
冷暴露对小鼠回肠机械屏障的影响及其机制*
2022-09-08 09:21:46刘萌萌张紫薇袁建彬郭景茹
中国应用生理学杂志 2022年3期
刘萌萌,张 ,张紫薇,袁建彬,郭景茹
(黑龙江八一农垦大学,大庆 163319)
北方寒冷天气是多种疾病的诱因,机体的免疫系统、消化系统等都易受到寒冷环境的影响[1,2]。有研究表明,冷暴露能够导致机体肠道菌群破坏、免疫细胞数量下降、生长速度减慢、生产性能和饲料转化率降低,使动物感染疾病的几率大大增加,甚至造成新生幼崽死亡,母畜流产等,进而降低动物产品的产出,制约我国北方畜牧业的经济发展[3-5]。
肠道为机体主要消化吸收的部位,极易受到外部环境的变化致肠道疾病发生。有研究表明,慢性冷暴露增加大鼠的肥大细胞数量,引起大鼠小肠上皮细胞增殖速率受损,从而促进炎症性肠炎的发生[6]。回肠具有消化、排空等功能,是动物吸取营养物质的主要部位。据报道,长期处于高温或低温的环境会破坏机体回肠微生物的多样性[7],影响机体生长发育、消化吸收、营养免疫等功能。肠道上皮屏障能够隔离肠腔与血流,可分为机械屏障、免疫屏障、生物屏障和化学屏障四种类型[8]。其中机械屏障则是肠道复杂功能网络的基础,它是由紧密连接和肠黏膜上皮细胞组成。肠上皮细胞间的连接是一种复杂的分子结构,被称为紧密连接,这种紧密连接能够保持上皮细胞和内皮细胞结构的完整性,形成一种致密的黏膜屏障来维护肠道内环境稳态[9]。紧密连接包括闭合蛋白(Claudins)、咬合蛋白(Occludin)、带状闭合蛋白(zonula occludens,ZOs)等,其中Claudin-1和Occludin是构成紧密连接的重要组成部分,ZO-1是紧密连接中跨膜蛋白的主要成分。紧密连接相关蛋白互相连接形成致密的复合物,以保证细胞与细胞之间的紧密结合[10]。以往研究发现,外界温度骤降能够减弱紧密连接功能,增加小鼠肠道通透性[11],并指出完整的紧密结构对于机体健康的重要性。此外机体的免疫系统被激活后,免疫细胞活化,炎症细胞因子释放增加,激活相关通路,也会导致上皮细胞的紧密连接受到破坏,增加通透性,病原体和细菌趁机侵入肠道,增加肠道患病的几率[12]。另有研究发现,当肠道受到抗原刺激时会使免疫防御机制遭到破坏,肠道微生物失调而引起炎症性相关疾病的发生[13]。
据此,本研究旨在探讨冷暴露对小鼠回肠屏障功能损伤的作用及NF-κB对其调控机制,为机体肠道健康及应激性肠病的靶向治疗提供基础数据。
1 材料及方法
1.1 建立小鼠冷暴露模型
将20只5周龄雄性C57BL/6小鼠(购于北京维通利华实验动物技术有限公司)随机分为两组,即对照组和冷暴露组,每组10只。试验前于室温气候室内预饲一周,保证自由饮食。预饲结束后将对照组小鼠置于恒温气候室内,饲养保持温度(24±2)℃,湿度40%。冷暴露组小鼠每天转移至温度为(4±2)℃湿度为40%的恒温气候室中3 h,连续冷暴露三周。冷暴露周期结束后注射戊巴比妥麻醉处死,沿腹部中线剪开,距离盲肠前6 cm剪断,采集回肠组织,用生理盐水反复漂洗至干净,滤纸吸干多余液体后,部分冻存于-80℃冰箱内以备用后续实验,另取部分回肠组织置于4%多聚甲醛溶液中固定。
1.2 试剂
BCA蛋白质分析试剂盒,货号:P0012S购于碧云天生物技术公司;抗体:p-p65(AF2006,1∶1 000)购于:Affinity Biosciences有限公司,Claudin-1(13050-1-AP,Proteintech,1∶5 000)、Occlundin(27260-1-AP,Proteintech,1∶3 000)、ZO-1(21773-1-AP,Proteintech,1∶7 000)、IL-1β(26048-1-AP,Proteintech,1∶800)、IL-6(66146-1-lg,Proteintech,1∶1 000)、p65(66535-1-Ig,1∶1 000)、HRP-conjugated Affinipure Goat Anti-Mouse IgG(H+L)(SA00001-1,1∶8 000)和HRP-conjugated Affinipure Goat Anti-Rabbit IgG(H+L)(SA00001-2,1∶8 000)、CoraLite 488-Conjugated ZO-1 Polyclonal Antibody(CL488-21773,IF:1∶50~1∶500)均购于武汉三鹰生物技术有限公司。
1.3 小鼠回肠组织石蜡切片
取2~3 mm厚的小鼠回肠组织,脱水后放入4%的多聚甲醛后进行固定,随后按照常规的切片制作步骤进行相应处理。然后将切片贴于已经处理后的载玻片上,烤片经一昼夜后保存,次日应用HE染色和Masson染料染色后,光学显微镜观察其形态学变化。
1.4 免疫荧光染色
制作回肠组织切片,使用PBS冲洗切片4次,每次3 min,冲洗结束后滴加10%正常山羊血清37℃下封闭45 min。封闭结束后,冲洗干净分别加入按照说明书稀释的胞质紧密粘连蛋白1抗体(zonula occludens-1,ZO-1),放入湿盒中37℃下孵育1 h,置于4℃下过夜。孵育结束后,使用PBS冲洗3次,每次5 min,然后避光加入相应的荧光二抗(1∶200),37℃下孵育45 min,孵育结束后,吸弃二抗,不再清洗,使用抗荧光淬灭剂(4',6-diamidino-2-phenylindole,DAPI)在避光条件下封片,置于荧光显微镜下观察,保存图像。
1.5 免疫蛋白印迹
蛋白样品的制备及浓度的测定:取5周龄C57BL/6对照组和冷暴露组的雄性小鼠的回肠组织,制备所需要的蛋白样品,高温变性后冻存于-20℃。将提前制备好SDS-PAGE胶置于电泳液中,按照一定顺序在梳孔加入蛋白样品后进行电泳,电泳结束后进行电转,室温封闭PVDF膜1 h,使用TBST漂洗,依次加入相应抗体,置于4℃摇床过夜。一抗孵育结束后使用TBST进行漂洗,然后使用二抗室温孵育1 h,再次漂洗干净。使用ECL化学发光试剂将带有目的蛋白的PVDF膜进行显影。应用lmage Lab软件,检测紧密连接蛋白、NF-κB蛋白、炎症细胞因子蛋白并分析各条带灰度值,以目的蛋白条带灰度值与内参蛋白条带灰度值的比值作为蛋白表达相对水平。
1.6 统计学处理
2 结果
2.1 冷暴露对小鼠回肠组织的影响
HE染色发现,对照组小鼠回肠绒毛结构完整,排列整齐。与对照组相比,冷暴露组小鼠回肠绒毛长度变短,部分绒毛缺失,隐窝深度增加,环形肌层变薄,出现大量炎症浸润现象(图1)。Masson染色结果显示:与对照组相比,冷暴露组胶原纤维沉积增加,出现轻微纤维化(图2)。免疫荧光检测可见,对照组ZO-1主要集中分布于绒毛紧密连接处,分布较为均匀、紧密;与对照组相比,冷暴露组小鼠回肠组织ZO-1荧光强度明显减弱,分布松散,失去紧密结构(图3)。

Fig.1 Effects of cold exposure on mice ileum(HE×400)
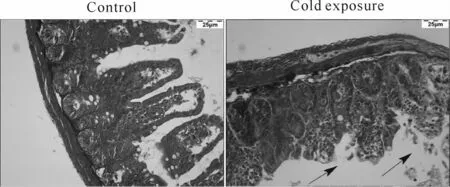
Fig.2 Effects of cold exposure on collagen fibre in mice ileum(Masson×400)

Fig.3 Effects of cold exposure on tight junction of mice ileum determined by immunofluorescence method(Scale bar=25 μm)
2.2 冷暴露对小鼠回肠组织紧密连接相关蛋白表达的影响
Western blot检测结果表明,与对照组相比,冷暴露组小鼠回肠组织Claudin-1、Occludin和ZO-1蛋白表达水平均显著下调(P<0.01,P<0.05,图4,表1)。
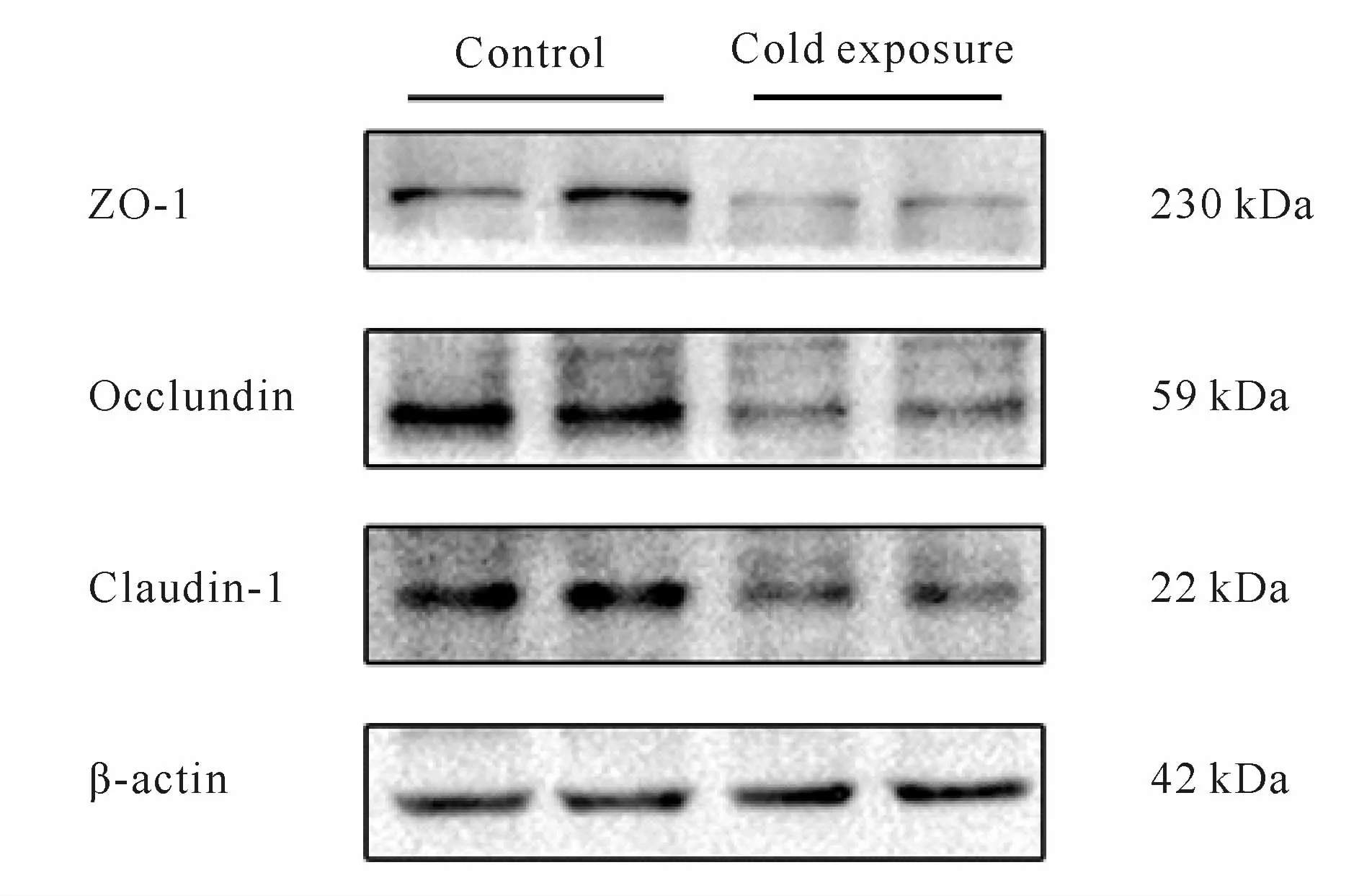
Fig.4 The protein expressionsof claudin-1,occludin and ZO-1 detected by Western blot(n=10)
Tab.1 The protein expressions levels of claudin-1,occludin,ZO-1 and p-p65 in different groups(±s,n=10)

Tab.1 The protein expressions levels of claudin-1,occludin,ZO-1 and p-p65 in different groups(±s,n=10)
*P<0.05,**P<0.01 vs control
Croup Claudin-1 Occludin ZO-1 p-p65 Control 2.35±0.28 0.58±0.21 0.69±0.17 2.95±1.18 Cold exposure 1.63±0.15**0.31±0.12*0.49±0.09*5.77±1.91*
2.3 冷暴露对小鼠回肠组织NF-κB通路相关蛋白表达的影响
Western blot检测结果表明,与对照组相比,冷暴露组小鼠回肠中NF-κB通路的相关蛋白p-p65蛋白表达显著上调(P<0.05,图5,表1)。
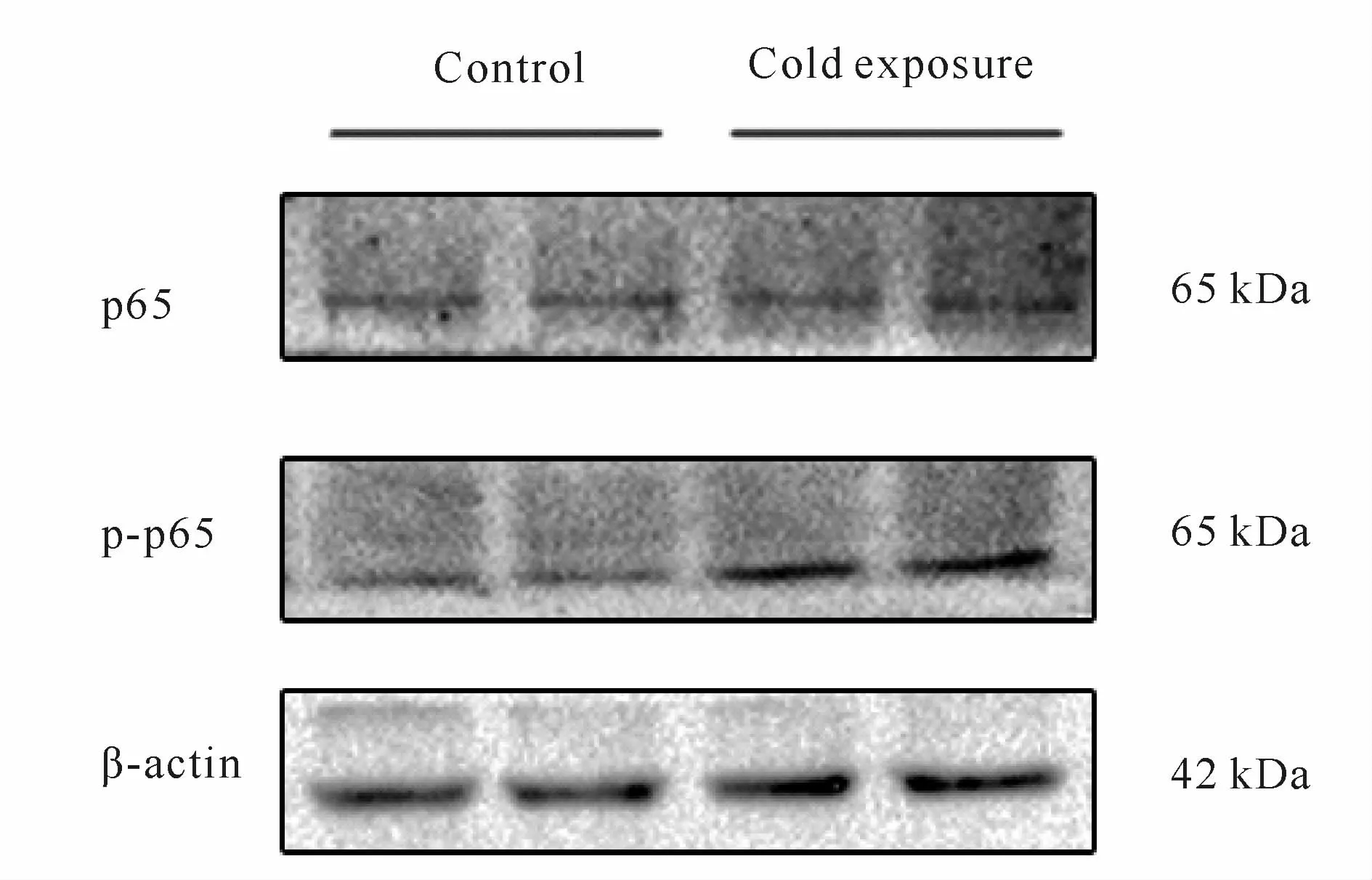
Fig.5 The protein expressions of p65 and p-p65 detected by Western blot(n=10)
2.4 冷暴露对小鼠回肠组织炎症细胞因子蛋白表达的影响
Western blot检测结果表明,与对照组相比,冷暴露组小鼠回肠组织炎性细胞因子白细胞介素-1β(interleukin-1β,IL-1β)和IL-6(interleukin-6,IL-6)蛋白表达水平均显著上调(P<0.05,P<0.01,图6,表2)。
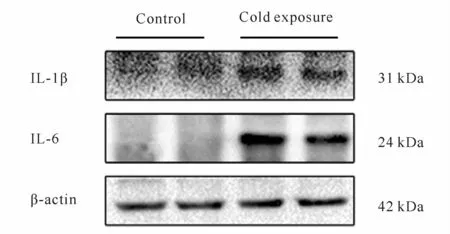
Fig.6 The protein expressions of IL-1βand IL-6 detected by Western blot(n=10)
Tab.2 The levels of IL-1βand IL-6 in different groups(±s,n=10)
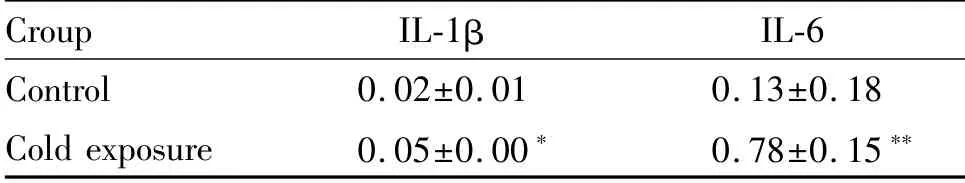
Tab.2 The levels of IL-1βand IL-6 in different groups(±s,n=10)
*P<0.05,**P<0.01 vs control
Croup IL-1β IL-6 Control 0.02±0.01 0.13±0.18 Cold exposure 0.05±0.00* 0.78±0.15**
3 讨论
近年来越来越多的研究表明,动物暴露在寒冷的环境时,可能会导致机体的体温调节发生障碍,中枢系统受损,严重可影响动物的生命安全[14]。肠道是消化管中最长的一段,功能尤为重要。回肠是消化和吸收的主要场所之一,紧密连接功能作为屏障的重要组成部分,当其功能被破坏时就会对肠道的健康造成负面影响。一旦肠屏障功能受损,肠道通透性升高,就会增加肠内细菌与毒素入血的概率,严重者或可导致肠源性脓毒血症,继发全身炎症。在紧密连接蛋白受到抑制时易产生腹腔疾病,在早晚期的肠道肿瘤中也发现几种紧密连接蛋白表达缺陷[15]。
本研究发现,冷暴露能够造成小鼠回肠结构损伤,与以往相关报道的结果相一致[16],均表明肠道绒毛结构完整性及机械物理屏障在肠道疾病的发生发展中起到不可或缺的作用。
紧密连接的相邻细胞膜间形成的屏障层,是肠道机械屏障主要的组成部分之一。动物肠道中有大量的菌群和致病因子,健康肠道的肠黏膜屏障可以有效阻止大量抗原物质的侵害,从而保证肠道的正常功能及其内环境的稳态。为适应复杂多变的肠道环境,紧密连接蛋白呈现高度动态变化,其完整性也与多种肠道疾病密切相关。以往研究发现,紧密连接蛋白的表达变化直接与小肠上皮的屏障功能和完整性直接相关[17]。本实验在冷暴露的条件下,小鼠回肠的紧密连接蛋白显著下降,致使肠道屏障受损。发生严重烫伤时,在创伤应激条件下紧密连接被破坏,肠黏膜屏障功能被削弱,通透性升高,从而导致细菌移位和内毒素血症[18]。
核因子κB(nuclear factor kappa-B,NF-κB)能够参与体内的应激反应、免疫应答与炎症反应[19]。细胞正常状态下,NF-κB主要以二聚体形式存在,其中大部分二聚体与抑制蛋白(inhibitor of NF-κB,IκB)相结合,而处于无活性状态,一旦细胞受到炎症因子刺激后,IκB将与NF-κB二聚体解离而使其p65亚基发生磷酸化并进入细胞核,致使NF-κB被激活,促使炎症发生[20]。近年来国内外的研究都指出NF-κB与肠道疾病之间的密切联系[21]。有研究表明,患有溃疡性结肠炎小鼠的结肠中发现通透性升高与肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)、NF-κB p65升高呈正相关[22];在肠道黏膜上皮处检测到NF-κB的表达增多,促炎细胞因子也随之增加,加剧炎症反应[23]。肖南[24]等人研究发现,通过抑制NF-κB p65的活化,能够缓解葡聚糖硫酸钠盐诱导小鼠结肠炎的病理损伤,减少炎症细胞因子的释放,抑制结肠细胞凋亡。据报道巨噬细胞过度活化可以激活NF-κB通路,促进释放炎症细胞因子,引起组织炎症反应[25]。本研究结果表明,冷暴露后回肠组织内p-p65的蛋白表达水平上调,激活回肠内NF-κB通路,表明冷暴露能够促进炎症反应的发生。当肠道受刺激时,免疫巨噬细胞释放大量炎症细胞因子,能够引发克罗恩病、溃疡性结肠炎在内的多种炎症性疾病[26]。其机制是由于促炎细胞因子和抗炎细胞因子之间平衡被打破[27]。白细胞介素是介导免疫细胞活化和增殖的一类细胞因子,参与免疫反应的调节。本研究结果显示:冷暴露后小鼠回肠炎症细胞因子IL-1β和IL-6的蛋白表达有所增加。表明促炎细胞因子聚积,导致炎症性疾病的发生。
综上所述,本研究发现冷暴露能够破坏回肠组织紧密连接与机械屏障的完整性,增加回肠组织的通透性和疾病易感性,激活NF-κB信号通路,促进炎症反应的发生。
猜你喜欢
江苏安全生产(2022年8期)2022-11-01 09:14:32
小资CHIC!ELEGANCE(2021年36期)2021-10-15 14:36:34
现代临床医学(2021年4期)2021-07-31 07:55:54
艺术评鉴(2020年5期)2020-04-30 06:47:57
人大建设(2018年10期)2018-12-07 01:13:46
中成药(2017年9期)2017-12-19 13:34:27
CHINESE JOURNAL OF AERONAUTICS(2017年1期)2017-11-21 12:54:14
中成药(2017年5期)2017-06-13 13:01:12
中西医结合心脑血管病杂志(2016年20期)2016-03-01 04:20:37
医学研究杂志(2015年12期)2015-06-10 06:57:46
