accumulation pattern[D].Harbin:Northeast Agricultural University, 2014. (in Chinese with English abstract)
[30] 武琦. 不同生育时期低温胁迫下寒地粳稻淀粉积累规律的研究[D]. 哈尔滨: 东北农业大学, 2013.
Wu Q.Research on starch accumulation rule in different growth stages ofrice in cold region[D]. Harbin: Northeast Agricultural University, 2013. (in Chinese with English abstract)
[31] 何照范. 谷物淀粉组份分离及测试方法评述[J]. 粮食储藏, 1985, 6: 32-38.
He Z F.Review on separation and testing methods of grain starch components [J]., 1985, 6: 32-38. (in Chinese with English abstract)
[32] 李太贵, 沈波, 陈能, 罗玉坤. Q酶在水稻籽粒垩白形成中作用的研究[J]. 作物学报, 1997, 23(3): 338-344.
Li T G, Shen B, Chen N, Luo Y K.Study on the role of Q enzyme in the formation of chalky rice grains[J]., 1997, 23(3): 338-344. (in Chinese with English abstract)
[33] Doehlert D C, Kuo T M, Felker F C. Enzymes of sucrose and hexose metabolism in developing kernels of two inbreds of maize[J]., 1988, 86(4): 1013-1019.
[34] 程方民, 蒋德安, 吴平, 石春海. 早籼稻籽粒灌浆过程中淀粉合成酶的变化及温度效应特征[J]. 作物学报, 2001, 27(2): 201-206.
Cheng F M, Jiang D A, Wu P, Shi C H. Changes and temperature effects of starch synthase during grain filling in early indica rice[J]., 2001, 27(2): 201-206. (in Chinese with English abstract)
[35] 杨志奇, 杨春刚, 汤翠凤, 郭桂珍, 余腾琼, 张俊国, 曹桂兰, 阿新祥, 徐福荣, 张三元, 戴陆园, 韩龙植. 中国粳稻地方品种孕穗期耐冷性评价及聚类分析[J]. 植物遗传资源学报, 2008, 9(4): 485-491.
Yang Z Q, Yang C G, Yang C F, Guo G Z, Yu T Q, Zhang J G, Cao G L, A X F, Xu F R, Zhang S Y, Dai L Y, Han L Z.Evaluation and cluster analysis of cold tolerance of Chinese japonica rice landraces at booting stage[J]., 2008, 9(4): 485-491. (in Chinese with English abstract)
[36] 张文倩. 昼夜高温对水稻颖花发育及籽粒结实的影响[D]. 北京: 中国农业科学院, 2019.
Zhang W Q.Effects of day and night high temperature on rice spikelet development and grain setting[D]. Beijing:Chinese Academy of Agricultural Sciences, 2019. (in Chinese with English abstract)
[37] Yang J, Peng S, Visperas R M. Grain filling pattern and cytokinin content in the grains and roots of rice plants[J]., 2000, 30(3): 261-270.
[38] 王新鹏. 孕穗期干旱胁迫对寒地粳稻碳代谢及产量形成影响的研究 [D]. 哈尔滨: 东北农业大学, 2020.
Wang X P.Effects of drought stress at booting stage on carbon metabolism and yield formation of japonica rice in cold regions[D]. Harbin:Northeast Agricultural University, 2020. (in Chinese with English abstract)
[39] Hu Y, Li L, Tian J. Effects of dynamic low temperature during the grain filling stage on starch morphological structure, physicochemical properties, and eating quality of soft japonica rice [J]., 2020, 97(2): 540-550.
[40] 李晓光, 刘海英, 金正勋, 刘洪亮, 黄星, 徐美兰. 水稻杂交后代灌浆成熟期籽粒淀粉合成关键酶和谷氨酰胺合成酶活性变化的初步研究[J]. 中国水稻科学, 2009, 4: 443-446.
Li X G, Liu H Y, Jin Z X, Liu H L, Huang X, Xu M L. Preliminary study on the changes of key enzymes in grain starch synthesis and glutamine synthase activities during grain filling and maturity of rice hybrid offspring[J]., 2009, 4: 443-446. (in Chinese with English abstract)
[41] 李银银, 陈静, 周群, 许更文, 芮梦凯, 徐心杰, 张耗. 水稻籽粒灌浆的研究进展与展望[J]. 中国稻米, 2015, 21(4): 20-24.
Li Y Y, Chen J, Zhou Q, Xu G W, Rui M K, Xu X J, Zhang H.Research progress and prospect of rice grain filling[J]., 2015, 21(4): 20-24. (in Chinese with English abstract)
[42] Weng F, Zhang W, Wu X. Impact of low-temperature, overcast and rainy weather during the reproductive growth stage on lodging resistance of rice[J]., 2017, 7(1): 9983-10004.
[43] 郭连安, 胡运高, 杨国涛, 鄢圣敏, 易军. 不同直链淀粉含量水稻籽粒淀粉积累及其相关酶的活性变化研究[J]. 云南大学学报:自然科学版, 2014, 36(6): 942-949.
Guo L A, Hu Y G, Yang G T, Yan S M, Yi J.Study on the accumulation of starch in rice grains with different amylose contents and changes in the activities of related enzymes[J]., 2014, 36(6): 942-949. (in Chinese with English abstract)
[44] 金正勋, 杨静, 钱春荣, 刘海英, 金学泳, 秋太权. 灌浆成熟期温度对水稻籽粒淀粉合成关键酶活性及品质的影响[J]. 中国水稻科学, 2005, 19(4): 377-380.
Jin Z X, Yang J, Qian C R, Liu H Y, Jin X Y, Qiu T Q.Effects of temperature at grain filling maturity on the activities and quality of key enzymes for starch synthesis in rice grains[J]., 2005, 19(4): 377-380. (in Chinese with English abstract)
[45] 王军可, 王亚梁, 陈惠哲, 向镜, 张义凯, 朱德峰, 张玉屏. 灌浆初期高温影响水稻籽粒碳氮代谢的机理[J]. 中国农业气象, 2020, 41(12): 32-42.
Wang J K, Wang Y L, Chen H Z, Xiang J, Zhang Y K, Zhu D F, Zhang Y P.Mechanism of high temperature affecting rice grain carbon and nitrogen metabolism at the early stage of grain filling[J]., 2020, 41(12): 32-42. (in Chinese with English abstract)
[46] Hao Z, Li H W, Yang L M. Post-anthesis alternate wetting and moderate soil drying enhances activities of key enzymes in sucrose-to-starch conversion in inferior spikelets of rice[J]., 2012, 63(1): 215-227.
[47] Chen L, Deng Y, Zhu H L, Hu Y X, Jiang Z G, Tang S, Wang S H, Ding Y F. The initiation of inferior grain filling is affected by sugar translocation efficiency in large panicle rice. [J]., 2019, 12(1): 75.
[48] 朱红. 抽穗后低温胁迫对水稻若干生理特性的影响[D]. 合肥: 安徽农业大学, 2015.
Zhu H.Effects of low temperature stress after heading on some physiological characteristics of rice[D]. Hefei:Anhui Agricultural University, 2015. (in Chinese with English abstract)
[49] 杨建昌, 彭少兵, 顾世梁, Visperas R M. 水稻灌浆期籽粒中3个与淀粉合成有关的酶活性变化[J]. 作物学报, 2001, 27(2): 157-164.
Yang J C, Peng S B, Gu S L, Visperas R M.Changes in the activities of three enzymes related to starch synthesis in rice grains at grain filling stage[J]., 2001, 27(2): 157-164. (in Chinese with English abstract)
[50] 赵步洪, 张文杰, 常二华, 王志琴, 杨建昌等. 水稻灌浆期籽粒中淀粉合成关键酶的活性变化及其与灌浆速率和蒸煮品质的关系[J]. 中国农业科学, 2004(8): 1123-1129.
Zhao B H, Zhang W J, Chang E H, Wang Z Q, Yang J C.Activity changes of key enzymes in starch synthesis in rice grains during grain filling and their relationship with grain filling rate and cooking quality[J]., 2004(8): 1123-1129. (in Chinese with English abstract)
[51] Dobo M, Ayres N, Walker G. Polymorphism in the GBSS gene affects amylose content in US and European rice germplasm[J]., 2010, 52(3): 450-456.
[52] Du X D, Zhao H W, Wang J G. Changes in starch accumulation and activity of enzymes associated with starch synthesis under different nitrogen applications in japonica rice in cold region[J]., 2012, 38(1): 159-167.
[53] 徐云姬. 三种禾谷类作物强、弱势粒灌浆差异机理及其调控技术[D]. 扬州: 扬州大学, 2016.
Xu Y J.Mechanisms of grain filling differences between strong and weak grains of three cereal crops and their control techniques[D]. Yangzhou:Yangzhou University, 2016. (in Chinese with English abstract)
[54] Okamura M, Arai-sanoh Y, Yoshida H. Characterization of high-yielding rice cultivars with different grain-filling properties to clarify limiting factors for improving grain yield [J]., 2018, 219, 139-147.
[55] Ahmadi A, Baker D A. The effect of water stress on the activities of key regulatory enzymes of the sucrose to starch pathway in wheat[J]., 2001, 35(1): 81-91.
[56] 朱宽宇, 展明飞, 陈静, 王志琴, 杨建昌, 赵步洪. 不同氮肥水平下结实期灌溉方式对水稻弱势粒灌浆及产量的影响[J]. 中国水稻科学, 2018, 32(2): 155-168.
Zhu K Y, Zhan M F, Chen J, Wang Z Q, Yang J C, Zhao B H. Effects of irrigation methods at fruiting stage on grain filling and yield of weak grains in rice under different nitrogen fertilizer levels[J]., 2018, 32(2): 155-168. (in Chinese with English abstract)
[57] 张诚信. 灌浆结实期低温弱光复合胁迫对水稻产量和品质的影响[D]. 扬州: 扬州大学, 2020.
Zhang C X.Effects of low temperature and low light compound stress on rice yield and quality at grain filling stage[D].Yangzhou: Yangzhou University, 2020. (in Chinese with English abstract)
[58] Tanno H, Kiuchi H, Hirayama Y, Kikuchi H. Development of a simple testing method for cool weather tolerance at the flowering stage of rice using an air conditioned room[J]., 2000, 69(1): 43-48.
[59] 曾研华, 张玉屏, 潘晓华, 朱德峰, 向镜, 陈惠哲, 张义凯. 花后低温对水稻籽粒灌浆与内源激素含量的影响[J]. 作物学报, 2016, 42(10): 1551-1559.
Zeng Y H, Zhang Y P, Pan X H, Zhu D F, Xiang J, Chen H Z, Zhang Y K. Effects of low temperature after flowering on grain filling and endogenous hormone content in rice[J]., 2016, 42(10): 1551-1559. (in Chinese with English abstract)
[60] Chu G, Wang Z, Zhang H, Yang J C, Zhang J H. Agronomic and physiological performance of rice under integrative crop management [J]., 2016, 108(1): 117-128.
[61] Xue Y, Duan H, Liu L, Xue Y, Duan H, Liu L. An improved crop management increases grain yield and nitrogen and water use efficiency in rice [J]., 2013, 53(1): 271-284.
[62] Fang M S, Song J B, Yang L M, Fan M, Shen J, Yuan L, Jiang R, Chen X, Davies W J, Zhang F. Improving crop productivity and resource use efficiency to ensure food security and environmental quality in China[J]., 2012, 63(1): 13-24.
[63] Chand J U, Abhishek B, Rintu J. Breeding approaches and genomics technologies to increase crop yield under low-temperature stress. [J]., 2017, 36(1): 1-35.
Effects of Low Temperature Stress During Grain Filling on Starch Formation and Accumulation of Superior and Inferior Grains in Rice
CHEN Hongyang, JIA Yan*, ZHAO Hongwei*, QU Zhaojun, WANG Xinpeng, DUAN Yuyang, YANG Rui, BAI Xu, WANG Changcheng
(Key Laboratory of Germplasm Enhancement, Physiology and Ecology of Food Crops in Cold Region, Ministry of Education/Northeast Agricultural University, Harbin 150030, China; Corresponding author, email:,)
【Objective】Our purposes are to illuminate the effects of low temperature stress during grain filling on the contents, accumulation rates and key enzyme activities of starch components in superior and inferior grains in rice, to clarify the regulatory effect of changes in the activities of key enzymes in starch synthesis on the rate of starch accumulation and to explore the effect of difference in granule starch formation and accumulation in superior and inferior grains on rice yield formation. 【Method】This experiment was carried out with the chill-tolerant variety Dongnong 428 and chill-sensitive variety Songjing 10 as the test materials at room temperature (daytime temperature 28℃, 14 h/night temperature 22℃, 10 h; 7 days), and low temperature (17℃, four treatments).The low temperature treatments lasted 1, 3, 5, and 7 days(abbreviated to D1, D3, D5and D7), respectively.The difference in the accumulation of starch components of superior and inferior grains and the changes in the activities of key synthesizing enzymes under low temperature stress during grain-filling period were analyzed.【Result】Compared with the control, low temperature stress during grain filling period reduced the peak activities of adenosine diphosphoglucose pyrophosphorylase (AGPase), soluble starch synthase (SSS), starch branching enzyme (SBE) and the contents of amylopectin and total starch in the superior and inferior grains of the two rice varieties, improved granule-bound starch synthase (GBSS) activity and amylose content in D3, D5, and D7treatments 28-38 days after heading. Compared with the control, the highest enzyme activity and the fastest accumulation of starch in low temperature treatments occurred at a later time, and the D7treatment had the greatest impact.The contents of amylopectin and total starch of the superior and inferior grains in D7decreased most on the 13th and 18th days after heading, and the amylose content increased most on the 28th dayafter heading.Correlation analysis showed that the amylose content of superior and inferior grains and the content of superior amylopectin and total starch are extremely significantly positively correlated with their maximum accumulation rates. The accumulation of inferior grain amylopectin and total starch is also related to the time to reach the maximum accumulation rate.The changes in the activities of AGPase, GBSS, SSS, and SBE are closely related to the accumulation rate and time of starch accumulation, and have a significant impact on the changes in starch and starch component contents.At the same time, low temperature stress during the grain filling stage significantly reduced the thousand-grain weight, seed setting rate and yield of rice, and the decrease gradually increased with the increasing low temperature treatment days.Low temperature stress during the grain filling stage has a greater impact on the changes in the key enzyme activities of starch synthesis in the inferior grains than the superior ones. The starch synthesis and accumulation of inferior grains slowed down and the content decreased, resulting in a significant decrease in thousand-grain weight and yield. 【Conclusion】In terms of varieties, the cold-tolerant Dongnong 428 has relatively high key enzyme activities of starch synthesis, and the contents of starch and its components is relatively high, so that the rice yield can be maintained at a relatively high level under low temperature stress.Therefore, the key enzyme activities of starch synthesis of granules in superior and inferior grains play a very critical role in the regulation of starch synthesis, and the changes and levels of starch components have a very important impact on yield.
rice;grain filling stage; low temperature; starch synthesis
10.16819/j.1001-7216.2022.211105
2021-11-08;
2022-02-28。
黑龙江省应用技术研究与开发计划资助项目(GA20B101);黑龙江省自然科学基金资助项目(LH2020C005)。
猜你喜欢
现代畜牧科技(2021年4期)2021-12-05 15:34:45
现代畜牧科技(2021年10期)2021-11-19 08:42:26
疯狂英语·初中天地(2020年8期)2020-09-11 07:43:38
中国化肥信息(2020年7期)2020-03-19 01:54:06
中国化肥信息(2019年7期)2019-08-26 09:46:54
中国粮油学报(2019年4期)2019-07-12 09:06:32
河北农业科学(2018年2期)2018-07-26 11:28:14
商洛学院学报(2017年2期)2017-05-17 05:19:53
中国塑料(2016年2期)2016-06-15 20:29:57
福利中国(2015年1期)2015-01-03 08:40:47
