
猕猴桃多酚在羊肉反复冻融过程中的改善肉质和延缓氧化作用
2022-09-01 08:01张朵朵刘永峰古明辉杨泽莎张昊阳
食品科学 2022年16期
张朵朵,刘永峰,古明辉,杨泽莎,张昊阳
(陕西师范大学食品工程与营养科学学院,陕西 西安 710119)
羊肉因具有丰富的营养物质而被大众所喜爱,冷冻贮藏作为肉制品在流通运输时的一种贮藏手段,可以降低酶活性并抑制微生物增殖,但在羊肉冻结时,冰晶体积增大,破坏细胞膜,解冻后引起汁液流失,损失蛋白等营养成分及众多风味物质,并降低肉及肉制品的品质;更值得关注的是,由于在流通过程中羊肉冷链的不健全,例如冷链“断链”问题严重以及生鲜冷链物流市场化程度不高等都会使肉多次冻融,特别在偏远地区冷链设施落后,冷冻肉在贮运期间冻融现象更严峻。李金平等将牛外脊置于-16 ℃冷冻24 h、2 ℃解冻24 h的冻融循环中,观察到肉值逐渐降低,且在冻融5 次后下降更为显著;Cheng Shasha等也研究发现牛半膜肌在经过多次循环冻融后值显著降低,这可能是由于反复冻融中水分损失引起相关色素物质流失所致。邓思杨等冻融次数对镜鲤鱼肌原纤维蛋白功能和结构特性变化的影响,冷冻-解冻循环破坏了镜鲤鱼肌原纤维蛋白的完整结构,进而造成鱼肉的硬度降低。发现冻融循环中形成冰晶可使肌原纤维断裂,反复冻融也会引起肉脂肪氧化,进而影响产品的感官和保质期。Chen Qingmin、Ali等研究显示,随着冻融循环次数的不断增加,肉中的脂肪氧化程度逐渐加深。彭新颜等对生猪肉糜中进行不同冻融循环(0~9 次)处理,发现猪肉中脂肪的氧化程度随着冻融次数的增加而加深,并且肌肉组织变硬,最终导致风味变差。氧化反应是肉类在冷冻储运过程中肉质劣变的最主要机制之一,而羊肉由于有较高的肌红蛋白含量和铁氧化水平而具有较强的过氧化势生成潜能,再加上其他内外部因素,致使羊肉中氧化反应加剧,引起脂质降解和蛋白交联,因此在多次冻融循环过程中羊肉品质明显下降。
在肉制品中添加抗氧化剂是减少组织损伤及改善氧化应激的主要方法之一,然而,化学添加剂自身潜在的毒副作用和残留问题将严重影响肉品品质、食品安全及人体健康。已有研究表明,某些天然抗氧化剂对脂质和蛋白质氧化除了具有一定的抑制作用,还可以有效改善肉品理化特性和感官品质。在用天然抗氧化剂对肉品保鲜的研究中发现,苹果幼果酚类提取物具有延缓脂质氧化、护色、抑制蛋白降解的作用,从而有效改善肉品品质。将板栗壳、叶和茎中的酚类提取物添加到牛肉中对其保鲜效果进行研究,结果发现添加0.05%板栗壳提取物对维持牛肉饼中肌红蛋白含量稳定有一定的效果,而添加0.1%叶片中酚类提取物具有抑制牛肉脂质氧化的最佳效果。此外,大量研究表明,植物多酚可以降低肉制品中硫代巴比妥酸反应物(thiobarbituric acid reactive substances,TBARS)值和挥发性盐基氮(total volatile base nitrogen,TVB-N)值,说明植物多酚能够延缓脂肪氧化,减少氨基酸的破坏,保持肉类的新鲜度,提高肉制品品质。中国是世界猕猴桃生产大国,由于“疏花疏果”处理,每年有大量疏果被丢弃在果园中,有研究表明猕猴桃疏果富含多酚类抗氧化剂,可以作为天然抗氧化剂将其用于改善肉品质的研究。因此,将猕猴桃疏果用于冻融过程中改善肉品质的天然抗氧化剂,可以变废为宝。目前关于猕猴桃及其活性成分利用的研究较多,然而缺少其改善肉类品质方面的研究,尤其是猕猴桃疏果多酚的应用报道更为少见。
基于此,本研究以陕北白绒山羊的背最长肌为实验材料,通过分析比较猕猴桃疏果多酚(后称疏果多酚)和表儿茶素、叔丁基对苯二酚(-butyl hydroquinone,TBHQ)3 种抗氧化剂在羊肉反复冻融中调节肉品质的作用,探究疏果多酚在抑制羊肉脂肪氧化、稳定其理化特性、延缓腐败变质、改善肉品质等方面的优势作用,旨在为疏果多酚在肉品保鲜中的应用提供理论参考和技术支撑,从而实现将猕猴桃疏果综合应用于肉品产业中。
1 材料与方法
1.1 材料与试剂
以宰后24 h的1.5 周岁左右健康的陕北白绒山羊(公)的背最长肌为研究对象。疏果多酚:参考Jiao Yang等处理方式,经过冷冻干燥制成冻干粉,再经X-5大孔树脂纯化冻干,最后用高效液相色谱进行定性定量分析,其主要成分为表儿茶素((139.45±3.54)mg/g)、槲皮素((36.03±1.23)mg/g)、儿茶素((5.84±0.24)mg/g)。
无水乙醇、硼酸、氧化镁、无水硫酸铜、硫酸钾、石英砂、石油醚、氯仿、甲醇、正己烷(均为分析纯)天津市科密欧化学试剂有限公司;山梨酸钾 天津市福晨化学试剂厂;表儿茶素(纯度≥98%) 安徽酷尔生物工程有限公司;TBHQ(纯度≥98%) 润友化学有限公司。
1.2 仪器与设备
TA.XT.Plus质构仪 英国Stable Micro System公司;NS800分光测色仪 深圳市三恩驰科技有限公司;ME21数码生物显微镜 日本奥林巴斯公司;2010 ultra气相色谱-质谱联用仪 日本岛津公司;UV-1200型紫外-可见分光光度计 上海美析仪器有限公司;PHS-3C精密pH计 上海仪电科学仪器股份有限公司;BCD-578WPU9CX风冷冰箱 长虹美菱股份有限公司。
1.3 方法
1.3.1 样品前处理
对购买的3 只山羊胴体的背最长肌(每块背最长肌2 kg左右),剔除脂肪组织、筋膜,进行等厚(1.5 cm左右)切分,进行实验。
把每块山羊胴体的背最长肌作为单个样本,分为空白组、疏果多酚组、表儿茶素组和TBHQ组4 组,每组3 个平行,共12 份(每份肉120 g)。疏果多酚组、表儿茶素组、TBHQ组分别以猕猴桃疏果多酚、表儿茶素、TBHQ为溶质,均用蒸馏水为溶剂配制1 mg/mL的溶液,空白对照组以蒸馏水处理山羊肉;分别在空白组、疏果多酚组、表儿茶素组和TBHQ组的肉样中按照肉液比1∶1(/)添加蒸馏水、猕猴桃疏果多酚溶液、表儿茶素溶液和TBHQ溶液120 mL,在0~4 ℃浸泡15 min,然后在0~4 ℃将水分沥干至无水滴落下,自封袋包装。将4 组样品置于-18 ℃冰箱中冷冻24 h后取出,4 ℃冷藏解冻12 h,此为1 个冻融处理。此方法分别进行1、3、5 次冻融处理,每次处理结束后,分别在12 份肉样中称取30 g样品用于后续指标测定。
1.3.2 疏果多酚处理后肉品质指标的测定
1.3.2.1 汁液流失率

式中:为解冻前肉样的质量/g;为解冻后用吸水纸吸干的肉样质量/g。
1.3.2.2 显微结构测定
参考Guo Yuhan等的方法,每个处理选择3 个样品,将肉样用体积分数4%的多聚甲醛溶液进行过夜固定,经包埋、切片、HE染色等处理后,采用光学显微镜进行观察。
1.3.2.3 物性指标测定
将实验样品剪成1.5 cm×1.5 cm×0.5 cm的肉块,采用质构仪参数设置:P36R探头;测前、测中、测后速率均为1.0 mm/s;测试时间间隔5 s;触发力5 g;数据采集速率400 pps;应变量均为75%,测定硬度和弹性2 个质构指标。
1.3.2.4 pH值测定
按照GB 5009.237—2016《食品pH值的测定》进行检测。
1.3.2.5 色泽及肌红蛋白氧化状态测定
色泽:采用色差仪进行黑白板校正测色仪后,测定样品(红度)。测试前将解冻完全的肉样在低温避光环境中暴露约15 min,放置在光源上方,平行测定3 次,取平均值。
肌红蛋白氧化状态:采用Krzywicki的方法。取解冻肉样5 g,加入20 mL 0.04 mol/L pH 6.8的磷酸钠缓冲液,室温(20 ℃)匀浆25 s。匀浆液在4 ℃冰箱中放置1 h,然后在15 ℃、950×离心30 min。上清液经滤纸过滤,用同样的缓冲液补足至25 mL,测定其在525、545、565、572 nm波长处的吸光度。脱氧肌红蛋白、氧合肌红蛋白和高铁肌红蛋白相对含量计算公式如下:

式中:为脱氧肌红蛋白相对含量/%;为氧合肌红蛋白相对含量/%;为高铁肌红蛋白相对含量/%;A为相应波长条件下的吸光度。
1.3.2.6 TBARS值测定
按照GB 5009.181—2016《食品中丙二醛的测定》第二法进行检测,结果用每千克肉中丙二醛的质量(mg/kg)表示。
1.3.2.7 TVB-N值测定
参照GB 5009.228—2016《食品中挥发性盐基氮的测定》,用半微量定氮法进行测定。
1.3.2.8 脂肪酸含量测定
按照GB/T 5009.168—2016《食品中脂肪酸的测定》进行检测。
粗脂肪的提取:称取样品各3 g,分别放入150 mL锥形瓶,加入60 mL氯仿-甲醇溶液,充分搅拌,用保鲜膜封口后置于摇床振荡5 h,振荡结束后放入40 ℃水浴30 min,再加入饱和NaCl溶液15 mL,待分层后取上层有机溶剂置于圆底烧瓶中旋转蒸干,每个样品重复3 次。
脂肪的皂化、甲酯化:向烧瓶中加入6 mL 0.5 mol/L氢氧化钠-甲醇溶液,上端连接冷凝管,67 ℃皂化30 min,当溶液中有油滴漂浮时,加入沸石,从冷凝管上端口加入15%三氟化硼-甲醇溶液5 mL,15 min后加入3 mL甲醇,再15 min后加入2 mL正己烷,取下烧瓶,加入20 mL饱和氯化钠溶液,剧烈振荡15 s,继续加饱和氯化钠至瓶颈,待分层后吸上层溶液放入1.5 mL离心管中,封存于-20 ℃。
脂肪酸分析:脂肪酸气相色谱-质谱联用检测条件:升温程序:初始温度120 ℃,保持1 min,8 ℃/min升到310 ℃,保持 5 min,汽化室温度为260 ℃;进样方式:不分流;载气:氢气;脂肪酸甲酯的相对含量采用面积归一法计算。
1.4 数据处理
采用Microsoft Excel 2013软件对数据进行处理及分析。采用SPSS 22.0中的ANOVA进行方差分析,Duncan多重检验进行差异显著性分析。通过OriginPro 2015软件进行聚类和基于双标图的相似性分析,聚类方法为类平均法,利用R语言3.6.1软件的pheatmap包进行热图绘制。
2 结果与分析
2.1 疏果多酚处理后冻融期间羊肉汁液流失率的变化

表1 冻融期间羊肉汁液流失率的变化Table 1 Changes in goat meat drip loss during freeze-thaw cycles%
由表1可知,4 组羊肉的汁液流失率都随着冻融次数的增加而显著增加(<0.05),其排序为疏果多酚组<TBHQ组<表儿茶素组<空白组。在冻融1、3、5 次时,疏果多酚组的汁液流失率均显著低于其余3 组处理(<0.05),在冻融1、3、5 次时,空白组的汁液流失率显著高于其余3 组(<0.05);在冻融结束时,疏果多酚组的汁液流失率较表儿茶素、TBHQ及空白组分别降低了17.57%、16.83%和40.10%。综上所述,多次冻融会加速羊肉汁液的流失,但疏果多酚的加入会减缓羊肉汁液的流失,从而可以改善羊肉中水溶性物质因汁液流失而造成的羊肉品质下降问题。
2.2 疏果多酚处理后冻融期间羊肉肌纤维显微结构的变化

图1 冻融期间羊肉肌纤维显微结构的变化(×400)Fig. 1 Microstructure changes of goat meat during freeze-thaw cycles (× 400)
由图1可知,4 组羊肉的显微结构均随着冻融次数增加结构变得不紧致、排列变得疏松、形状变得离散。在冻融期间,疏果多酚组、表儿茶素组、TBHQ组和空白组羊肉的肌纤维排列逐渐疏松、结构松弛;在冻融1 次时,4 组羊肉的肌纤维结构差异不大,其结构完整、轮廓清晰,排列致密有序、形状规则;而在冻融5 次时,空白组和TBHQ组羊肉的肌纤维结构完整性完全丧失,肌纤维排列混乱断裂、肌纤维间隙增大、结构疏松,致密结构几乎全部被破坏,而疏果多酚与表儿茶素组的肌纤维结构相对完整。综上可见,羊肉在疏果多酚处理下有利于维持其肌纤维结构的完整性。
2.3 疏果多酚处理后冻融期间羊肉物性的变化
由图2可知,4 组羊肉的硬度随冻融次数增加而降低,在冻融期间,4 组羊肉的硬度由大到小的顺序为疏果多酚组、表儿茶素组、TBHQ组和空白组,其中在冻融1 次时,疏果多酚组羊肉的硬度值显著高于表儿茶素组、TBHQ组和空白组29.57%、26.72%和36.90%(<0.05),冻融3 次后疏果多酚组和表儿茶素组羊肉的硬度相近,且疏果多酚组的硬度比空白组显著增加了27.01%(<0.05),在5 次冻融结束时疏果多酚组、表儿茶素组、TBHQ组和空白组的硬度分别较初始值降低了24.48%、23.59%、30.37%和37.28%;随着冻融次数的增加,4 组羊肉的弹性均呈现先降低后稳定的变化趋势,在冻融1 次时疏果多酚组的弹性显著高于空白组36.68%(<0.05),而在冻融3 次和5 次时,各组间无显著差异(>0.05),在冻融结束时疏果多酚组、表儿茶素组、TBHQ组和空白组的弹性分别较初始值降低了35.33%、36.59%、39.67%和40.22%。综上可见,酚类处理有利于羊肉物性的稳定保持,它可以减缓冻融羊肉硬度过度降低,保持肉口感。

图2 冻融期间羊肉物性的变化Fig. 2 Texture changes of goat meat during freeze-thaw cycles
2.4 疏果多酚处理后冻融期间羊肉pH值的变化
由图3可知,pH值变化总体范围较小为5.50~5.91;在冻融过程中疏果多酚组羊肉的pH值最高,冻融1 次时4 组间无显著差异(>0.05),在冻融3 次时,疏果多酚组的pH值比TBHQ组显著增加了4.62%(<0.05),冻融5 次结束后,疏果多酚组的pH值最高且比表儿茶素组显著升高了4.55%(<0.05)。综上可见,羊肉冻融次数越多其羊肉的pH值越低,且疏果多酚处理的冻融羊肉pH值相对最高。

图3 冻融期间羊肉pH值的变化Fig. 3 Changes in pH of goat meat during freeze-thaw cycles
2.5 疏果多酚处理后冻融期间羊肉色泽和肌红蛋白相对含量变化
值能反映肉表面色泽的红色程度,值越大则肉产品越红,其色泽越好。大量研究表明,反复冻融过程中水分的丧失导致色素相关物质流失,从而加剧肉色劣变。因此,选取冻融羊肉,并对其值进行分析,结果如图4所示,4 组羊肉的随着冻融次数增加而降低;在羊肉冻融期间,空白组羊肉值最低,表儿茶素组值最高(<0.05),且在冻融1 次后疏果多酚组和表儿茶素组羊肉的值逐渐变得接近,在冻融3 次时疏果多酚组值比TBHQ组显著增大了13.13%(<0.05),在冻融结束时,疏果多酚组羊肉的值比TBHQ组和空白组显著增大了35.74%和25.91%(<0.05),且分别是表儿茶素组、TBHQ组和空白组的1.02、1.25 倍和1.31 倍。

图4 冻融期间羊肉a*值的变化Fig. 4 Changes in a* value of goat meat during freeze-thaw cycles
脱氧肌红蛋白、氧合肌红蛋白和高铁肌红蛋白的含量决定羊肉的色泽,冻融期间羊肉肌红蛋白氧化状态变化如表2所示。随着冻融次数的增加,氧合肌红蛋白相对含量逐渐下降,其与图4中值的结果基本一致,而高铁肌红蛋白相对含量呈先下降后增加的趋势。疏果多酚组在冻融5 次时脱氧肌红蛋白含量显著高于其余3 组,这与值在冻融5 次时的变化规律一致;在冻融期间,表儿茶素组的氧合肌红蛋白相对含量最高(<0.05),在冻融1 次时,疏果多酚组的氧合肌红蛋白相对含量比TBHQ组和空白组显著增加了63.71%和20.66%(<0.05),在羊肉冻融3 次时,疏果多酚组的氧合肌红蛋白含量与表儿茶素组更接近,两者间无显著差异(>0.05),但比TBHQ组和空白组显著增加了18.54%和20.68%(<0.05);在羊肉冻融1 次后,疏果多酚组的高铁肌红蛋白相对含量最高(<0.05),但在冻融3 次时,疏果多酚组的高铁肌红蛋白相对含量最低为32.03%(<0.05),且较表儿茶素组、TBHQ组及空白组分别降低了4.45%、11.72%和18.39%,在冻融5 次结束后疏果多酚组的高铁肌红蛋白相对含量为34.01%,显著低于其余3 组(<0.05),较表儿茶素组、TBHQ组及空白组分别降低了15.21%、12.32%和9.67%。综上可见,经过多次冻融后,羊肉的红度和氧合肌红蛋白相对含量降低,高铁肌红蛋白相对含量增加,但酚类处理会延缓这种变化,从而确保肉品色泽稳定。

表2 冻融羊肉肌红蛋白氧化状态变化Table 2 Changes in chemical state of myoglobin on goat meat during freeze-thaw cycles%
2.6 疏果多酚处理后冻融期间羊肉TBARS值的变化
多次冻融也会导致脂肪氧化,本研究对羊肉的脂肪氧化情况(2-硫代巴比妥酸法)进行分析,冻融期间羊肉TBARS值结果如图5所示,4 组羊肉的TBARS值随着冻融次数增加均先减小后增加。在羊肉冻融期间,冻融1 次时4 组羊肉TBARS值无显著差异(>0.05),疏果多酚组的TBARS值最低为0.13 mg/kg;在冻融3 次时,疏果多酚组的TBARS值比TBHQ组和空白组显著降低了14.90%和27.76%(<0.05);在冻融5 次结束后,疏果多酚组TBARS值显著低于其余3 组(<0.05),且较表儿茶素组、TBHQ组及空白组分别降低了11.92%、16.12%和28.21%(<0.05)。总之,在不同的处理条件下,在羊肉中添加疏果多酚可以极大地抑制羊肉在冻融过程中的脂肪氧化。
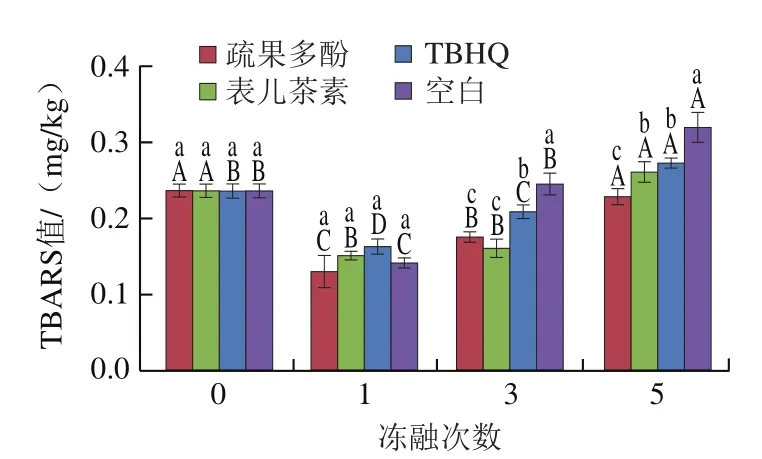
图5 冻融期间羊肉TBARS值的变化Fig. 5 Changes in TBARS value of goat meat during freeze-thaw cycles
2.7 疏果多酚处理后冻融期间羊肉TVB-N值的变化
如图6所示,4 组羊肉的TVB-N值随着冻融次数的增加而增加,其值排序为疏果多酚组<表儿茶素组<TBHQ组<空白组,在整个冻融期间,疏果多酚组的TVB-N值都显著低于其余3 组(<0.05);冻融结束时,疏果多酚组的TVB-N值分别为表儿茶素组、TBHQ组及空白组的67.78%、74.12%和65.26%,且在冻融结束时疏果多酚组的TVB-N值仅为3.91 mg/100 g,远小于GB 2707—2016《鲜(冻)畜、禽产品》中规定的15 mg/100 g。综上,反复冻融会加速羊肉腐败变质,但疏果多酚的加入会延缓这种变化,从而保证羊肉在反复冻融过程中的新鲜。

图6 冻融期间羊肉TVB-N值的变化Fig. 6 Changes in TVB-N value of goat meat during freeze-thaw cycles
2.8 疏果多酚处理后冻融期间羊肉脂肪酸相对含量变化
如图7所示,4 种处理条件下羊肉共检出16 种脂肪酸,包括饱和脂肪酸(saturated fatty acid,SFA)8 种,单不饱和脂肪酸(monounsaturated fatty acid,MUFA)4 种,多不饱和脂肪酸(polyunsaturated fatty acid,PUFA)4 种。随着冻融次数的增加,SFA相对含量呈下降趋势,MUFA相对含量呈上升趋势,PUFA相对含量保持稳定;随着冻融次数的增加,疏果多酚组的MUFA相对含量先上升后降低且在冻融3 次达到最高值为56.59%;随着冻融次数的增加,疏果多酚组的PUFA相对含量呈现先上升后降低的趋势且在冻融1 次达到最高值,其相对含量分别为TBHQ组、表儿茶素组及空白组的1.42、1.49 倍和1.50 倍。羊肉中C、C、C 相对含量较高,且这3 种脂肪酸随着冻融次数的增加含量相对稳定,疏果多酚对C、C和C效果明显;经过冻融1次后,疏果多酚处理组的C、C和C相对含量高于其余3 组;冻融3 次后,TBHQ组的C相对含量高于其他组,猕猴桃多酚组的C和C相对含量是4 个处理组中最高的;5 次冻融结束后,疏果多酚组C含量高于其余3 组,而除空白组外,表儿茶素组的C和C相对含量为其余3 组中最高的。
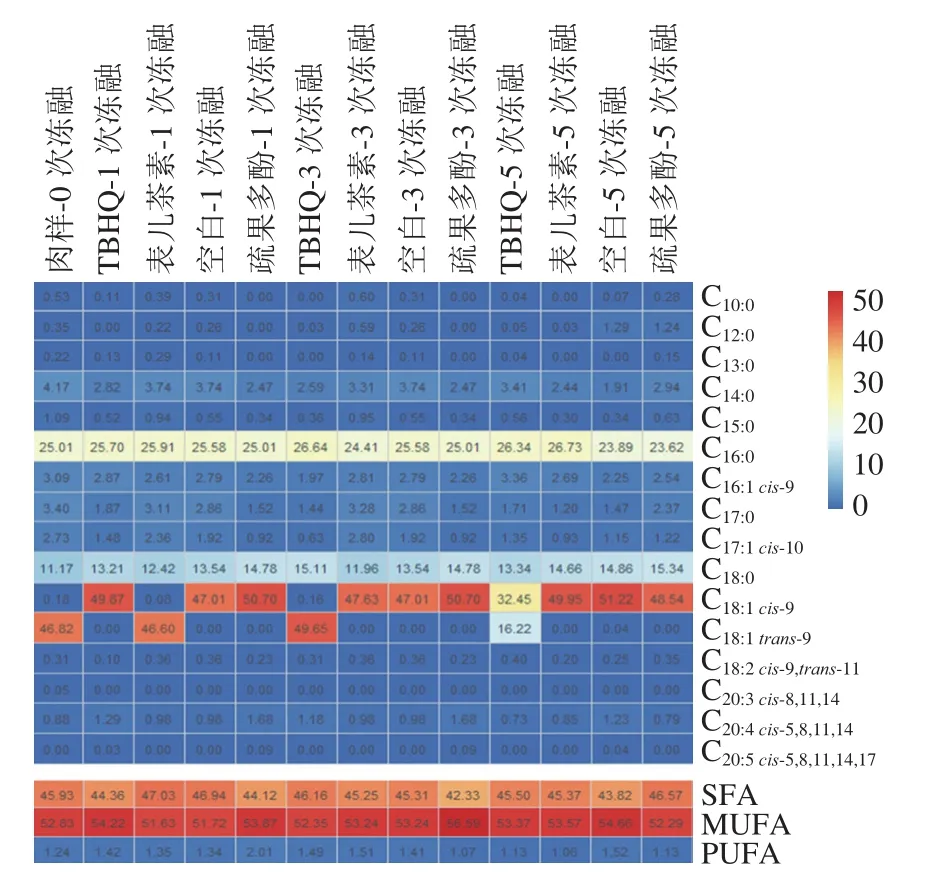
图7 羊肉脂肪酸相对含量的热图Fig. 7 Heat map of relative contents of fatty acids in goat meat
4 组样品在反复冻融条件下用16 个脂肪酸相对含量作为聚类变量。如图8所示,样品可分为3 类:未冻融肉样和冻融1 次的表儿茶素组为第1类,脂肪酸营养品质最高,说明冻融1 次后,表儿茶素组的脂肪酸组成及相对含量与未冻融肉样最接近,相比其余3 组处理表儿茶素组对羊肉的效果更好;冻融1 次的TBHQ组和疏果多酚组、冻融5 次的表儿茶素组、冻融3 次的疏果多酚组和TBHQ组为第2类,品质一般,且冻融1 次和冻融3 次的疏果多酚组的肉样相较其他肉样更为接近,说明冻融次数的增加对疏果多酚处理组影响不大,冻融1 次的TBHQ组和冻融5 次的表儿茶素组这两组的肉样较为接近,说明表儿茶素组和TBHQ组相比,表儿茶素处理对羊肉的效果较好;冻融1 次和3 次的空白组、冻融3 次的表儿茶素组、冻融5 次的TBHQ组、空白组和疏果多酚组为第3类,品质开始变差,说明冻融3 次后疏果多酚处理对羊肉的效果较好,冻融5 次后表儿茶素处理对羊肉的效果较好。该聚类结果与之前脂肪酸相对含量分析结果相似。

图8 多次冻融羊肉相对脂肪酸含量的聚类分析Fig. 8 Cluster analysis of relative contents of fatty acids in goat meat subjected to multiple freeze-thaw cycles
利用双标图分析各脂肪酸在不同处理羊肉评价上的相似性,从中心到各个脂肪酸作一条线段,两脂肪酸线段之间夹角的余弦值是它们的相关系数,夹角小于90度表示正相关,线段的长度是脂肪酸对不同处理羊肉的区分能力,线段越长,区分能力越强。相关性结果如图9所示,脂肪酸C、C、C夹角较小,存在正相关;C、C、C 、C、夹角较小,存在正相关;C、C夹角较小,存在正相关;C、C、C 、C 比其他脂肪酸有较强的区分能力。双标图中点之间的距离,反映它们对应的样本之间的差异大小,两点相距较远,对应样本差异大;两点相距较近,对应样本差异小,存在相似性。在反复冻融过程,冻融1 次的表儿茶素组与未冻融肉样两点之间的距离接近,说明这两组样本差异小;冻融1 次和冻融3 次的疏果多酚组两点相距近,则这两组间差异小,更进一步说明在冻融3 次后疏果多酚组的羊肉脂肪酸品质高于其余处理组。

图9 基于双标图的羊肉脂肪酸相似性分析Fig. 9 PCA biplot for similarity analysis of fatty acids in goat meat
综上可见,反复冻融对羊肉脂肪酸有较大影响,在冻融3 次时疏果多酚处理羊肉的效果最优,在冻融5 次时表儿茶素处理的效果较优于疏果多酚组,但疏果多酚处理效果仅次于表儿茶素组。
3 讨 论
随着冻融次数的增加,羊肉的汁液流失不断增大。彭新颜等在研究反复冷冻-解冻对猪肉品质特性和微观结构的影响时,发现随着冷冻-解冻次数的增加,猪肉的解冻损失不断增加;这是因为羊肉被冷冻后,肌肉细胞内水分变成冰晶,体积增大对肌肉细胞膜和细胞器造成机械损伤,肌肉的保水能力下降,其汁液流失随着冻融次数的增加而增大,并且在冷冻贮藏期间,由冰晶引起的肌肉微观结构的变化以及肌纤维蛋白的变性,也会导致解冻时汁液损失增加。此外,由于冻融过程中冰晶的破坏作用,羊肉肌原纤维断裂,导致肌肉硬度减小、弹性下降;在冻融过程中,羊肉的硬度和弹性均有明显变化,说明冻融会影响羊肉物性,这与戚军研究不同冻融次数羊肉加工特性变化得到的结果一致。用疏果多酚处理羊肉有利于维持其肌纤维结构的完整性,能够提高肌肉的保水能力,减少汁液流失,这也与酚类对肉中肌纤维蛋白聚集模式的影响有关;而效果为疏果多酚尤于表儿茶素,推测主要是因为疏果多酚中含有丰富的多种酚类物质,更有利于保持羊肉肌纤维结构稳定性。此外,肉类脂肪在贮藏过程中会经常发生氧化水解现象,而冻融会加剧这一过程,并且脂肪氧化产生的自由基会诱导蛋白质与脂质之间发生自由基反应,从而形成蛋白质-脂质和蛋白质-蛋白质交联网络,而多酚类物质能够通过抑制氧化稳定蛋白质交联网络,从而保持羊肉的物性稳定。因此,疏果多酚之所以能够维持羊肉物性的稳定,可能与多酚延缓脂肪氧化有关。另外肉产品的颜色为消费者对肉的第一印象,新鲜的肉红色能够增强消费者的购买欲望,但反复冻融会加剧肉色劣变,影响肉产品销售,陈骋等对牦牛肉经过冻融处理后测定其表面色泽,研究表明多次冻融使肉的红色度降低,失去鲜肉应有的颜色。并且有研究表明,肉色泽受其中肌红蛋白形态和含量的影响,其中肌红蛋白呈紫红色,氧合肌红蛋白呈鲜红色,高铁肌红蛋白呈褐色,在冻融期间肉中氧合肌红蛋白会被氧化生成褐色的高铁肌红蛋白,使肉色变暗、肉品质下降。在冻融过程中,肉的值随着冻融次数增加而降低,并且氧合肌红蛋白相对含量也会下降,这可能是由于随着冻融次数增加,微生物数量逐渐增多,从而促进肉中脂肪的氧化,导致肉表面褪色,并且氧合肌红蛋白不稳定,很容易被氧化成褐色的高铁肌红蛋白,导致氧合肌红蛋白含量降低;相比之下,酚类处理会延缓肉色劣变,从而确保肉色泽稳定,也从一定程度上反映了其抑制脂肪氧化的功能。另一方面,反复冻融过程中肉类水分的流失会引起色素相关物质流失,进而导致肉值的降低;同时冻融期间羊肉中矿物质及小分子蛋白质混合物的损失,会导致肉中离子平衡被破坏,进而造成pH值略有下降,蛋白质分子的静电荷效应改变肌肉保水性,这与本研究冻融期间羊肉pH值下降的结果一致。
本研究显示羊肉在1 次冻融时TBARS值有减小趋势,与Qi Jun等将绵羊背最长肌反复冻融过程中TBARS值的变化结果不同,这可能与羊肉中的脂肪氧化程度有关,前期羊肉脂肪氧化产物丙二醛与肉中活性氨基作用生成1-氨基-3-氨基丙烯造成TBARS值减小,而后期由于不饱和脂肪酸在氧的作用下发生氧化生成的醛和酮造成TBARS值增加,说明脂肪氧化加剧。并且本研究发现加入疏果多酚能够降低TBARS值增长的速率,即能减缓羊肉在冻融过程中脂肪氧化的过程,由于疏果多酚的加入减缓了不饱和脂肪酸的氧化,PUFA的损失速率也下降,这与疏果多酚组PUFA相对含量增加一致。羊肉中脂肪酸的组成和含量不仅可以衡量羊肉的营养品质,而且还会影响肉质的硬度、嫩度以及风味。
羊肉在冻融期间一部分脂肪会随着汁液流失而损失,这就造成脂肪酸总含量会减少。本研究发现随着冻融次数的增加,SFA相对含量呈下降趋势,MUFA相对含量呈上升趋势,PUFA相对含量保持稳定;冻融过程中游离脂肪酸含量变化比较复杂,是一个动态的过程,一方面磷脂和甘油脂水解成游离脂肪酸,使其含量增加,另一方面游离脂肪酸氧化,含量减少。SFA相对含量显著下降,这是由于脂类的氧化作用;MUFA相对含量呈上升趋势是因为水分的流失,肌肉细胞液浓度增大,脂肪酶和磷脂酶活力急剧增加,使得MUFA相对含量增加;疏果多酚组MUFA的相对含量会随着冻融次数的增加出现先升高后下降的趋势,在冻融5 次结束后疏果多酚组MUFA相对含量下降可能是由于疏果多酚阻止了磷脂和甘油酯水解成游离脂肪酸。
羊肉的TVB-N值与新鲜度相关,在冻融期间,TVB-N值呈缓慢上升趋势,可能是反复地冻融破坏了肌纤维结构,而由于肌纤维结构的降解和水分的流失,羊肉自身的酶被释放出来,在酶和微生物共同作用下,蛋白质及其他含氮物质被分解所致,而加入疏果多酚可以减缓冻融过程中TVB-N的增加,这可能是因为疏果多酚影响了肌纤维蛋白的聚集模式,有利于维持肌纤维的稳定性,使得肌肉细胞的内源性酶减少释放,同时,也可能与疏果多酚具有的抗菌作用有关。
4 结 论
羊肉在反复冻融过程中品质会发生明显劣变。疏果多酚较表儿茶素、TBHQ能有效改善羊肉在反复冻融过程中的汁液流失问题;并且有利于维持冻融羊肉肌纤维结构的完整性,保持其物性的稳定,保持羊肉口感;冻融过程会降低羊肉红度和氧合肌红蛋白相对含量,增加其高铁肌红蛋白相对含量,但疏果多酚处理会延缓这种变化,从而保证肉色泽稳定;同时在冻融过程中,添加疏果多酚可以极大地抑制羊肉脂肪氧化,能较好地保持脂肪酸相对含量的稳定,延缓羊肉腐败变质。因此,疏果多酚在反复冻融过程中能有效改善羊肉品质和延缓氧化,延长冻融羊肉的货架期。研究结果将为猕猴桃疏果多酚在肉品保鲜中的应用提供理论参考和技术支撑,同时有利于猕猴桃疏果资源的综合利用。
猜你喜欢
健康博览(2021年11期)2021-12-10
作文·小学中高年级(2021年2期)2021-07-01
健康博览(2020年11期)2020-11-23
科教新报(2020年15期)2020-10-21
家庭医药(2020年6期)2020-06-30
江苏农业科学(2020年9期)2020-06-21
饮食与健康·下旬刊(2019年10期)2019-03-09
农村百事通(2019年1期)2019-01-30
中国绿色画报(2017年8期)2017-09-01
农家参谋(2017年1期)2017-08-13
