
不同温度条件下水分和盐分胁迫对画眉草种子萌发的影响
2022-09-06 14:36万琪叶耘伶唐丽黄伟纪佳怡冯亚亭骆凯张瑞
草原与草坪 2022年3期
万琪,叶耘伶,唐丽,黄伟,纪佳怡,冯亚亭,骆凯,张瑞
(1. 海南大学热带作物学院,海南 海口 570228;2. 海南大学档案馆,海南 海口 570228)
画眉草(Eragrostis pilosa),是禾本科画眉草属一年生暖季型草本植物,广泛分布于热带和温带地区[1]。该草具有抗旱、抗病、耐高温、耐瘠薄、耐粗放管理以及冬季无防护可越冬等特点,被广泛应用于园林绿化、固土护坡等[2-3]。因其分蘖能力强,生长迅速,极易形成较密集的植被覆盖层,通常作为防风固沙以及水土保持的优选草种之一[4]。
种子萌发是高等植物生命周期中最为关键的阶段,易受到多种外界环境因子的综合影响,如光照、温度、水分、盐分等[5-6]。据统计,我国干旱及半干旱土地面积超过国土面积的50%,盐碱地面积约有1 亿hm2,约占全国土地面积的10%[7],严重影响了我国农业发展和生态环境建设[8]。加之,全球气候变暖已成为人类的共识。有研究表明种子萌发对水分和盐分胁迫的响应与萌发温度密切相关,如5 ℃/28 ℃下,NaCl 溶液对羊草(Leymus chinensis)种子萌发抑制作用最小,而在5 ℃/35 ℃下,对羊草种子萌发的抑制作用最大[9]。歪头菜(Vicia unijuga)种子萌发的抗旱性(基础水势值,Ψb)随温度的升高呈先升后降的趋势,20 ℃条件下种子发芽的Ψb最低,为-1.27 MPa[10]。在盐胁迫条件下,低温(10~20 ℃)对小麦(Triticum aestivum)种子萌发抑制效应最强[11]。此外,相比高温条件(25 ℃/15 ℃和35 ℃/25 ℃),低温(15 ℃/5 ℃)显著(P<0.05)降低甜高粱(Sorghum bicolor)种子的发芽率和发芽势[12]。截至目前,围绕画眉草种子萌发的研究已有许多[4,13-14],但关于温度与水分或盐分互作及种子萌发相应参数报道较少。此外,相比大粒种子来说,画眉草种子较小,前期建植困难。探究画眉草种子萌发对环境因子如温度、水分、盐分等的需求,对于寻求画眉草最佳建植方法和深入了解该物种的生态适应性具有重要意义。
基于此,本研究在不同温度条件下探究水分胁迫(PEG⁃6000)和盐分胁迫(NaCl)对画眉草种子萌发的影响,应用水势模型预测不同温度条件下画眉草种子萌发抗旱和耐盐能力,为画眉草在干旱、盐碱地区成功建植和推广提供理论参考。
1 材料和方法
1.1 试验材料
供试画眉草种子购自北京百斯特草业有限公司(原产地为美国),于4 ℃条件下储藏备用。种子萌发试验于2021年6~8月在海南大学海甸校区进行,种子千粒重为(0.3985 ± 0.0098)g,属于极小粒种子。挑选籽粒饱满、大小均一的画眉草种子进行种子萌发试验。
1.2 试验方法
根 据Michel 和Kaufmann[15]的方法及计算公式,分别设置0(CK)、-0.2、-0.4、-0.6、-0.8 MPa 的PEG-6000 溶 液 和0(CK)、-0.25、-0.75、-1.25 MPa 的NaCl 溶液。采用纸上萌发法(TP 法)在25、30、35 和40 ℃4 个恒温条件下进行。分别在25、30、35和40 ℃恒温条件下进行水分和盐胁迫试验。将种子整齐放置于直径为9 cm 的培养皿,分别加入4 mL 不同浓度的PEG⁃6000 和NaCl 溶液,将培养皿分别置于相应温度条件的恒温人工培养箱,12 h 光照/12 h 黑暗,湿度80%。每个处理4 次重复,每个重复50 粒种子。试验期间,为确保培养皿内溶液浓度相对恒定,用封口膜封口以防止水分蒸发,且每2 天更换1 次溶液。以胚根突破种皮作为判断种子萌发的标准,试验结束期以统计14 d 以上且所有处理连续3 d 不再有新萌发种子出现为准。当试验结束后,计算种子最终萌发率(Germination percentage)和萌发速率(Germina⁃tion rate)。第7 天测定幼苗胚根长和胚芽长,每重复测定10 株,计算其平均值。
种子最终萌发率=发芽终期全部正常幼苗数/供试种子数×100%
萌发速率=1/t50,t50指运用Germinator 软件包计算种子萌发数达到非硬实种子的50%所需的天数。
1.3 水势模型计算方法
根据水势模型描述种子萌发对水势的响应:
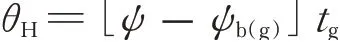
式中:θH为水势常数(MPa·h),表示达到发芽率g所需的累计水势,ψ表示环境水势,ψb(g)表示当发芽率达到g 时的基本水势或理论阈值,tg表示当发芽率达到g 时所需的发芽时间(h)。
通过重复概率单位回归分析(Repeated Probit)计算获得种子萌发的基础水势,水势时间值带入间隔为0.1 MPa/h,直到所获回归方程残差均方最小,根据上式计算水势时间累积值。
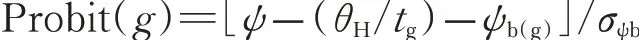
1.4 数据处理与分析
采用SPSS 20.0 软件,制图采用Excel 2019 软件进行数据处理。应用方差分析(ANOVA)检验各处理对测定参数的效应,若效应显著,采用Duncan 多重比较法比较各处理间的均值。
2 结果与分析
2.1 不同温度条件下画眉草种子萌发对水分和盐分胁迫的响应
双因素方差分析结果表明,温度、水分及两者互作极显著(P<0.01)影响画眉草的萌发率和萌发速率。除温度和盐分互作不显著影响画眉草种子萌发率外,温度、盐分及两者互作均极显著(P<0.01)影响画眉草的萌发率和萌发速率(表1)。
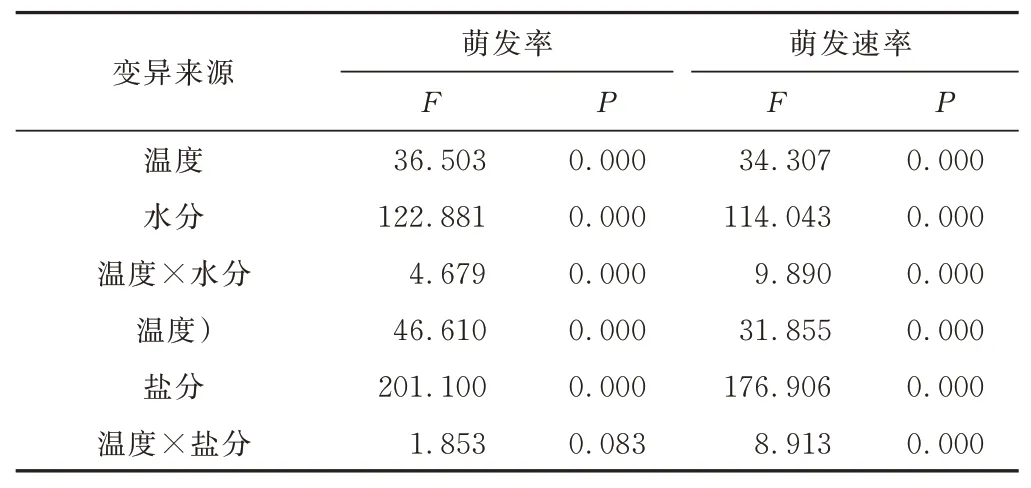
表1 温度、水分/盐分及其互作的画眉草种子萌发率和萌发速率方差分析Table 1 Two⁃way ANOVA for the effects of temperature,water stress/salt stress and their interaction on seed germina⁃tion percentage and rate ofE. pilosa
画眉草种子萌发率和萌发速率均随水分和盐分胁迫程度的增加呈下降趋势,且下降幅度因温度不同而异(表2)。水分胁迫条件下,在0、-0.2 和-0.4 MPa 下,画眉草种子萌发率在30、35 和40 ℃之间无显著差异,但均显著高于25 ℃(P<0.05);而在-0.6和-0.8 MPa 下,30 ℃条件下种子萌发率最高,显著高于25 ℃和40 ℃;盐分胁迫条件下,在-0.25、-0.75和-1.25 MPa 下,画眉草种子萌发率在30、35 和40 ℃之间无显著差异,但均显著高于25 ℃。
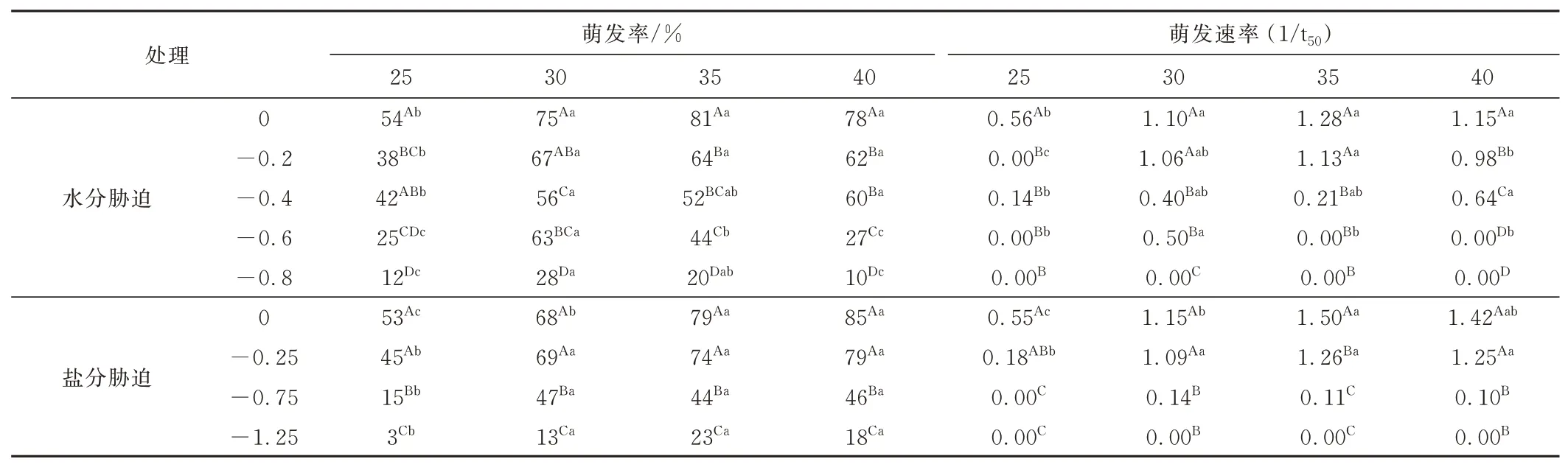
表2 不同温度条件下水分和盐分胁迫画眉草种子的萌发率和萌发速率Table 2 Effects of water stress and salt stress on seed germination and germination rate(1/t50)ofE. pilosaunder different temperatures
水分胁迫条件下,在0、-0.2 和-0.4 MPa 下,画眉草种子萌发速率在30、35 和40 ℃均高于或显著高于25 ℃(P<0.05);在-0.6 MPa 下,30 ℃条件下的萌发速率显著高于其他温度条件下。盐分胁迫条件下,在0、-0.25 MPa 下,画眉草种子的萌发速率在30、35和40 ℃均显著高于25 ℃,且35 ℃条件下种子的萌发速率最快;而在-0.75 和-1.25 MPa 下,各温度条件下的萌发速率无显著差异。
随水分和盐分胁迫的增加,画眉草幼苗生长整体上表现为先促进后抑制(图1)。同一水分胁迫条件下,胚芽长随温度的升高而降低,而胚根长则呈先增加后减小的趋势。其中,除-0.6 MPa 条件下,幼苗胚根长表现为35 ℃显著(P< 0.05)高于30 ℃外,其他水分条件下,30 ℃时画眉草幼苗生长均表现最佳。当水分胁迫达-0.8 MPa 时,30 ℃条件下幼苗胚芽长和胚根长分别为0.4 cm 和3.4 cm,而当温度达40 ℃时,幼苗不生长。同一盐分胁迫条件下,胚根长随温度的升高呈先增加后减小趋势,且在30 ℃时幼苗生长表现最佳,在-0.75 MPa 下,30 ℃条件下幼苗胚芽长和胚根长分别为1.1、2.7 cm。

图1 不同温度条件下水分和盐分胁迫画眉草幼苗的胚芽长和胚根长Figure 1 Effect of water stress and salt stress on growth of seedling shoot and root ofE. pilosaunder different temperatures
2.2 基于Probit 分析估计不同温度条件下画眉草种子萌发基础水势
由重复Probit 回归分析获得画眉草种子萌发的基础水势值(ψb(50))随温度的升高呈先降低后升高的趋势。水分胁迫条件下,种子萌发所需的基础水势值表现为25 ℃>40 ℃>35 ℃>30 ℃,其中以30 ℃处理最低,为-0.75 MPa,表明在30 ℃条件下有利于画眉草种子在干旱环境下萌发。盐分胁迫条件下,种子萌发所需的基础水势值表现为25 ℃>30 ℃>40 ℃>35 ℃,其中以35 ℃时最低,为-0.78 MPa,表明在35 ℃条件下有利于其在盐碱环境萌发。此外,水分胁迫条件下画眉草种子萌发所需的θH值随温度升高呈下降趋势,具体表现为25 ℃(16.5)>30 ℃(15.5)>35 ℃(9.0)>40 ℃(6.5),而盐分胁迫条件下θH值则随温度升高而增加,具体表现为25 ℃(3.0)<30 ℃(5.0)<35 ℃(7.0)=40 ℃(7.0)(表3)。

表3 基于重复Probit 回归估计不同温度条件下画眉草种子萌发的基础水势Table 3 Estimation of water potential value based on repeated probit analysis ofE. pilosaunder different temperatures
3 讨论
温度是影响种子萌发的关键环境因子,通过改变种子萌发过程中膜的通透性、膜结合蛋白能力以及影响种子内部的酶活性和物质代谢,进而影响种子萌发及幼苗生长[16-19]。本研究发现25 ℃显著降低画眉草种子萌发率,而40 ℃虽促进画眉草种子的萌发,却抑制幼苗生长,与马亚杰等[20]关于环境因素对牛筋草种子萌发的影响结果相似,这可能是由于高温虽能促进种子萌发,但温度过高导致后期幼苗生长过程中蒸腾加剧、气孔关闭,造成光合作用减弱,进而减缓或抑制幼苗生长。此外,画眉草种子萌发率和萌发速率随水分和盐分胁迫的增加呈下降趋势,这与齐淑艳等[21]关于入侵植物牛膝菊(Galinsoga parviflora)种子、黄勇等[22]对石竹(Dianthus chinensis)种子、王景瑞等[23]对干旱荒漠区4 种一年生植物种子及徐曼等[24]对长穗偃麦草(Elytrigia elongate)种子萌发的研究结果相似。
种子萌发是种子对逆境胁迫最为敏感的时期,在干旱或盐碱环境下,温度过高或过低对种子造成的伤害或对种子萌发的抑制作用更大[11]。当水分胁迫低于-0.6 MPa 时,在25、35 和40 ℃温度条件下画眉草种子萌发率显著(P<0.05)低于30 ℃条件下,且在30 ℃条件下种子萌发的基础水势最低,为-0.75 MPa,表明在30 ℃条件下画眉草种子萌发时对干旱胁迫的耐受性最强,较低温度或较高温度降低了画眉草种子萌发时的耐旱性。唐伟等[10]研究发现,在较低水势(-1.0~-0.6 MPa)条件下,20 ℃时歪头菜种子萌发和幼苗生长显著(P<0.05)高于15 ℃和25 ℃,表明适宜的萌发温度能够一定程度缓解水分胁迫对种子萌发的抑制作用。此外,画眉草种子萌发的耐盐性在不同温度条件下表现各异,特别是低温显著(P<0.05)降低了其萌发阶段的耐盐性,与雷春英等[25]对濒危植物盐桦(Belula halophila)的研究结果相似。王志恒等[12]发现,随盐分胁迫增加,低温(15 ℃/5 ℃)对甜高粱(Sorghum bicolor)种子萌发的抑制作用较高温(25 ℃/15 ℃和35 ℃/25 ℃)更大。蔺吉祥等[11]研究也表明,随盐分胁迫增加,低温条件下(10~20 ℃)小麦种子的萌发率显著(P<0.05)低于15~25 ℃,而高温25~35 ℃则与15~25 ℃无显著(P>0.05)差异。与上述研究结果一致,低温(25 ℃)和高盐(-1.25 MPa)双重胁迫对画眉草种子萌发抑制作用更强,而在较高温度条件下有利于种子在盐胁迫环境顺利萌发,究其原因可能是在低温条件下,种子吸水难度增加,抗氧化物酶活性降低,胚的生长受到抑制,且种子在低温环境下容易被诱导休眠,一定程度上降低了种子萌发的耐盐性[26]。
水 势 模 型(hydrotime model)是 由Gummerson[27]提出,用于描述萌发速度与水势梯度之间的关系,用以预测胁迫条件下种子的萌发策略。本研究发现,画眉草种子萌发的基础水势值(ψb(50))随温度的升高呈先降低后升高的趋势,表明适宜的温度条件能够降低种子萌发对水分的需求。此外,相同温度条件下画眉草种子萌发对水分的需求即基础水势值因胁迫类型不同而存在差异,这可能是由于PEG 处理对种子萌发的抑制作用主要归因于渗透效应,而NaCl 处理除渗透效应外还存在离子效应等[28]。本研究结果表明,水势为-0.2 MPa PEG 处理条件下,画眉草种子的萌发率和萌发速率在不同温度条件下均低于-0.25 MPa NaCl,说明在高渗环境下水分胁迫对种子萌发的抑制作用大于盐分胁迫,与张紫薇等[29]对藜麦(Chenopo⁃dium quinoa)种 子 和张 红 香等[28]对 大 麦(Hordeum vulgare)种子的研究结果一致,究其原因可能是种子在NaCl 胁迫下能够从溶液中吸收少量的无机离子以增加细胞溶液浓度来进行渗透调节,而PEG 是大分子物质,不能自由进入细胞从而使种子内部形成较低的水势,因而萌发抑制作用更强[30]。
4 结论
水分、盐分胁迫及其与温度交互作用显著影响画眉草种子萌发和幼苗生长,25 ℃抑制种子萌发,而40 ℃虽能促进种子萌发,但却抑制其幼苗生长。随水分和盐分胁迫强度的增加,画眉草种子的萌发率和萌发速率均呈降低趋势,其中,低温是影响种子萌发抗旱性的主要因素,而高温加剧了盐分胁迫对种子的伤害。
猜你喜欢
安全与环境工程(2021年5期)2021-10-08
祝您健康·文摘版(2019年3期)2019-06-11
阅读与作文(小学低年级版)(2018年9期)2018-10-26
阅读与作文(小学低年级版)(2018年10期)2018-10-25
意林·少年版(2017年15期)2017-09-05
中学化学(2016年10期)2017-01-07
读者·校园版(2016年6期)2016-03-07
中学化学(2015年8期)2015-12-29
文苑(2011年10期)2011-08-15
祝您健康(1985年1期)1985-12-29
