
肌联蛋白与运动研究进展
2022-09-05 02:08:24苏香楠高扬于亮
中国运动医学杂志 2022年7期
苏香楠 高扬 于亮
北京体育大学(北京100084)
肌联蛋白(titin)作为一种骨架蛋白,跨越了从Z 盘到M 线半个肌小节[1],能够通过对齐肌球蛋白使邻近肌小节保持稳定,是骨骼肌纤维中除了肌动蛋白和肌球蛋白以外最丰富的蛋白。此外,肌联蛋白可调节肌肉主动收缩,并与肌球蛋白和肌动蛋白共同作用,对粗肌丝和细肌丝进行精准调控以提升肌肉收缩力量[2]。不同方式[3]和强度[4]的适度训练,不仅能使肌原纤维中粗肌丝肌球蛋白和细肌丝肌动蛋白合成水平提高,还会使肌联蛋白合成水平出现同一趋势变化,并伴随着肌肉张力增加。目前,已有研究证实肌联蛋白结构改变可增强或抑制肌肉力量[5-6],并在运动过程中发挥重要作用。另一方面,肌联蛋白“分子弹簧”效应[7]也逐步明确,当适宜外力拉伸骨骼肌时,肌联蛋白会增加刚度以增强肌力[8],若肌小节受到过度牵拉时,肌联蛋白的弹性成分会缓冲以保护肌肉[9]。基于肌联蛋白在运动过程中收缩、舒张、影响力量和缓冲等多方面的潜在作用,本文通过检索截至2022 年6 月在CNKI、万方及Pubmed、Web of science 等数据库中“肌联蛋白”、“骨架蛋白”、“收缩蛋白”、“肌联蛋白与运动”、“titin”、“skelemin”、“titin AND exercise”等关键词,梳理肌联蛋白结构、功能以及运动对肌联蛋白的影响等文献资料,结合其在运动过程中的作用,对肌联蛋白与运动的关系进行归纳总结,以期为进一步研究提供参考依据。
1 概述
1.1 基本情况
肌联蛋白又称为“第三肌丝”,是骨骼肌纤维中第三丰富的肌肉蛋白(仅次于肌球蛋白和肌动蛋白)。在功能上,肌联蛋白有维持肌小节长度、保持肌小节完整、缓冲收缩舒张等作用[10-12];在结构上,肌联蛋白由一系列模块化的弹性结构串联组成,主要分为四个区域:Z 盘(N 端)、I 带、A 带、M 线(C 端),其主要物理特性是由A带和I带决定的,肌联蛋白的弹性成分位于I带,而A 带区域几乎不可伸展[13-16]。其中,I带中心部分(图1)弹性区域由串联排列的Ig 重复序列组成,这些重复序列包含在两个弹性结构中,即近端和远端串联Ig 结构域。它们之间串联的免疫球蛋白组由2200 多个残基隔开,这些残基富含脯氨酸(proline,P)、谷氨酸(glutamate,E)、缬氨酸(valine,V)和赖氨酸(lysine,K),因此被称为“PEVK 区”,而近端Ig 结构域和PEVK 区被N2A 区隔开,N2A 区被称为第三区域。肌联蛋白的I带部分包含多达100 多个免疫球蛋白样结构域,是横纹肌组织具有被动刚度的主要原因[17-19]。
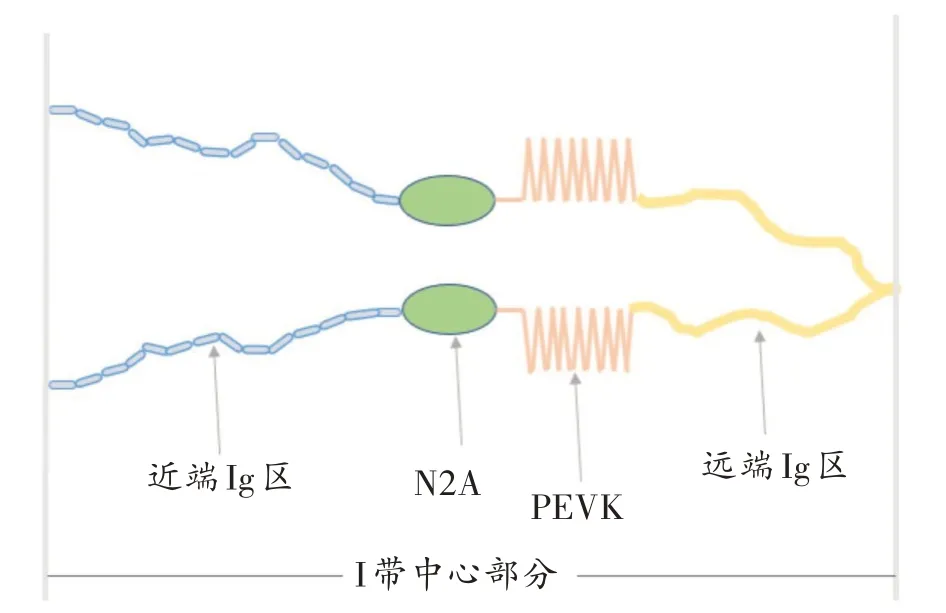
图1 肌联蛋白I带中心部分示意图
1.2 肌小节中肌联蛋白的分布
采用免疫电镜研究观察10 种非重复的肌联蛋白单克隆抗体,发现单分子的肌联蛋白从Z 盘延伸到接近M 线,包括Z 盘上肌联蛋白[肌联蛋白的氨基末端(≤80 kDa)跨越了整个Z 盘区域,来自肌小节的肌联蛋白肌丝在Z 盘内重叠]、I 带内肌联蛋白(800~1500 kDa 的肌联蛋白)、A 带上肌联蛋白(肌联蛋白羧基端2000 kDa)、M 线上肌联蛋白(肌联蛋白羧基末端约200 kDa)[20]。六个肌联蛋白丝与每个粗肌丝相连,且每个肌联蛋白丝也与单个细肌丝相连[21]。
1.3 收缩过程中的肌联蛋白
经典的肌肉收缩理论“肌丝滑行学说”充分描述了骨骼肌在向心收缩和等长收缩过程中的大部分功能,但是从“肌源”角度解释骨骼肌在离心运动状态下比向心运动产生更大张力的原因尚不明确,可能是由于肌肉内部存在“分子弹簧”结构。肌联蛋白附着在A 带的粗肌丝上,且可以自由延伸至I 带区域,在生理条件下像“分子弹簧”一样工作,以增加张力产生并能够抵抗过度拉伸以防止肌节损伤。当肌肉收缩时,肌联蛋白不仅与肌球蛋白对齐并稳定相邻的肌节,而且部分释放的Ca2+能够与肌联蛋白结合,改变肌肉刚度[22]。因此,探究肌联蛋白结构及其在骨骼肌中的作用机制是深入研究骨骼肌运动能力的重要任务。
2 肌联蛋白的作用
2.1 肌肉的“防护器”:肌联蛋白的稳定作用
肌联蛋白可以维持肌小节的稳定性。从结构上来看,一方面肌联蛋白分子将细肌丝固定在Z 盘上[23],贯穿粗肌丝的整个肌小节直到M 线[20],形成了六角形网状结构,在横向和纵向上维持每个肌小节结构完整[9]。而且肌联蛋白的伸展性又为肌小节提供了较大的被动张力,增强了相邻肌小节的稳定性[24]。另一方面,肌联蛋白的I带由一系列弹性结构组成,可以抵抗肌小节过度拉伸[9]。有研究对蛙完整肌纤维中I带肌联蛋白的弹性进行了定量分析,发现肌小节长度2.7~3.1 μm 范围内,肌联蛋白会出现“自我适应”现象使弹性保持不变[25],这可能是由于在肌小节较长的情况下,I带宽度增加保持了肌联蛋白弹性长度[26]。虽然不同种属之间肌肉的粗、细肌丝长度存在差异,但由于人和蛙具有相同的粗肌丝长度,其最适长度平台期的长度相同且超过最适长度时的下降斜率也相似[27],因此,当人的肌小节过度拉伸时,其肌联蛋白I 带长度会增加以保持弹性不变。总之,肌联蛋白作为肌肉的“防护器”,在收缩过程中不仅能够对齐肌球蛋白位置、稳定相邻肌小节,还能防止肌肉在被动拉伸时肌小节过度伸展造成的损伤。
2.2 肌肉的“强助力器”:肌联蛋白的“分子弹簧”功能
肌联蛋白作为哺乳动物体内分子量最大的蛋白,其肽链具有“分子弹簧”功能区,在肌肉收缩和拉伸中起着重要作用。当肌肉主动收缩时,肌联蛋白可以在适度收缩后(图2)促使肌小节恢复至初始长度,而在被动拉伸后,肌联蛋白能够牵拉M 线和相对运动的肌丝,起到缓冲的作用[28]。另外,肌联蛋白在很大程度上决定肌纤维的被动刚度[29],抵抗过度张力的同时,还可以使肌小节在舒张过程中恢复到适当的静止长度(图3)。另外,Labeit 等[30]研究发现肌联蛋白在肌肉激活时与Ca2+结合,增加了肌联蛋白的刚度,从而增加了肌肉拉伸时的力,但也有研究在肌肉运动后测量发现肌联蛋白刚度与肌肉力量变化无关,具体原因尚未形成统一结论,后续研究可深入探讨。作为肌肉的“强助力器”,肌联蛋白的“分子弹簧”功能在肌肉运动中发挥着重要作用。
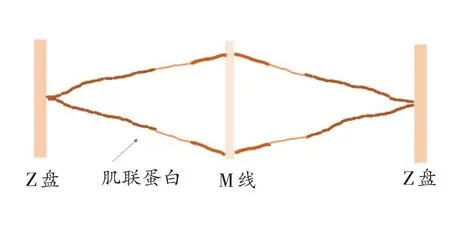
图2 肌联蛋白收缩状态
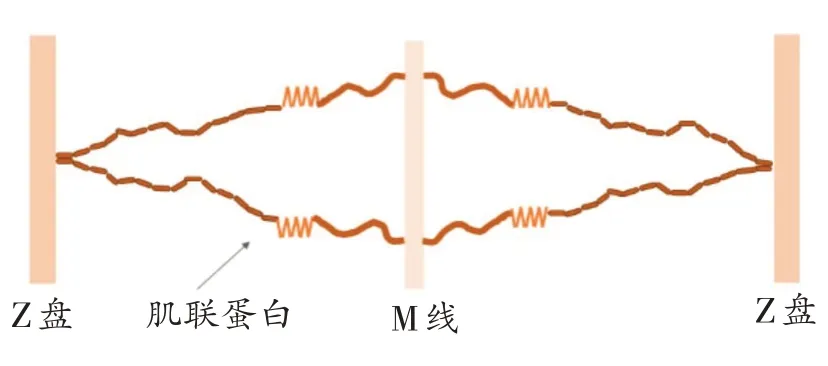
图3 肌联蛋白舒张状态
2.3 肌肉的“调节器”:肌联蛋白与肌动蛋白、肌球蛋白的相互作用
除了肌联蛋白的刚度,肌小节的被动张力也可以通过肌联蛋白与肌动蛋白和肌球蛋白的相互作用[31]实现调节。骨骼肌收缩过程中残余力的增强可能会产生和肌联蛋白相关的附加效应。目前有学者认为,当Ca2+浓度升高时,在骨骼肌离心收缩过程中,特定的肌联蛋白弹性结构可能与肌动蛋白相互作用,导致其肌联蛋白长度缩短,并且在离心运动期间以及收缩后的一段时间内增加其被动张力[32]。当肌联蛋白的N2A 区与肌动蛋白结合时,肌原纤维被动拉伸,I 带近端串联Ig 结构域的力容易受到影响[33]。因此,当被动拉伸Ca2+激活的肌小节时,只有刚度较高的肌联蛋白的PEVK 区会被拉长,从而产生更大的力[34]。假设肌联蛋白是一个可调节的“分子弹簧”,离心运动中许多无法说明的现象都可以通过横桥理论的框架得以解释[35]。
另外,还有学者提出了“winding filament”假说,该假说认为在Ca2+内流后肌联蛋白被激活,由于横桥运动,肌联蛋白缠绕在细肌丝上,使肌动蛋白形态略有改变,这些横桥可以支配和调节细肌丝[36]。肌联蛋白与依赖于细肌丝的Ca2+结合解释了主动拉伸时的力量比被动拉伸时增加的多的原因,可能是由于骨骼肌肌联蛋白的N2A 区在激活时与肌动蛋白结合,增加了肌联蛋白的刚度所致。此外,肌动蛋白与肌球蛋白末端肌丝之间的相互作用可能会缩短肌联蛋白远端Ig结构域和部分PEVK 区,增加肌联蛋白的刚度,进而增加肌联蛋白在进行主动拉伸时的力量且不需要额外的能量[37]。由此可见,肌联蛋白与肌动蛋白、肌球蛋白之间相互协同,作为肌肉的“调节器”,在收缩或舒张过程中增加肌肉刚度和肌肉力量。
3 运动对肌联蛋白的影响
3.1 急性运动和长期运动对肌联蛋白的影响
肌联蛋白作为维持肌细胞张力的重要结构蛋白,对调节肌小节被动张力具有重要作用,已有动物研究发现,急性运动后大鼠比目鱼肌肌联蛋白含量明显下降,48 小时后恢复正常水平[38],而在人体研究中也发现成年男性在急性离心运动后肌联蛋白水平降低[39],这与动物研究结果相一致,表明了在急性运动后骨骼肌出现了肌联蛋白合成代谢减少、分解代谢增加等微细结构损伤,肌联蛋白降解或解聚使维持骨骼肌正常收缩功能的骨骼肌细胞骨架受到破坏,在功能上表现为收缩能力下降,但随后骨骼肌收缩功能逐渐恢复至正常水平,这可能与恢复期时长有关。而长期运动对肌联蛋白的影响与急性运动有所不同,有研究发现[3],进行8 周有氧运动干预后,糖尿病大鼠肌联蛋白免疫荧光染色区域明显增多,平均荧光强度明显增高,提示有氧运动可以增加肌联蛋白的表达。上述结果表明,虽然长期运动与肌联蛋白的表达存在一定关系,但随着运动时间的延续是否会对肌联蛋白结构产生其他影响尚无定论。
3.2 超等长收缩运动对肌联蛋白的影响
肌联蛋白作为骨架蛋白,几乎与其他所有肌小节蛋白相互作用,因此当肌联蛋白剪切或缺失时会破坏肌小节结构[40]。有研究发现超等长收缩训练后肌小节出现不同程度的损伤,Macaluso 等[4]在实验干预前对受试者进行一次最大超等长收缩测试,发现受试者肌小节Z 盘完全被破坏,这提示急性超等长收缩训练可能会引起肌联蛋白受损。而进行8 周最大深蹲跳训练后再次进行最大超等长收缩测试,却发现受试者肌小节Z盘数目并未减少或受损,表明长期超等长收缩训练可能加强了对肌小节及肌联蛋白的保护。除此之外,超等长收缩训练还有助于改善运动经济性,且肌联蛋白亚型T1越低,运动经济性越高[41]。由此可见,长期超等长收缩训练不仅会增加肌肉爆发力[42],还会加强肌联蛋白的保护作用,达到预防损伤的效果,同时,可以通过提高肌联蛋白亚型T1 改善运动经济性,减少能量消耗。
另外,离心和等长收缩阶段的张力增强与肌联蛋白也密切相关。肌联蛋白作为一种表现为弹性肌丝的大分子蛋白质,在肌肉舒张过程中仅附着在Z 盘和M线上,但在离心收缩阶段一部分肌联蛋白附着在细肌丝上,肌联蛋白丝变得更硬,这使得肌联蛋白与肌动蛋白的连接以及由此增强的刚性在随后的等长收缩期得以保持[43-44]。而进行超等长收缩训练时肌肉先做离心式拉长继而做向心式收缩,该过程也涉及了离心收缩阶段,间接表明了超等长收缩训练与肌联蛋白之间也存在一定联系。
4 肌联蛋白在运动中的作用
4.1 肌联蛋白在被动牵张中的作用
肌联蛋白可增加骨骼肌被动牵张过程中的张力。有研究表明,单根肌纤维[45]中90%~95%的被动张力以及骨骼肌[7]中大部分被动张力由肌联蛋白提供,但是当肌联蛋白被破坏后,肌肉的被动张力可能出现下降或消失。有研究发现使用胰蛋白酶[46]或者采用电离辐射[47]破坏肌联蛋白时,肌肉的被动张力降低了50%~100%,而且当肌联蛋白I 带被水解或从粗肌丝上移除后,肌丝长度在其生理范围内几乎没有被动张力产生[48]。
另外,肌联蛋白I带长度能够影响肌联蛋白的被动张力大小。肌联蛋白I 带具有弹性[49],当进行被动拉伸时,肌联蛋白可以通过结合PEVK 区和Ig 结构域中的Ca2+增加其刚度,缩短I 带长度以产生被动张力[50]。一般来讲,肌联蛋白I 带越短,被动张力越高[51]。此外,肌肉的长度也可以影响肌联蛋白的被动张力[39]。当肌小节长度较短时,被动拉伸会导致折叠的Ig结构域变直,被动张力变化不大。但当肌小节长度较长时,PEVK 区延长,被动张力明显增加[52]。但近些年来也有研究指出,剪切部分PEVK 区会增加被动张力,部分原因是肌联蛋白弹性区拼接发生了改变,这表明了肌联蛋白的PEVK区域在调节被动张力方面的作用[2]。当肌小节长度在其生理范围内时,I 带的PEVK 区的延长很大程度上决定了骨骼肌纤维的被动弹性。
4.2 肌联蛋白在主动收缩中的作用
作为“第三肌丝”,肌联蛋白不仅在骨骼肌被动牵张过程中进行调节,在主动收缩时也发挥重要作用[51]。Higuchi 等[32]将肌联蛋白从小鼠的肌原纤维中消除后,肌纤维的主动张力降低了50%,这可能是由于肌联蛋白被破坏时,粗肌丝在向另一侧的Z 盘移动,肌小节中的粗肌丝排列紊乱,粗肌丝和细肌丝之间的重叠减少,从而使主动张力变小。另外,Powers 等[53]采用离体实验将无任何干预的正常小鼠肌肉的肌原纤维放置在Ca-ATP 的激活溶液中以完成主动拉伸,发现张力增加的大部分原因是肌联蛋白与细肌丝的结合使主动张力发生了变化,而少部分则归因于Ca2+对肌联蛋白的直接作用,推测可能由于Ca2+与PEVK 区的E-rich 区以及部分Ig 结构域结合时,蛋白质刚度增加,从而增加张力。Nishikawa 等[36]也认同这一观点,发现肌营养不良伴肌炎型(the muscular dystrophy with myositis,mdm)小鼠的肌联蛋白突变导致PEVK 区和N2A 区缺失83个氨基酸,在Ca2+激活的过程中,mdm 小鼠比目鱼肌无刚度变化,且在主动收缩过程中mdm 小鼠的张力并未发生明显变化,可见mdm 小鼠中缺失的序列影响了肌联蛋白在主动收缩中的作用。但也有学者认为肌联蛋白可能是通过调节粗肌丝和细肌丝之间的距离来提升主动张力[3]。
4.3 肌联蛋白在肌力增强和抑制方面的作用
由于肌联蛋白由一系列模块化的弹性结构串联组成,所以在横桥产生的全部力量传递到肌肉附着的骨之前,这些串联弹性结构必须完全被拉伸[54]。而在离心收缩过程中,由于肌肉被完成激活且串联弹性结构被拉长,肌肉张力显著增强[55]。另外,为了研究肌联蛋白在肌肉增强和抑制方面的作用,有学者对mdm 小鼠和野生型小鼠进行离体主动和被动拉伸实验,发现野生型小鼠肌肉肌力增加,但mdm小鼠肌力无明显变化,这可能是由于mdm 型小鼠的骨骼肌肌联蛋白N2A 和PEVK区氨基酸缺失[8-9],抑制了肌力的增加。而力量产生的原因可能是骨骼肌被Ca2+释放激活时,一部分Ca2+与肌联蛋白结合,增加了其刚度。由此可见,肌联蛋白可以增强肌肉张力。相反,当肌联蛋白缺失时肌张力降低。Joumaa 等[46]在兔模型中同样观察到了肌联蛋白降解后肌张力降低现象,当使用胰蛋白酶消除法使肌联蛋白部分降解时,骨骼肌主动张力和被动张力均显著降低。可见,肌联蛋白在肌力增强和抑制方面发挥了重要作用。
5 小结
肌联蛋白可以维持肌小节的稳定性,具有分子弹簧的作用。肌联蛋白与肌球蛋白、肌动蛋白相互作用,在骨骼肌被动牵张、主动收缩中都可起到强化、缓冲等辅助效果,具有调节增强肌力、在适度范围内抑制肌力、保护骨骼肌降低损伤风险的作用。建议进一步研究不同运动强度、运动时间训练模式对肌联蛋白结构和刚度的影响,以及不同类型肌纤维中肌联蛋白的作用,为解决运动科学领域中与肌联蛋白相关的骨骼肌收缩问题以及更加科学地制订力量训练方案提供依据。
猜你喜欢
中国畜牧杂志(2020年1期)2020-01-16 04:09:54
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
医学研究杂志(2015年5期)2015-06-10 06:43:26
生命科学研究(2015年2期)2015-04-29 23:09:30
中国医科大学学报(2015年10期)2015-03-01 02:09:59
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:43
心血管病学进展(2015年4期)2015-02-22 07:56:44
中国医学科学院学报(2013年6期)2013-03-11 20:26:00
实用老年医学(2013年7期)2013-03-11 18:39:07
