
利用Ecopath模型评价鲢鳙放养对千岛湖生态系统的影响
2022-09-13 08:46郑一琛常剑波
生态学报 2022年16期
邓 悦,郑一琛,常剑波,3,*
1 武汉大学水利水电学院, 武汉 430072 2 武汉大学水生态研究所, 武汉 430072 3 武汉大学水资源与水电工程科学国家重点实验室, 武汉 430072
千岛湖是钱塘江的重要水源,对保障钱塘江中下游的水环境质量以及周边上千万人口的饮用水安全起着非常重要的作用。然而随着地区经济的加速发展,千岛湖的污染负荷迅速增长,于1998年和1999年爆发了两次大面积的水华。自此后千岛湖实施了以鲢、鱅放养和凶猛鱼类控制为主要措施的“保水渔业”,目的是为通过下行效应遏制水华现象。但近年来千岛湖局部水域仍不时爆发蓝藻水华,可见生态系统在人工干预下的结构化进程尚未被真正了解。
Ecopath模型最早由Polovina在1984年提出[1],随后结合Ulanowicz的能量分析生态学理论,发展成为一种生态系统能量流动的定量分析方法[2]。它基于物质平衡原理,可以对生态系统的食物网结构、能量流动过程和流动特征等进行量化综合分析[3—4],目前已经成为水域生态系统研究的重要工具,广泛应用于生态系统的食物网结构及能流特征研究[3,5—7],生态修复和渔业的影响评价[8—13]以及区域生态系统对比分析中[14—19]。本研究依据2008—2010年间在千岛湖进行的生态和渔业资源调查数据,构建了千岛湖生态系统2010年的Ecopath模型。通过分析2004—2016年间生态系统的基本结构和总体特征变化趋势,总结在“保水渔业”的人工干预下的生态系统结构化过程,评价长期投放鲢、鳙鱼对生态系统造成的影响。本研究工作从生态学原理上分析修复措施给淡水水体带来的影响,是一次有益的探索和尝试,可为后期类似水体生态修复评价提供数据支撑。
1 材料和方法
1.1 研究区域概况
千岛湖位于浙江西部与安徽南部交界的淳安县境内,地处29°11′—30°02′N,118°34′—119°15′E,是20世纪50年代为修建新安江水电站筑坝拦蓄新安江上游而形成的淡水湖泊。千岛湖水域面积约为570 km2,东西长约60 km,南北宽约50 km,正常库容量为178.4×108m3,最深处达97 m,平均水深34 m,多年平均水温15.8℃。千岛湖是饮用水重要水源地,其渔业也十分发达,是淳安县的支柱产业。
1.2 Ecopath模型
1.2.1划分功能组
功能组是指具有共同栖息地、相似食性、相似尺寸、相似生活史特征的物种集合。为使模型更具可比性,在本研究中,除依据各生物种类的食性、个体尺寸及栖息地特点等特性外,还参考了2004年的划分标准,将千岛湖生态系统划分为17个功能组。由于千岛湖生态系统种群结构的变动,2004年、2016年和2010年的功能组组成有所不同(表1)[12,20]。
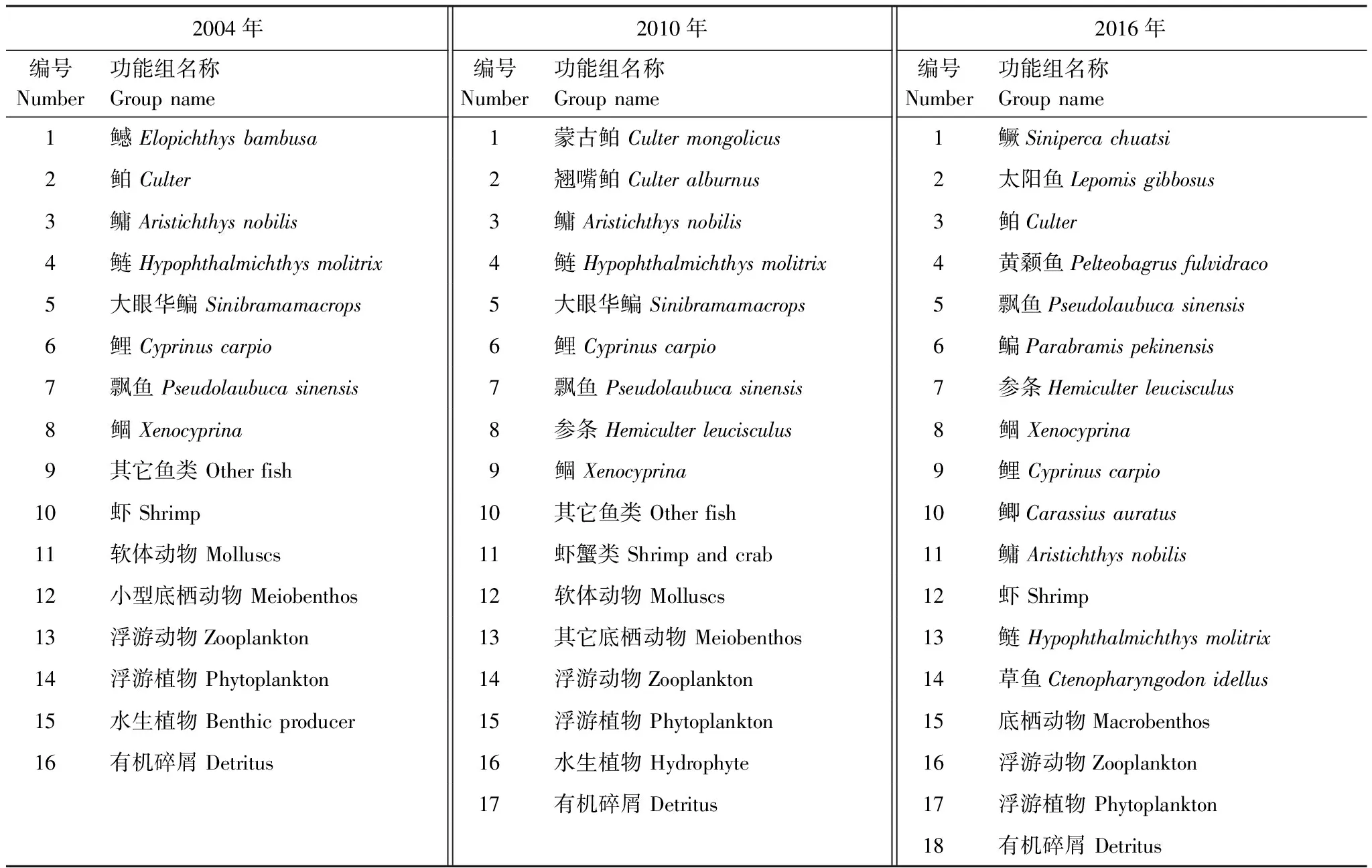
表1 千岛湖生态系统3个年份的功能组
1.2.2生态学参数
各功能组的各类生态学参数的估计如下:
(1)生物量(B)
生物量在Ecopath模型中选用生物湿重(t/km2)的能量形式来表示。部分鱼类的生物量数据由捕捞量数据计算而出,具体的计算方法为:
B=Y/F=Y/(Z-M)
(1)
式中,Y表示渔获量,F为捕食死亡率,Z为总死亡系数,Z=P/B,M为自然死亡系数,M的数值由 Pauly. D在1980年提出的经验公式估算出[21]:
logM=-0.2107-0.0824logW∞+0.6757logK+0.4627logT
(2)
logM=-0.0066-0.279logL∞+0.6543logK+0.4634logT
(3)
式中,L∞为渐进体长(cm),W∞为渐进体重(g),K为生长系数,T为该水域年平均水温(℃),取15.8℃;
研究中其它底栖生物、浮游动物、浮游植物的生物量参考在2008—2010年的实测数据[22—24]。部分功能组的生物量因为无已知的生态调查数据,故采用预设的生态营养转换效率EE算出。参照其它许多生态系统模型通行的方法[25], 将虾蟹类、软体动物、底栖藻类的EE值分别假定为 0.95、0.95和0.5。碎屑的生物量计算采用与初级生产碳相关的相关经验公式[26]。
(2)生产量/生物量(P/B)
P/B系数指年生产量和年平均生物量的比值。鱼类和虾蟹类的P/B系数参考千岛湖和竺山湖的相关文献[12,27];软体动物和其它底栖生物的P/B系数参考太湖的相关研究[28];浮游动植物以及底栖藻类的P/B系数引自千岛湖相关文献[12]。
(3)消耗量/生物量(Q/B)
Q/B系数为单位时间某种生物的消耗量与生物量的比值。研究中鱼类的Q/B由Pauly的经验公式计算[21]:
log(Q/B)=7.964-0.204logW∞-1.965T′+0.083A+0.532h+0.398d
(4)
T′=1000/(T+273.15)
(5)
A=H2/a
(6)
式中,W∞是渐进体重(g),T为该水域的表层水温(℃),A为尾鳍形状参数(一般鱼类取1.32),h为布尔型变量(草食鱼类取1,其它类取0),d为布尔型变量(碎屑食性的鱼类取1,其它类取为0),H为鱼类的尾鳍高度(cm),a则为尾鳍面积(cm2)。所需要的参数数据通过查询渔业数据库网站(http://www.fishbase.org),并参考相关的文献[12,28—29]。
虾类、软体动物、其它底栖动物和浮游动物的Q/B由Q/B=(P/B)/(P/Q) 间接计算而来, 它们的P/Q的取值参照一些公认的资料[30],分别为0.075、0.125、0.02和0.05。
(4)捕捞量和迁移量产出(EX)
参考千岛湖鱼类资源数据中做的渔业捕捞系统调查得到,统计中额外考虑鲢、鳙鱼每年的投放量,作迁入量计。
(5)生态营养效率(EE)
虾、软体动物、底栖藻类的EE值分别假定为 0.95、0.95和0.5[25],部分鱼类功能组参考相关文献来取值[12,31]。
(6)食物矩阵(DC)
鱼类的食性组成参考千岛湖和太湖的相关文献[27,31];对包含几种动物的功能组,食物组成则根据权重算出。浮游动物、软体动物、水蚯蚓和虾蟹类的食性参照目前的食性研究结果[26,32]。
1.2.3模型调试
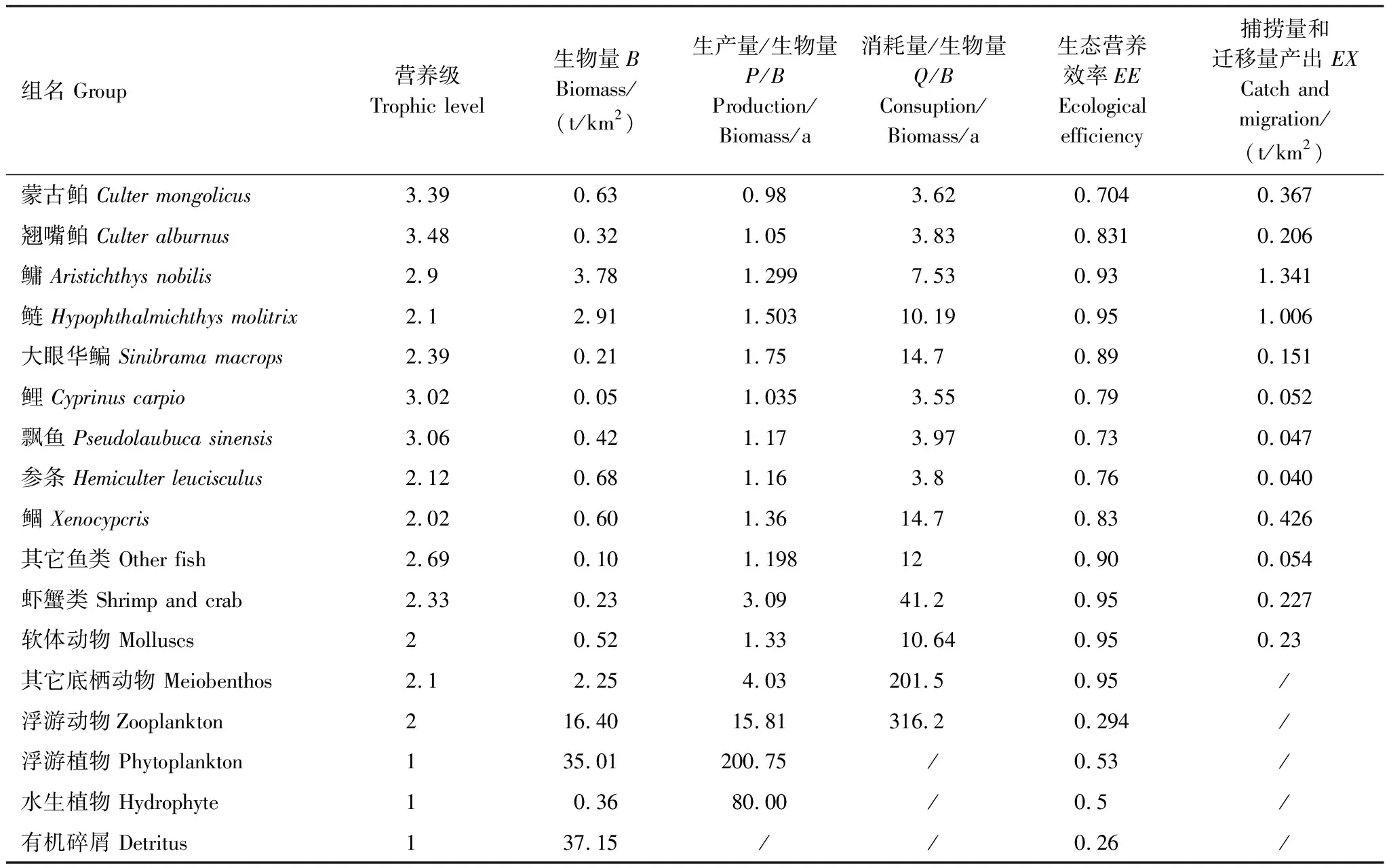
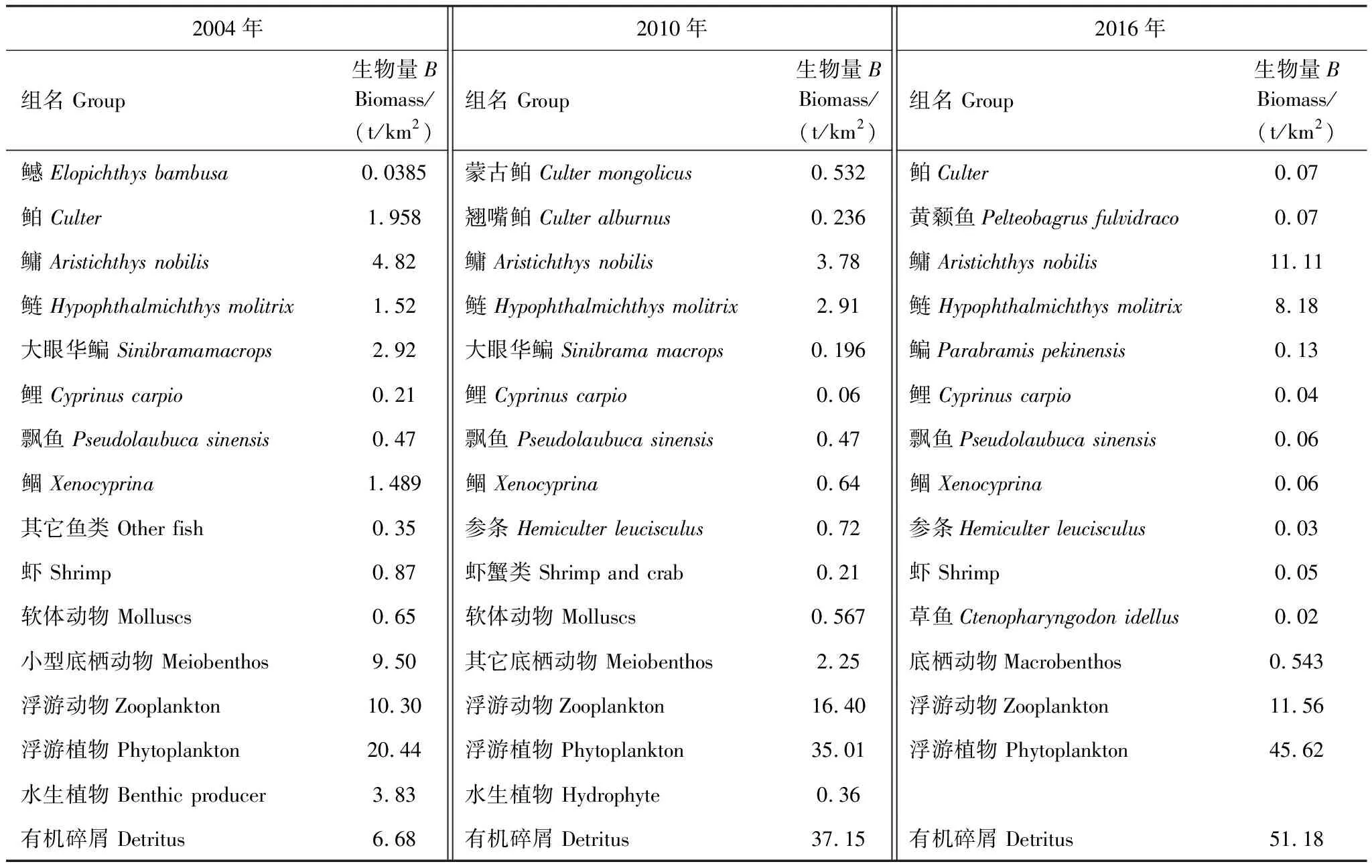
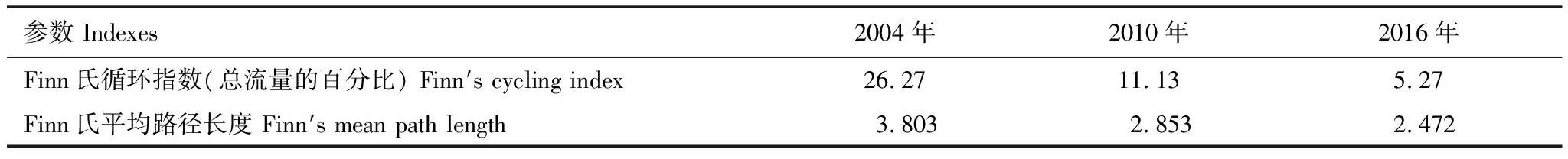
基本参数输入模型后,需要保证各功能组的生态营养转换效率EE值都要调整到在0和1之间。因为超过1的EE值是不正常的,任一个物种被捕食和被捕捞的量不可能大于其自身的能量产出[33—34]。这时需要通过调节部分参数,使模型中各个功能组的输入和输出达到平衡。模型在首次运行时出现有的功能组的EE值大于1,通过调整P/B或者DC等来使0 参数输入模型后,进行模型的平衡与计算,输出千岛湖生态系统Ecopath模型的运行结果如表2所示。 将其中一些重要的参数与刘其根[12]构建的2004年模型结果以及于佳等[20]的2016年模型结果进行比较。生物量列表如下表3,由于千岛湖生态系统种群结构的变动以及划分功能组的不同,此处选取一些具有代表性的功能组作对比。 各功能组生物量变化可以经由表3看出。鲢、鳙属于人工放养鱼类,其生物量大小主要取决于放养和捕捞情况。其它鱼类里,鳡鱼的生物量经由1999和2000年的除鳡行动在2004年已经大幅减少,到2010年几乎完全消失。鲌鱼、飘鱼、参条和鲴鱼等鱼在2004年的生物量相较于保水渔业实施前有短期的提升[12]。但经过六年后却都大幅度减少,到2016年仍呈现继续减少的趋势。除此之外,底栖动物的生物量也大幅减少,浮游植物和有机碎屑的生物量却成倍增长。 表2 千岛湖2010年生态系统模型功能组参数和输出结果 表3 3个年份的各功能组生物量对照 为了更直观的表示出食物网关系,将不同功能组的营养流合并为数个营养级,称为整合营养级[35]。2010年的能量流动过程可以形象的用下图1的林氏锥分析法图来表示。在忽略了生物量和生产量等都非常低的营养级之后,千岛湖生态系统2004年、2010年和2016年的整合营养级为4个(表4)。其中低营养级的能流在整体系统中占比较大,越往顶级越小,成典型的金字塔型。2004年营养级Ⅰ、Ⅱ的总流量分别为4214 t km-2a-1和2285 t km-2a-1,占全部营养级能流量的64.5%和35%;2010年营养级Ⅰ、Ⅱ的总流量分别为7057 t km-2a-1和3737 t km-2a-1,占总能流量的65.0%和34.4%;2016年营养级Ⅰ、Ⅱ的总流量分别为9991 t km-2a-1和3683 t km-2a-1,占总能流量的72.8%和26.9%。在流向碎屑的总量上,2004年为3483 t km-2a-1,2010年为5921 t km-2a-1,2016年为8826 t km-2a-1,分别占总能流的53.3%、54.6%和64.3%。 图1 千岛湖2010年生态系统各营养级之间的能量流动/(t km-2 a-1)Fig.1 Energy flows between trophic levels of Qiandao Lake ecosystem in 2010 表4 千岛湖3个年份生态系统整合营养级能流分布 从表5可见,2010年的生态系统在生产力和营养物总流通量上较2004年有较大程度的增加,2016年则又在2010年的基础上提升明显。2004年系统的总生产力为4440.0 t km-2a-1,到2010年升高为7341.1 t km-2a-1,增加了65 %,2016年则为10244 t km-2a-1,为原先的2.3倍;总净初生产力为初级生产者产出的产量总和,2010年和2016年分别为7059 t km-2a-1和9991 t km-2a-1,增幅分别为67%和137%;在总消耗和总呼吸值上,2004年到2010年和2016年的数值逐渐变大;系统的总流通量通常可以表征生态系统的规模,2004年的值为16041.0 t km-2a-1,2010年和2016年则分别为20112.6 t km-2a-1和24698.2 t km-2a-1,呈现持续增大的趋势。 表5 千岛湖生态系统3个年份的总体特征 表5中还陈列出一些可以反映生态系统成熟度特征的参数。系统总初级生产量/总呼吸量(TPP/TR)在三年间的值分别为3.73、4.35和6.51,远大于1且逐渐升高。连接指数是该生态系统食物网的实际连接数与可能连接数的比率;系统杂食指数被定义为所有消费者的平均杂食指数[34],这两个指数可以在一定程度上反映出生态系统内部食物网联系的复杂程度。千岛湖生态系统3个年份的连接指数(CI)和杂食性指数(SOI)相差不大,在杂食性指数上略有增大的趋势。 Finn氏循环指数和 Finn氏平均路径长度也与系统成熟度、复原力和稳定性密切相关[34]。如表6所示,千岛湖生态系统2004年、2010年和2016年的Finn氏循环指数(FCI)分别为26.27%、11.13%和5.27%,Finn氏平均路径长度(FMPL)为3.803、2.853和2.472,两项指标的数值都大幅减小。 表6 千岛湖生态系统3个年份的网络分析指数 从上述数据中可以总结出2010年千岛湖生态系统在营养级、能流状况等方面的特征。2010年的千岛湖中除人工投放的鲢、鳙鱼之外,大多数鱼类和底栖动物的生物量都较低,浮游植物和有机碎屑的生物量却较大。整合营养级的营养能流分布成典型的金字塔型,且高、低营养级的能量转化效率相差很大,总体来说低于林德曼提出的生态金字塔1/10效率,以及Christensen等[36]在1993年总结多个生态系统模型提出的生态系统平衡传输效率(9.2%)。高强度捕捞干扰和能量传递阻塞的现象较为明显。 生态系统的成熟度是相对于系统幼态化的一个概念,一个成熟度高的生态系统会更加稳定、更能抵抗外部冲击。Ecopath模型从Odum提出的24个用以表征生态系统结构和功能特征的指标中提取了一些比较重要的参数进行量化表示[37]。如系统总初级生产量/总呼吸量在系统的早期发展阶段会大于1,因为生产量会超过呼吸作用进行原始积累,越成熟的系统中,该比率应该越接近1[34]。千岛湖生态系统2010年的TPP/TR为4.35,大于其在2004年的值,与1999年和2000年的值(分别为2.07和1.99)相比也更大[38],反映出其成熟度的降低。与国内外其它水库相比,2010年千岛湖的TPP/TR高于印度的Wyra水库(其在实施渔业管理措施后降低至1.053)[39]和三峡水库的1.899[40],低于巴西Itaipu水库的6.3[41],处于不够成熟的发展阶段。除此之外,千岛湖生态系统的CI和SOI值分别为0.219和0.100,相比于三峡水库(0.371和0.205)[40],太湖(0.21和0.04)[28]和印度的Bakreswar水库(0.18和0.11)[42]等湖泊,表现出其在食物网结构和饮食组成的多样性方面仍缺乏复杂性。同时,2010年生态系统的FCI和FMPL分别为10.62%和2.843,与同类型其它水库相比较低。FCI代表一个生态系统的循环量,与整个系统的稳定性、恢复力和成熟度相关。它的值较高说明一个系统更加稳定和成熟[37],较低则表示生态系统较为脆弱,对营养输入的改变比较敏感[43]。模型计算出的浮游植物的高生物量在一定程度上证实了这一统计结果。由此可见,千岛湖2010年的生态系统稳定性较差,比较容易受到外界因素的干扰或破坏。 本研究通过Ecopath模型的对比分析,探明了千岛湖生态系统自实施了保水渔业后,在营养级状况、总体特征上的发展演变趋势。千岛湖渔获物的平均营养级呈现一直降低的趋势,低营养级渔获物数量增加,这种现象出现的原因可能是:自实施非经典生物操纵以来,顶级捕食者被大量清除,营养级较低的鲢、鳙鱼则被大量补充;且随着系统的发展,顶级消费者的群落组成和生物量都发生了明显的变化,特别是一些营养级较高的肉食性鱼类如鲌鱼生物量的减少,可能也是使得平均捕捞营养级下降的原因。从整合营养级来看,从2004年至2016年千岛湖系统低营养级占比在逐渐扩大,营养流中流向碎屑的部分也越来越大。营养级Ⅰ、Ⅱ的能量利用都不够充分,近年来能量传递阻塞的现象更加明显。且在持续性的人为干预下,系统内的鱼群种类结构和生物量均发生较大变动。鳡鱼被大量清除后,到2010年已经近乎完全消失。在保水渔业实施后的短期内,可能由于顶端捕食压力的减少和饵料的增加等原因,2004年的鲌类、鲴类、大眼华鳊、参条等鱼的生物量较2000年之前有一定上升[12]。但长时间的人工放养和捕捞使千岛湖的渔业资源结构发生变化, 食物网趋于简单,发展至10年之后,除了主要由人工放养控制的鲢、鳙鱼之外,鱼类资源量整体下降。然而,浮游植物和碎屑生物量却有比较明显的增加,这一方面可能是鱼类生物量下降的原因,另一方面是由于周围面源污染和水产养殖的剩余饵料在深水湖泊中的累积。 在能流特征上,系统发展后期和保水渔业实施第一年前后差别较大。刘其根等[38]在分析2000年的生态系统时发现,相比于1999年,系统的规模减小,但成熟度和稳定程度均有提升,蓝藻水华等也得到了一定程度的克制。但不可忽视的是,顶级捕食者和较高营养级鱼类生物量的变动势必会使整个系统的能流格局发生改变。TPP/TR的值逐渐增大,说明系统处于不成熟的发展阶段,且有成熟度下降的趋势。Finn氏循环指数(FCI)和Finn氏平均路径长度(FMPL)每过六年就几乎削减一半,表明在长期的鲢、鳙鱼放养累积下,生态系统的营养交互关系减弱,再循环率较低,复原力降低。系统退化到不成熟的拥有较高生产力的发育期。 综合上述指标分析,保水渔业实施初期系统规模瞬减,随后又自我调控,不断发展积蓄能量。但可能由于系统营养结构和外源输入的变化,初级生产者产出的能量未能得到有效的利用,很大一部分能量流向碎屑,鱼类资源量整体下降,可能会造成湖体营养物质过剩,影响生态系统的健康稳定发展。在保水渔业等修复工作实施后的十几年间,生态系统的营养级结构发生了一定程度的恶化。千岛湖作为一个由水库发展而来的淡水水体,捕食与被捕食的关系本身就较为简单,生态系统更容易被外界的扰动影响,故而应更加注意控制人类活动和人为手段。 “保水渔业”采用非经典生物操纵的思路,围网放养滤食性的鲢鱼和鳙鱼,直接摄食浮游植物,以期减少蓝藻水华。但鲢、鳙鱼主要抑制的是枝角类和大型浮游植物,间接促进桡足类和小型藻类种群增长,浮游植物的丰度增加。故而可能会出现蓝藻水华得到抑制,但湖域富营养化程度加重的情况。低营养级再通过上行效应影响系统中的其它组分,系统整体的稳定性降低。同时,浮游生物食性的鱼类还会加快氮、磷的循环速率,扩大初级生产力。上述分析也与文章第二部分中浮游植物生物量的逐渐上升的趋势,以及初级生产力规模的逐渐扩大相对应。 诸多国内外学者也曾探索生物治理措施对水生态系统造成的影响,研究滤食性鱼类对蓝藻水华和富营养化进程的作用。在不同水域、不同实验条件和不同研究方案下得出的结论不尽相同。王银平等通过原位培养滤食性鱼类鲢、鳙摄食微囊藻后的排泄物,利用叶绿素荧光技术研究鲢、鳙滤食对藻类光能及生长活性的影响,结果证明鲢、鳙单次滤食未对微囊藻造成生理上的致命损伤,反而可能会藻类超补偿生长[44]。而美国学者Mueller等[45]在斑点叉尾鮰养殖池塘中的实验,以及一些国内学者在东湖[46]、滆湖[47]的实验则说明了鲢、鳙控藻的可行性。因此,需要在长期、规范的实验下明确放养鱼类和系统其它组分之间的联系,准确把握不同实验结果背后的生态学原理,才能更好地验证非经典生物操纵理论的有效性。 鲢鳙鱼放养的生态修复方式使千岛湖的食物网结构发生了较大的变动。对淡水生态系统进行生态修复工作后,系统的结构化是逐渐推进的。人为的修复工作不止会影响直接作用的某几个物种,还会通过系统内部的联系传递到其它部分,从而影响整体。放养鲢鳙鱼可能会使蓝藻“水华”在短期内得到抑制,但经过一段时间的发展后,系统可能会呈现鱼类资源量下降、能量转换效率降低、成熟度和稳定度下降的状态。因此,在进行某项修复工作之前,需要从生态学原理上系统地考虑问题,从避免过多的人为扰动出发,尽量还原生态系统本身的恢复力。2 结果与分析
2.1 生物量和营养级结构

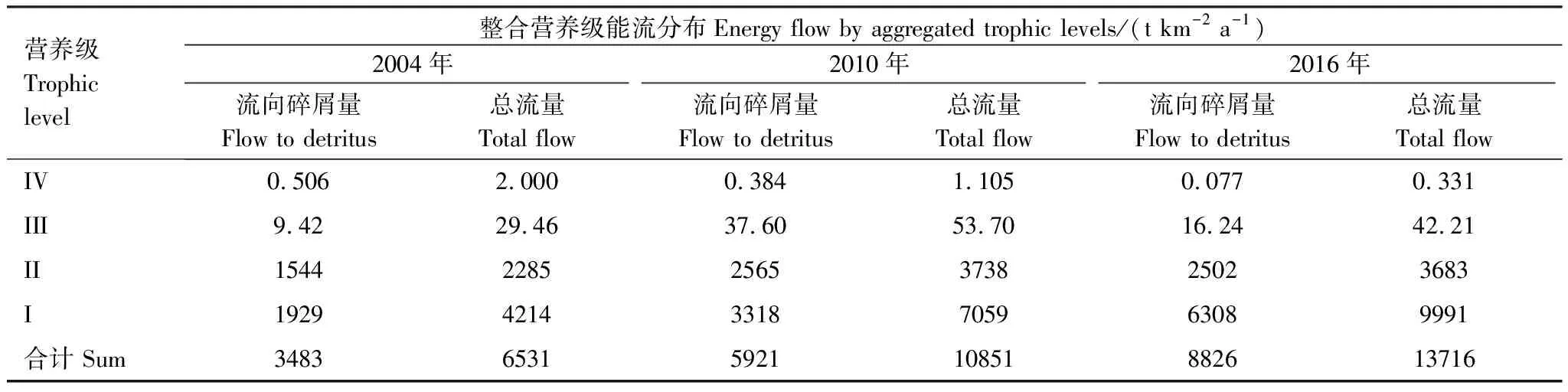
2.2 系统能量流动特征

3 讨论
3.1 生态系统总体特征
3.2 生态系统变化趋势
3.3 鲢鳙鱼放养影响
4 结论
猜你喜欢
人文天下(2022年7期)2022-09-15
今日农业(2020年19期)2020-12-14
湖北农业科学(2019年24期)2019-02-06
——千岛湖站
法人(2019年1期)2019-01-10
中国调味品(2018年9期)2018-09-15
中国自行车(2017年1期)2017-04-16
垂钓(2016年9期)2016-10-31
垂钓(2016年8期)2016-09-09
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
